
Prenatal Exposure to an EDC Mixture, NeuroMix: Effects on Brain, Behavior, and Stress Responsiveness in Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry
2.2. NeuroMix Design and Preparation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Dose | Vendor | Catalog # (Lot #) | NOAEL |
|---|---|---|---|---|
| Aroclor 1221 (A1221) | 100 µg/kg | Accustandard | C-221N-50MG (072-202-01) | 1 mg/kg/d [48] |
| Bisphenol A (BPA) | 2.5 µg/kg | Accustandard | BPA-A-N (11625) | 5 mg/kg/d [49] |
| Bisphenol S (BPS) | 2.5 µg/kg | Accustandard | BPA-S-N (20696) | 20 mg/kg/d [50] |
| Di(2-EthylHexyl)- | 20 µg/kg | Accustandard | ALR-097N (03312KZ-1-01) | 4.8 mg/kg/d [51] |
| phthalate (DEHP) | ||||
| Di-n-butyl phthalate (DBP) | 20 µg/kg | Accustandard | ALR-104N (24227) | 50 mg/kg/d [52] |
| Perfluorooctane | 20 µg/kg | Accustandard | PFOS-002N (24187) | 0.4 mg/kg/d [53] |
| sulfonate (PFOS) | ||||
| Polybrominated diphenyl | 100 µg/kg | Sigma | BCBR7972 (91834) | 0.7 mg/kg/d [54] |
| ether 47 (PBDE-47) | ||||
| PCB-153 | 100 µg/kg | Chem Service | BZ-153-10MG (7582000) | 16 mg/kg/d [55] |
| Vinclozolin | 100 µg/kg | Chem Service | N-13745-250MG (7416000) | 6–12 mg/kg/d [56] |
2.3. Chronic Restraint Stress
2.4. Behavioral Testing
2.4.1. Light:Dark Box
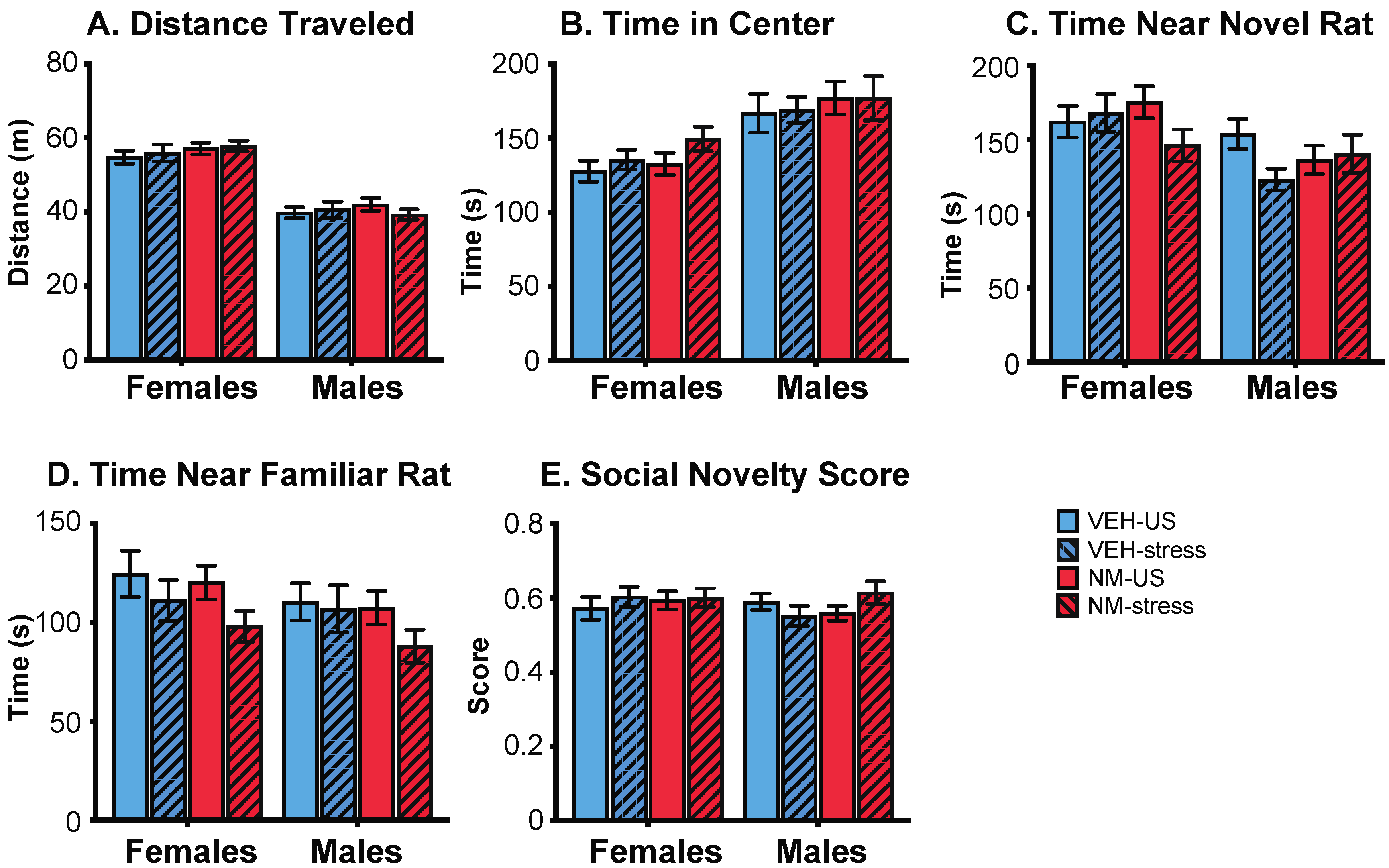
2.4.2. Sociability and Social Novelty
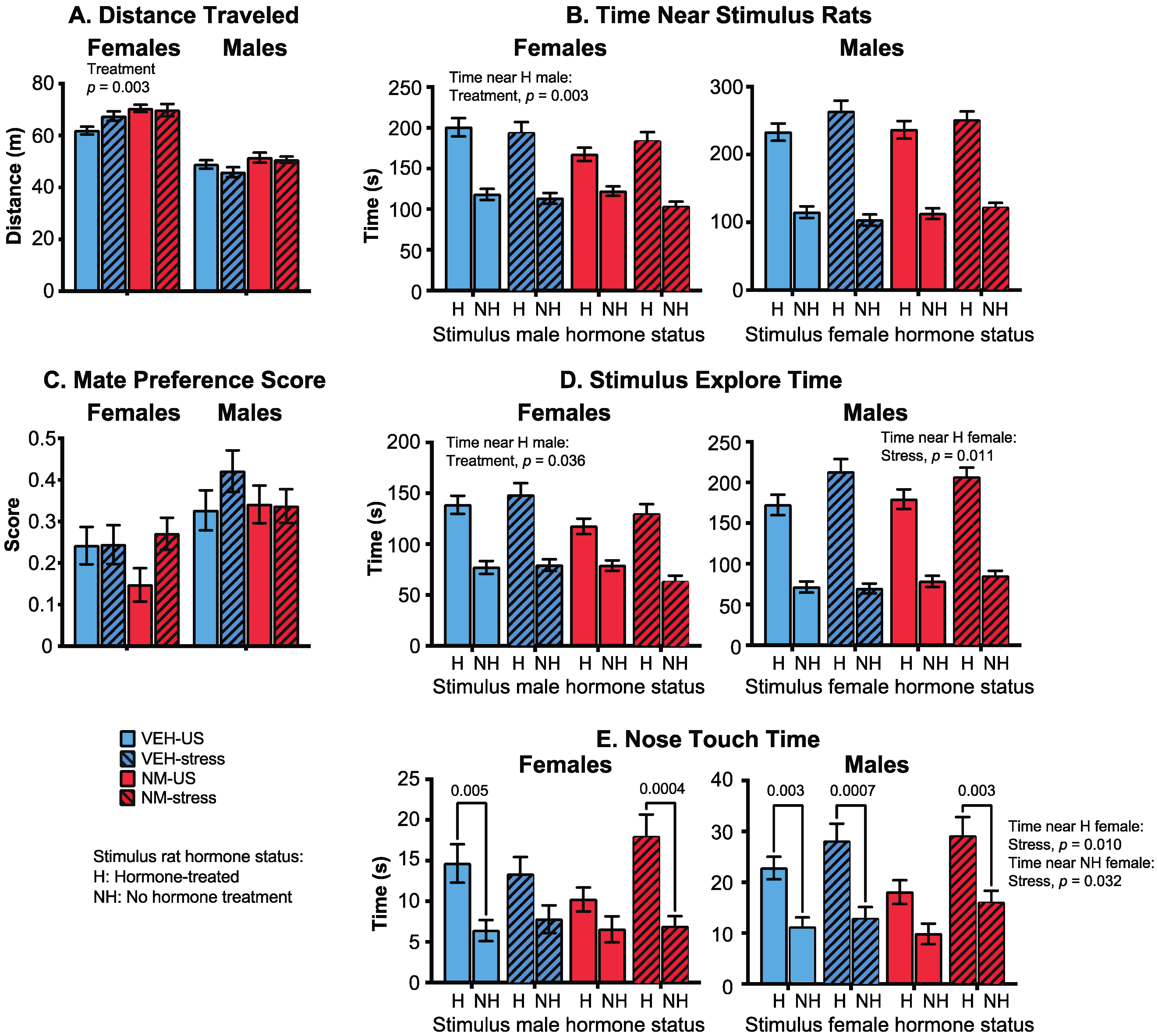
2.4.3. Mate Preference
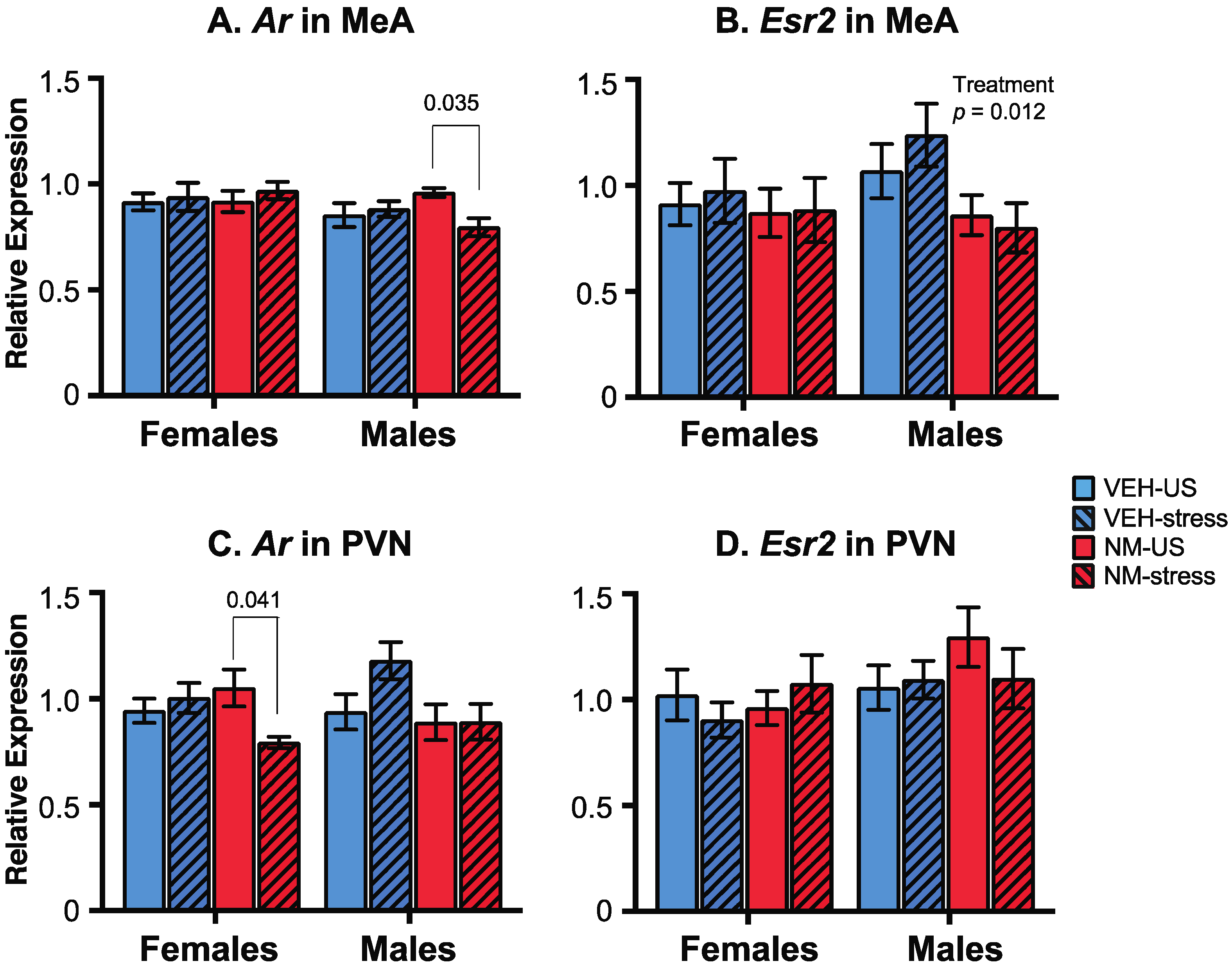
2.5. Brain Preparation, RNA Isolation, and qPCR
2.6. Corticosterone RIA
2.7. Statistical Analysis
3. Results
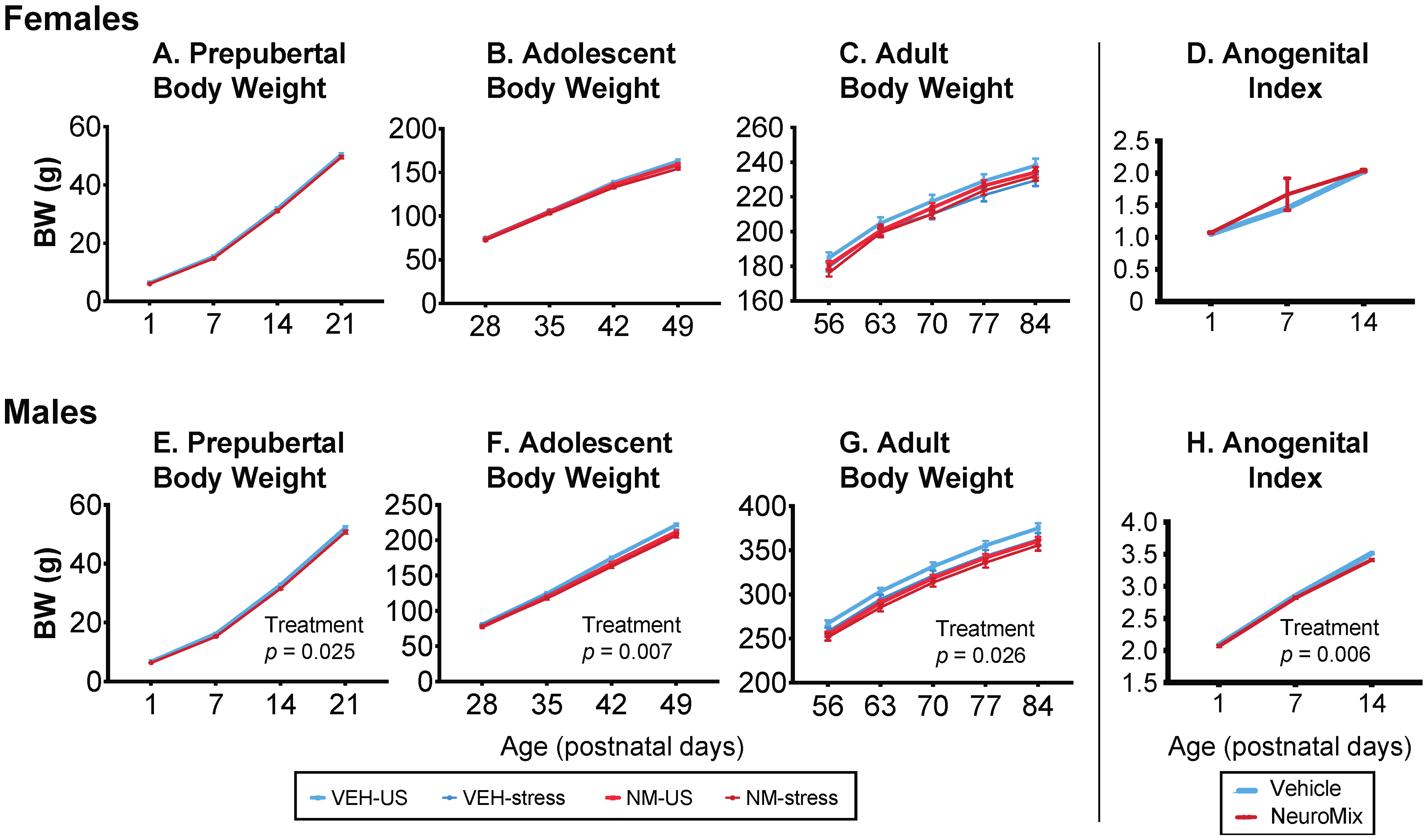
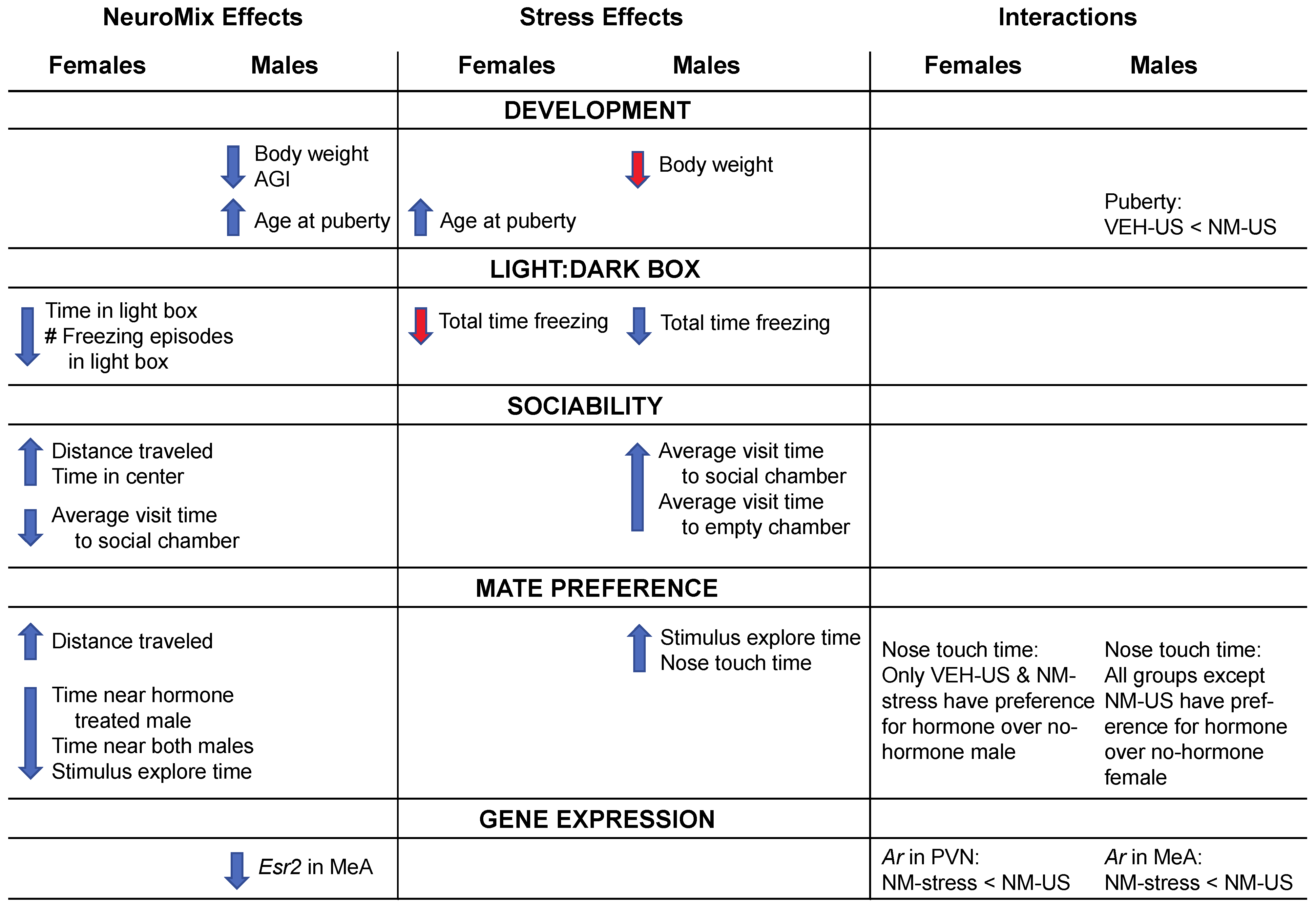
3.1. Development
3.1.1. Body Weight
3.1.2. Anogenital Index (AGI)
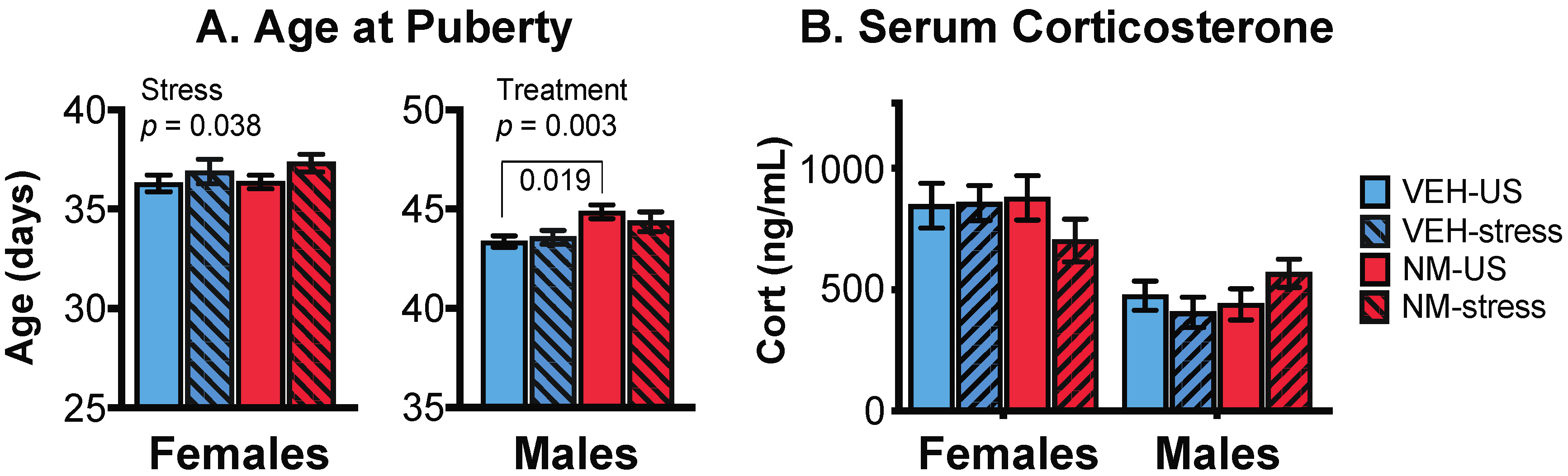
3.1.3. Puberty
3.2. Serum Corticosterone
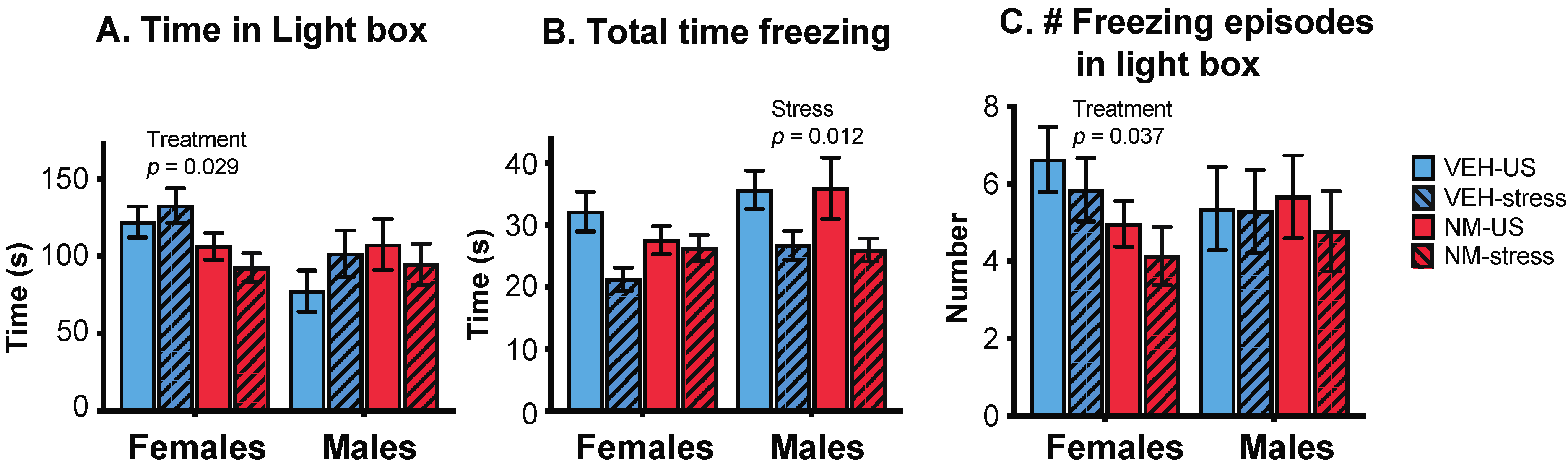
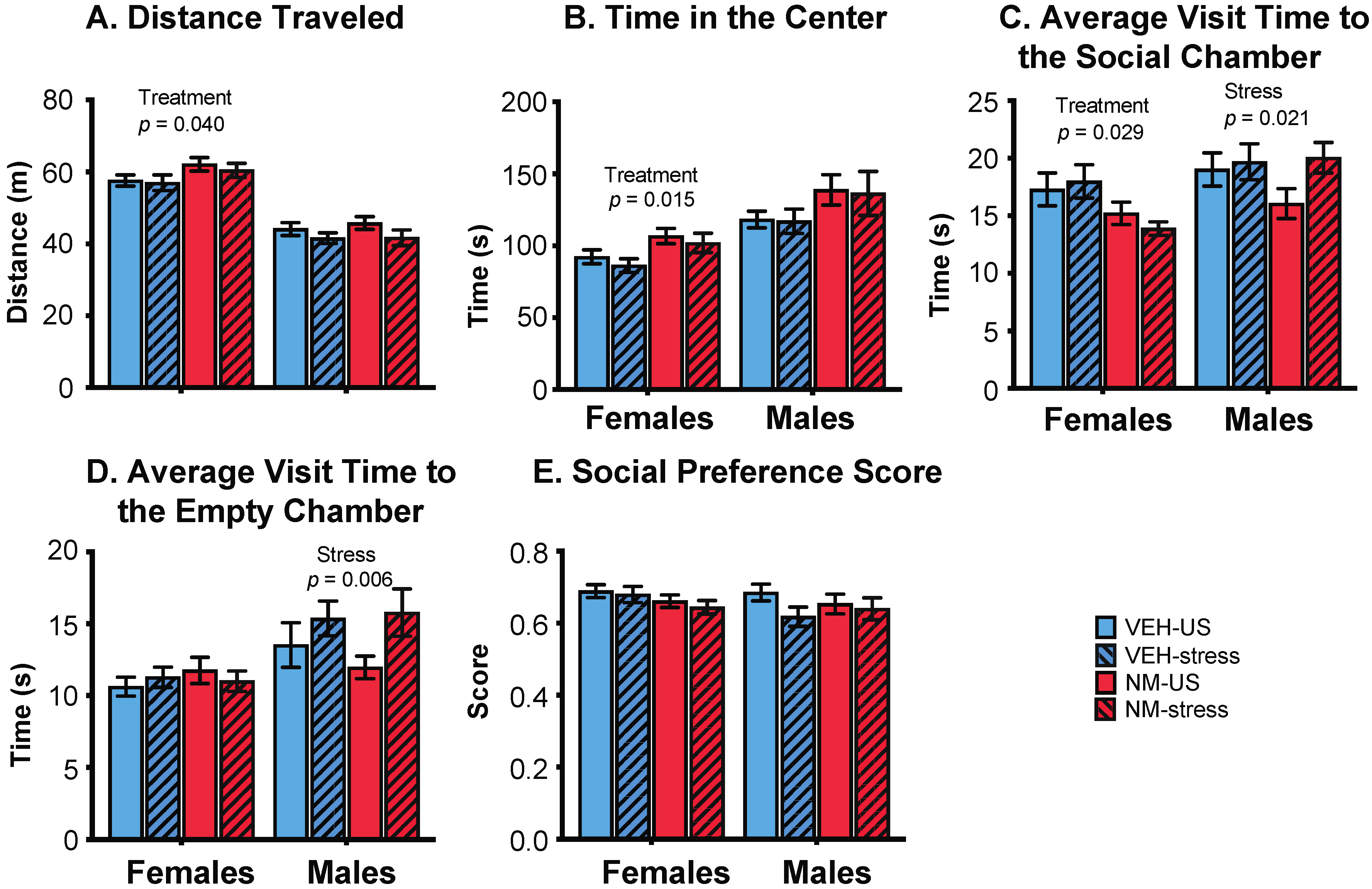
3.3. Behaviors
3.3.1. Light:Dark Box (LD)
3.3.2. Sociability
3.3.3. Social Novelty
3.3.4. Mate Preference
3.4. Gene Expression
4. Discussion
4.1. EDC and Stress Effects on Postnatal Development
4.2. EDC and Stress Effects on Behaviors
4.3. EDC and Stress Effects on Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gore, A.C.; Krishnan, K.; Reilly, M.P. Endocrine-disrupting chemicals: Effects on neuroendocrine systems and the neurobiology of social behavior. Horm. Behav. 2019, 111, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, A.W.; Beach, E.C.; Stetzik, L.A.; Perry, A.; D’Addezio, A.S.; Cushing, B.S.; Patisaul, H.B. A novel model for neuroendocrine toxicology: Neurobehavioral effects of BPA exposure in a prosocial species, the prairie vole (Microtus ochrogaster). Endocrinology 2014, 155, 3867–3881. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Bisphenol A and phthalate endocrine disruption of parental and social behaviors. Front. Neurosci. 2015, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reilly, M.P.; Weeks, C.D.; Crews, D.; Gore, A.C. Application of a Novel Social Choice Paradigm to Assess Effects of Prenatal Endocrine-Disrupting Chemical Exposure in Rats (Rattus norvegicus). J. Comp. Psychol. 2018, 132, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Reilly, M.P.; Weeks, C.D.; Topper, V.Y.; Thompson, L.M.; Crews, D.; Gore, A.C. The effects of prenatal PCBs on adult social behavior in rats. Horm. Behav. 2015, 73, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Topper, V.Y.; Reilly, M.P.; Wagner, L.M.; Thompson, L.M.; Gillette, R.; Crews, D.; Gore, A.C. Social and neuromolecular phenotypes are programmed by prenatal exposures to endocrine-disrupting chemicals. Mol. Cell Endocrinol. 2019, 479, 133–146. [Google Scholar] [CrossRef]
- Jones, B.A.; Watson, N.V. Perinatal BPA exposure demasculinizes males in measures of affect but has no effect on water maze learning in adulthood. Horm. Behav. 2012, 61, 605–610. [Google Scholar] [CrossRef]
- Tian, Y.H.; Baek, J.H.; Lee, S.Y.; Jang, C.G. Prenatal and postnatal exposure to bisphenol a induces anxiolytic behaviors and cognitive deficits in mice. Synapse 2010, 64, 432–439. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S.; Belzunces, L.P.; Canivenc, M.C.; Schroeder, H.; Chevrier, C.; Pasquier, E. Impairment of learning and memory performances induced by BPA: Evidences from the literature of a MoA mediated through an ED. Mol. Cell Endocrinol. 2018, 475, 54–73. [Google Scholar] [CrossRef]
- Kobrosly, R.W.; Evans, S.; Miodovnik, A.; Barrett, E.S.; Thurston, S.W.; Calafat, A.M.; Swan, S.H. Prenatal phthalate exposures and neurobehavioral development scores in boys and girls at 6–10 years of age. Environ. Health Perspect. 2014, 122, 521–528. [Google Scholar] [CrossRef] [Green Version]
- England-Mason, G.; Martin, J.W.; MacDonald, A.; Kinniburgh, D.; Giesbrecht, G.F.; Letourneau, N.; Dewey, D. Similar names, different results: Consistency of the associations between prenatal exposure to phthalates and parent-ratings of behavior problems in preschool children. Environ. Int. 2020, 142, 105892. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.J.; Joo, H.; Kim, D.; Lim, M.H.; Kim, E.; Ha, M.; Kwon, H.J.; Paik, K.C.; Kim, K.M. Associations between Exposure to Bisphenol A and Behavioral and Cognitive Function in Children with Attention-deficit/Hyperactivity Disorder: A Case-control Study. Clin. Psychopharmacol. Neurosci. 2020, 18, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, F.; Nolte, E.L.R.; Wang, Y.; Margolis, A.E.; Calafat, A.M.; Wang, S.; Garcia, W.; Hoepner, L.A.; Peterson, B.S.; Rauh, V.; et al. Bisphenol A exposure and symptoms of anxiety and depression among inner city children at 10–12 years of age. Environ. Res. 2016, 151, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeir, G.; Covaci, A.; Van Larebeke, N.; Schoeters, G.; Nelen, V.; Koppen, G.; Viaene, M. Neurobehavioural and cognitive effects of prenatal exposure to organochlorine compounds in three year old children. BMC Pediatr. 2021, 21, 99. [Google Scholar] [CrossRef]
- Jasarevic, E.; Williams, S.A.; Vandas, G.M.; Ellersieck, M.R.; Liao, C.; Kannan, K.; Roberts, R.M.; Geary, D.C.; Rosenfeld, C.S. Sex and dose-dependent effects of developmental exposure to bisphenol A on anxiety and spatial learning in deer mice (Peromyscus maniculatus bairdii) offspring. Horm. Behav. 2013, 63, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, S.; Matsuzawa, D.; Ishii, D.; Tomizawa, H.; Sutoh, C.; Nakazawa, K.; Amano, K.; Sajiki, J.; Shimizu, E. Effects of perinatal exposure to low dose of bisphenol A on anxiety like behavior and dopamine metabolites in brain. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 273–279. [Google Scholar] [CrossRef]
- Patisaul, H.B.; Bateman, H.L. Neonatal exposure to endocrine active compounds or an ERbeta agonist increases adult anxiety and aggression in gonadally intact male rats. Horm. Behav. 2008, 53, 580–588. [Google Scholar] [CrossRef]
- Xu, X.; Hong, X.; Xie, L.; Li, T.; Yang, Y.; Zhang, Q.; Zhang, G.; Liu, X. Gestational and lactational exposure to bisphenol-A affects anxiety- and depression-like behaviors in mice. Horm. Behav. 2012, 62, 480–490. [Google Scholar] [CrossRef]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and transgenerational effects of low doses of perinatal di-(2-ethylhexyl) phthalate (DEHP) on social behaviors in mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef]
- Carbone, S.; Ponzo, O.J.; Gobetto, N.; Samaniego, Y.A.; Reynoso, R.; Scacchi, P.; Moguilevsky, J.A.; Cutrera, R. Antiandrogenic effect of perinatal exposure to the endocrine disruptor di-(2-ethylhexyl) phthalate increases anxiety-like behavior in male rats during sexual maturation. Horm. Behav. 2013, 63, 692–699. [Google Scholar] [CrossRef]
- Bell, M.R.; Thompson, L.M.; Rodriguez, K.; Gore, A.C. Two-hit exposure to polychlorinated biphenyls at gestational and juvenile life stages: 1. Sexually dimorphic effects on social and anxiety-like behaviors. Horm. Behav. 2016, 78, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Gillette, R.; Reilly, M.P.; Topper, V.Y.; Thompson, L.M.; Crews, D.; Gore, A.C. Anxiety-like behaviors in adulthood are altered in male but not female rats exposed to low dosages of polychlorinated biphenyls in utero. Horm. Behav. 2017, 87, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobolewski, M.; Conrad, K.; Allen, J.L.; Weston, H.; Martin, K.; Lawrence, B.P.; Cory-Slechta, D.A. Sex-specific enhanced behavioral toxicity induced by maternal exposure to a mixture of low dose endocrine-disrupting chemicals. Neurotoxicology 2014, 45, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, E.; Rajapakse, N.; Kortenkamp, A. Something from “nothing”—eight weak estrogenic chemicals combined at concentrations below NOECs produce significant mixture effects. Environ. Sci. Technol. 2002, 36, 1751–1756. [Google Scholar] [CrossRef]
- Conley, J.M.; Lambright, C.S.; Evans, N.; Cardon, M.; Medlock-Kakaley, E.; Wilson, V.S.; Gray, L.E., Jr. A mixture of 15 phthalates and pesticides below individual chemical no observed adverse effect levels (NOAELs) produces reproductive tract malformations in the male rat. Environ. Int. 2021, 156, 106615. [Google Scholar] [CrossRef] [PubMed]
- Gaudriault, P.; Mazaud-Guittot, S.; Lavoué, V.; Coiffec, I.; Lesné, L.; Dejucq-Rainsford, N.; Scholze, M.; Kortenkamp, A.; Jégou, B. Endocrine Disruption in Human Fetal Testis Explants by Individual and Combined Exposures to Selected Pharmaceuticals, Pesticides, and Environmental Pollutants. Environ. Health Perspect. 2017, 125, 087004. [Google Scholar] [CrossRef] [PubMed]
- Fini, J.B.; Mughal, B.B.; Le Mevel, S.; Leemans, M.; Lettmann, M.; Spirhanzlova, P.; Affaticati, P.; Jenett, A.; Demeneix, B.A. Human amniotic fluid contaminants alter thyroid hormone signalling and early brain development in Xenopus embryos. Sci. Rep. 2017, 7, 43786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overgaard, A.; Holst, K.; Mandrup, K.R.; Boberg, J.; Christiansen, S.; Jacobsen, P.R.; Hass, U.; Mikkelsen, J.D. The effect of perinatal exposure to ethinyl oestradiol or a mixture of endocrine disrupting pesticides on kisspeptin neurons in the rat hypothalamus. Neurotoxicology 2013, 37, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Crofton, K.M.; Craft, E.S.; Hedge, J.M.; Gennings, C.; Simmons, J.E.; Carchman, R.A.; Carter, W.H., Jr.; DeVito, M.J. Thyroid-hormone-disrupting chemicals: Evidence for dose-dependent additivity or synergism. Environ. Health Perspect. 2005, 113, 1549–1554. [Google Scholar] [CrossRef] [Green Version]
- Martin, O.V.; Evans, R.M.; Faust, M.; Kortenkamp, A. A Human Mixture Risk Assessment for Neurodevelopmental Toxicity Associated with Polybrominated Diphenyl Ethers Used as Flame Retardants. Environ. Health Perspect. 2017, 125, 087016. [Google Scholar] [CrossRef] [Green Version]
- Bornehag, C.G.; Kitraki, E.; Stamatakis, A.; Panagiotidou, E.; Rudén, C.; Shu, H.; Lindh, C.; Ruegg, J.; Gennings, C. A Novel Approach to Chemical Mixture Risk Assessment-Linking Data from Population-Based Epidemiology and Experimental Animal Tests. Risk Anal. 2019, 39, 2259–2271. [Google Scholar] [CrossRef] [PubMed]
- Schildroth, S.; Wise, L.A.; Wesselink, A.K.; De La Cruz, P.; Bethea, T.N.; Weuve, J.; Fruh, V.; Botelho, J.C.; Sjodin, A.; Calafat, A.M.; et al. Correlates of Persistent Endocrine-Disrupting Chemical Mixtures among Reproductive-Aged Black Women. Environ. Sci. Technol. 2021, 55, 14000–14014. [Google Scholar] [CrossRef] [PubMed]
- Champagne, F.A.; Meaney, M.J. Stress During Gestation Alters Postpartum Maternal Care and the Development of the Offspring in a Rodent Model. Biol. Psychiatry 2006, 59, 1227–1235. [Google Scholar] [CrossRef]
- Romeo, R.D. Perspectives on stress resilience and adolescent neurobehavioral function. Neurobiol. Stress 2015, 1, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, M.R.; Hart, B.G.; Gore, A.C. Two-hit exposure to polychlorinated biphenyls at gestational and juvenile life stages: 2. Sex-specific neuromolecular effects in the brain. Mol. Cell Endocrinol. 2016, 420, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, R.A.; Kiss Von Soly, S.; Ratnayake, U.; Klug, M.; Binder, M.D.; Hannan, A.J.; van den Buuse, M. Long-term effects of combined neonatal and adolescent stress on brain-derived neurotrophic factor and dopamine receptor expression in the rat forebrain. Biochim. Biophys. Acta 2014, 1842, 2126–2135. [Google Scholar] [CrossRef] [Green Version]
- Hill, R.A.; Klug, M.; Kiss Von Soly, S.; Binder, M.D.; Hannan, A.J.; van den Buuse, M. Sex-specific disruptions in spatial memory and anhedonia in a “two hit“ rat model correspond with alterations in hippocampal brain-derived neurotrophic factor expression and signaling. Hippocampus 2014, 24, 1197–1211. [Google Scholar] [CrossRef]
- Panagiotidou, E.; Zerva, S.; Mitsiou, D.J.; Alexis, M.N.; Kitraki, E. Perinatal exposure to low-dose bisphenol A affects the neuroendocrine stress response in rats. J. Endocrinol. 2014, 220, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Gillette, R.; Dias, M.; Reilly, M.P.; Thompson, L.M.; Castillo, N.J.; Vasquez, E.L.; Crews, D.; Gore, A.C. Two Hits of EDCs Three Generations Apart: Effects on Social Behaviors in Rats, and Analysis by Machine Learning. Toxics 2022, 10, 30. [Google Scholar] [CrossRef]
- Krishnan, K.; Mittal, N.; Thompson Lindsay, M.; Rodriguez-Santiago, M.; Duvauchelle Christine, L.; Crews, D.; Gore Andrea, C. Effects of the endocrine-disrupting chemicals, vinclozolin and polychlorinated biphenyls, on physiological and sociosexual phenotypes in F2 generation Sprague-Dawley rats. Environ. Health Perspect. 2018, 126, 097005. [Google Scholar] [CrossRef]
- Dickerson, S.M.; Cunningham, S.L.; Patisaul, H.B.; Woller, M.J.; Gore, A.C. Endocrine disruption of brain sexual differentiation by developmental PCB exposure. Endocrinology 2011, 152, 581–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, A.P.; Gorski, R.A. Gonadal steroid induction of structural sex differences in the central nervous system. Ann. Rev. Neurosci. 1984, 7, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, J.T.; Freedman, A.N.; Kepper, S.L.; Keshava, A.M.; Bangma, J.T.; Fry, R.C. Per- and Polyfluoroalkyl Substances Differentially Inhibit Placental Trophoblast Migration and Invasion In Vitro. Toxicol. Sci. 2020, 175, 210–219. [Google Scholar] [CrossRef]
- Björvang, R.D.; Vinnars, M.T.; Papadogiannakis, N.; Gidlöf, S.; Mamsen, L.S.; Mucs, D.; Kiviranta, H.; Rantakokko, P.; Ruokojärvi, P.; Lindh, C.H.; et al. Mixtures of persistent organic pollutants are found in vital organs of late gestation human fetuses. Chemosphere 2021, 283, 131125. [Google Scholar] [CrossRef]
- Mamsen, L.S.; Björvang, R.D.; Mucs, D.; Vinnars, M.T.; Papadogiannakis, N.; Lindh, C.H.; Andersen, C.Y.; Damdimopoulou, P. Concentrations of perfluoroalkyl substances (PFASs) in human embryonic and fetal organs from first, second, and third trimester pregnancies. Environ. Int. 2019, 124, 482–492. [Google Scholar] [CrossRef]
- Gerona, R.R.; Woodruff, T.J.; Dickenson, C.A.; Pan, J.; Schwartz, J.M.; Sen, S.; Friesen, M.W.; Fujimoto, V.Y.; Hunt, P.A. Bisphenol-A (BPA), BPA glucuronide, and BPA sulfate in midgestation umbilical cord serum in a northern and central California population. Environ. Sci. Technol. 2013, 47, 12477–12485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandau, C.D.; Ayotte, P.; Dewailly, E.; Duffe, J.; Norstrom, R.J. Pentachlorophenol and hydroxylated polychlorinated biphenyl metabolites in umbilical cord plasma of neonates from coastal populations in Quebec. Environ. Health Perspect. 2002, 110, 411–417. [Google Scholar] [CrossRef]
- Golub, M.S.; Donald, J.M.; Reyes, J.A. Reproductive toxicity of commercial PCB mixtures: LOAELs and NOAELs from animal studies. Environ. Health Perspect. 1991, 94, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Tyl, R.W.; Myers, C.B.; Marr, M.C.; Thomas, B.F.; Keimowitz, A.R.; Brine, D.R.; Veselica, M.M.; Fail, P.A.; Chang, T.Y.; Seely, J.C.; et al. Three-generation reproductive toxicity study of dietary bisphenol A in CD Sprague-Dawley rats. Toxicol. Sci. 2002, 68, 121–146. [Google Scholar] [CrossRef]
- European Food Safety Authority; FitzGerald, R.; Van Loveren, H.; Civitella, C.; Castoldi, A.F.; Bernasconi, G. Assessment of new information on Bisphenol S (BPS) submitted in response to the Decision 1 under REACH Regulation (EC) No 1907/2006. EFSA Supporting Publ. 2020, 17, 1844E. [Google Scholar] [CrossRef]
- Blystone, C.R.; Kissling, G.E.; Bishop, J.B.; Chapin, R.E.; Wolfe, G.W.; Foster, P.M. Determination of the di-(2-ethylhexyl) phthalate NOAEL for reproductive development in the rat: Importance of the retention of extra animals to adulthood. Toxicol. Sci. 2010, 116, 640–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jiang, X.; Chen, B. Reproductive and developmental toxicity in F1 Sprague-Dawley male rats exposed to di-n-butyl phthalate in utero and during lactation and determination of its NOAEL. Reprod. Toxicol. 2004, 18, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Luebker, D.J.; Case, M.T.; York, R.G.; Moore, J.A.; Hansen, K.J.; Butenhoff, J.L. Two-generation reproduction and cross-foster studies of perfluorooctanesulfonate (PFOS) in rats. Toxicology 2005, 215, 126–148. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ma, R.; Dong, L.; Liu, L.; Zhou, G.; Tian, Z.; Zhao, Q.; Xia, T.; Zhang, S.; Wang, A. Autophagy impairment contributes to PBDE-47-induced developmental neurotoxicity and its relationship with apoptosis. Theranostics 2019, 9, 4375–4390. [Google Scholar] [CrossRef]
- Schantz, S.L.; Moshtaghian, J.; Ness, D.K. Spatial learning deficits in adult rats exposed to ortho-substituted PCB congeners during gestation and lactation. Fundam. Appl. Toxicol. 1995, 26, 117–126. [Google Scholar] [CrossRef]
- Hellwig, J.; van Ravenzwaay, B.; Mayer, M.; Gembardt, C. Pre- and postnatal oral toxicity of vinclozolin in Wistar and Long-Evans rats. Regul. Toxicol. Pharmacol. 2000, 32, 42–50. [Google Scholar] [CrossRef]
- Gillette, R.; Miller-Crews, I.; Nilsson, E.E.; Skinner, M.K.; Gore, A.C.; Crews, D. Sexually dimorphic effects of ancestral exposure to vinclozolin on stress reactivity in rats. Endocrinology 2014, 155, 3853–3866. [Google Scholar] [CrossRef] [Green Version]
- Reilly, M.P.; Kunkel, M.N.; Thompson, L.M.; Zentay, A.; Weeks, C.D.; Crews, D.; Cormack, L.K.; Gore, A.C. Effects of endocrine-disrupting chemicals on hypothalamic oxytocin and vasopressin systems. J. Exp. Zool. A Ecol. Integr. Physiol. 2022, 337, 75–87. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- La Merrill, M.A.; Vandenberg, L.N.; Smith, M.T.; Goodson, W.; Browne, P.; Patisaul, H.B.; Guyton, K.Z.; Kortenkamp, A.; Cogliano, V.J.; Woodruff, T.J.; et al. Consensus on the key characteristics of endocrine-disrupting chemicals as a basis for hazard identification. Nat. Rev. Endocrinol. 2020, 16, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Gao, L.; Flaws, J.A. Prenatal exposure to an environmentally relevant phthalate mixture disrupts reproduction in F1 female mice. Toxicol. Appl. Pharmacol. 2017, 318, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundakovic, M.; Gudsnuk, K.; Franks, B.; Madrid, J.; Miller, R.L.; Perera, F.P.; Champagne, F.A. Sex-specific epigenetic disruption and behavioral changes following low-dose in utero bisphenol A exposure. Proc. Natl. Acad. Sci. USA 2013, 110, 9956–9961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grønnestad, R.; Johanson, S.M.; Müller, M.H.B.; Schlenk, D.; Tanabe, P.; Krøkje, Å.; Jaspers, V.L.B.; Jenssen, B.M.; Ræder, E.M.; Lyche, J.L.; et al. Effects of an environmentally relevant PFAS mixture on dopamine and steroid hormone levels in exposed mice. Toxicol. Appl. Pharmacol. 2021, 428, 115670. [Google Scholar] [CrossRef] [PubMed]
- Hernandez Scudder, M.E.; Young, R.L.; Thompson, L.M.; Kore, P.; Crews, D.; Hofmann, H.A.; Gore, A.C. EDCs Reorganize Brain-Behavior Phenotypic Relationships in Rats. J. Endocr. Soc. 2021, 5, bvab021. [Google Scholar] [CrossRef] [PubMed]
- Hernandez Scudder, M.E.; Weinberg, A.; Thompson, L.; Crews, D.; Gore, A.C. Prenatal EDCs impair mate and odor preference and activation of the VMN in male and female rats. Endocrinology 2020, 161, bqaa124. [Google Scholar] [CrossRef]
- Wang, A.; Padula, A.; Sirota, M.; Woodruff, T.J. Environmental influences on reproductive health: The importance of chemical exposures. Fertil. Steril. 2016, 106, 905–929. [Google Scholar] [CrossRef] [Green Version]
- Ikezuki, Y.; Tsutsumi, O.; Takai, Y.; Kamei, Y.; Taketani, Y. Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. 2002, 17, 2839–2841. [Google Scholar] [CrossRef] [Green Version]
- Watkins, D.J.; Téllez-Rojo, M.M.; Ferguson, K.K.; Lee, J.M.; Solano-Gonzalez, M.; Blank-Goldenberg, C.; Peterson, K.E.; Meeker, J.D. In utero and peripubertal exposure to phthalates and BPA in relation to female sexual maturation. Environ. Res. 2014, 134, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Haines, D.A.; Murray, J. Human biomonitoring of environmental chemicals--early results of the 2007–2009 Canadian Health Measures Survey for males and females. Int. J. Hyg. Environ. Health 2012, 215, 133–137. [Google Scholar] [CrossRef]
- Li, N.; Papandonatos, G.D.; Calafat, A.M.; Yolton, K.; Lanphear, B.P.; Chen, A.; Braun, J.M. Gestational and childhood exposure to phthalates and child behavior. Environ. Int. 2020, 144, 106036. [Google Scholar] [CrossRef] [PubMed]
- Wickerham, E.L.; Lozoff, B.; Shao, J.; Kaciroti, N.; Xia, Y.; Meeker, J.D. Reduced birth weight in relation to pesticide mixtures detected in cord blood of full-term infants. Environ. Int. 2012, 47, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doremus-Fitzwater, T.L.; Varlinskaya, E.I.; Spear, L.P. Social and non-social anxiety in adolescent and adult rats after repeated restraint. Physiol. Behav. 2009, 97, 484–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandi, C.; Haller, J. Stress and the social brain: Behavioural effects and neurobiological mechanisms. Nat. Rev. Neurosci. 2015, 16, 290–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layton, A.C.; Sanseverino, J.; Gregory, B.W.; Easter, J.P.; Sayler, G.S.; Schultz, T.W. In vitro estrogen receptor binding of PCBs: Measured activity and detection of hydroxylated metabolites in a recombinant yeast assay. Toxicol. Appl. Pharmacol. 2002, 180, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Wrobel, M.; Kaminski, K.; Kotwica, J. In vitro effects of polychlorinated biphenyls (PCBs) on the contractility of bovine myometrium from the periovulatory stage of the estrous cycle. Reprod. Biol. 2005, 5, 303–319. [Google Scholar]
- Kavlock, R.; Cummings, A. Mode of action: Inhibition of androgen receptor function--vinclozolin-induced malformations in reproductive development. Crit. Rev. Toxicol. 2005, 35, 721–726. [Google Scholar] [CrossRef]
- Mylchreest, E.; Sar, M.; Cattley, R.C.; Foster, P.M. Disruption of androgen-regulated male reproductive development by di(n-butyl) phthalate during late gestation in rats is different from flutamide. Toxicol. Appl. Pharmacol. 1999, 156, 81–95. [Google Scholar] [CrossRef]
- Kirk, A.B.; Michelsen-Correa, S.; Rosen, C.; Martin, C.F.; Blumberg, B. PFAS and Potential Adverse Effects on Bone and Adipose Tissue Through Interactions With PPARγ. Endocrinology 2021, 162, bqab194. [Google Scholar] [CrossRef]
- Petrulis, J.R.; Bunce, N.J. Competitive behavior in the interactive toxicology of halogenated aromatic compounds. J. Biochem. Mol. Toxicol. 2000, 14, 73–81. [Google Scholar] [CrossRef]
- Song, J.; Li, Y.; Zhao, C.; Zhou, Q.; Zhang, J. Interaction of BDE-47 with nuclear receptors (NRs) based on the cytotoxicity: In vitro investigation and molecular interaction. Ecotoxicol. Environ. Saf. 2021, 208, 111390. [Google Scholar] [CrossRef] [PubMed]
- Behr, A.C.; Plinsch, C.; Braeuning, A.; Buhrke, T. Activation of human nuclear receptors by perfluoroalkylated substances (PFAS). Toxicol. Vitr. 2020, 62, 104700. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Wang, L.; Liang, Y.; Li, Z.; Feng, H.; Sun, Y.; Zhang, A.; Fu, J. Protonation state effects of estrogen receptor α on the recognition mechanisms by perfluorooctanoic acid and perfluorooctane sulfonate: A computational study. Ecotoxicol. Environ. Saf. 2019, 171, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Han, Y.; Zhao, Z.; Wu, Y.; Lu, Y.; Chen, G.; Jiang, J.; Qiu, L.; Gu, A.; Wang, X. Perfluorooctane sulfonate interferes with non-genomic estrogen receptor signaling pathway, inhibits ERK1/2 activation and induces apoptosis in mouse spermatocyte-derived cells. Toxicology 2021, 460, 152871. [Google Scholar] [CrossRef]
- Qin, W.P.; Cao, L.Y.; Li, C.H.; Guo, L.H.; Colbourne, J.; Ren, X.M. Perfluoroalkyl Substances Stimulate Insulin Secretion by Islet β Cells via G Protein-Coupled Receptor 40. Environ. Sci. Technol. 2020, 54, 3428–3436. [Google Scholar] [CrossRef]
- Mohajer, N.; Du, C.Y.; Checkcinco, C.; Blumberg, B. Obesogens: How They Are Identified and Molecular Mechanisms Underlying Their Action. Front. Endocrinol. 2021, 12, 780888. [Google Scholar] [CrossRef]
- Sharma, P.; Mandal, M.B.; Katiyar, R.; Singh, S.P.; Birla, H. A Comparative Study of Effects of 28-Day Exposure of Bisphenol A and Bisphenol S on Body Weight Changes, Organ Histology, and Relative Organ Weight. Int. J. Appl. Basic. Med. Res. 2021, 11, 214–220. [Google Scholar] [CrossRef]
- Tatsuta, N.; Kurokawa, N.; Nakai, K.; Suzuki, K.; Iwai-Shimada, M.; Murata, K.; Satoh, H. Effects of intrauterine exposures to polychlorinated biphenyls, methylmercury, and lead on birth weight in Japanese male and female newborns. Environ. Health Prev. Med. 2017, 22, 39. [Google Scholar] [CrossRef] [Green Version]
- Robledo, C.A.; Yeung, E.; Mendola, P.; Sundaram, R.; Maisog, J.; Sweeney, A.M.; Barr, D.B.; Louis, G.M. Preconception maternal and paternal exposure to persistent organic pollutants and birth size: The LIFE study. Environ. Health Perspect. 2015, 123, 88–94. [Google Scholar] [CrossRef]
- Gray, L.E., Jr.; Ostby, J.S.; Kelce, W.R. Developmental effects of an environmental antiandrogen: The fungicide vinclozolin alters sex differentiation of the male rat. Toxicol. Appl. Pharmacol. 1994, 129, 46–52. [Google Scholar] [CrossRef]
- Lopez-Rodriguez, D.; Franssen, D.; Heger, S.; Parent, A.S. Endocrine-disrupting chemicals and their effects on puberty. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101579. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, S.M.; Gore, A.C. Estrogenic environmental endocrine-disrupting chemical effects on reproductive neuroendocrine function and dysfunction across the life cycle. Rev. Endocr. Metab. Disord. 2007, 8, 143–159. [Google Scholar] [CrossRef]
- Sun, Y.; Fang, J.; Wan, Y.; Su, P.; Tao, F. Role of polygenic risk in susceptibility to accelerated pubertal onset following chronic stress exposure. Eur. J. Endocrinol. 2019, 181, 129–137. [Google Scholar] [CrossRef]
- Gur, R.E.; Moore, T.M.; Rosen, A.F.G.; Barzilay, R.; Roalf, D.R.; Calkins, M.E.; Ruparel, K.; Scott, J.C.; Almasy, L.; Satterthwaite, T.D.; et al. Burden of Environmental Adversity Associated With Psychopathology, Maturation, and Brain Behavior Parameters in Youths. JAMA Psychiatry 2019, 76, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.S.M.; Richardson, R. Early-life stress leads to sex-dependent changes in pubertal timing in rats that are reversed by a probiotic formulation. Dev. Psychobiol. 2019, 61, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Manzano Nieves, G.; Schilit Nitenson, A.; Lee, H.I.; Gallo, M.; Aguilar, Z.; Johnsen, A.; Bravo, M.; Bath, K.G. Early Life Stress Delays Sexual Maturation in Female Mice. Front. Mol. Neurosci. 2019, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, D.; Sun, Y. Adverse Childhood Experiences and Early Pubertal Timing Among Girls: A Meta-Analysis. Int. J. Environ. Res. Public Health 2019, 16, 2887. [Google Scholar] [CrossRef] [Green Version]
- Walley, S.N.; Krumm, E.A.; Yasrebi, A.; Wiersielis, K.R.; O’Leary, S.; Tillery, T.; Roepke, T.A. Maternal organophosphate flame-retardant exposure alters offspring feeding, locomotor and exploratory behaviors in a sexually-dimorphic manner in mice. J. Appl. Toxicol. 2021, 41, 442–457. [Google Scholar] [CrossRef]
- Kaur, S.; Kinkade, J.A.; Green, M.T.; Martin, R.E.; Willemse, T.E.; Bivens, N.J.; Schenk, A.K.; Helferich, W.G.; Trainor, B.C.; Fass, J.; et al. Disruption of global hypothalamic microRNA (miR) profiles and associated behavioral changes in California mice (Peromyscus californicus) developmentally exposed to endocrine disrupting chemicals. Horm. Behav. 2021, 128, 104890. [Google Scholar] [CrossRef]
- Wolstenholme, J.T.; Edwards, M.; Shetty, S.R.; Gatewood, J.D.; Taylor, J.A.; Rissman, E.F.; Connelly, J.J. Gestational exposure to bisphenol A produces transgenerational changes in behaviors and gene expression. Endocrinology 2012, 153, 3828–3838. [Google Scholar] [CrossRef] [Green Version]
- Hodes, G.E.; Pfau, M.L.; Purushothaman, I.; Ahn, H.F.; Golden, S.A.; Christoffel, D.J.; Magida, J.; Brancato, A.; Takahashi, A.; Flanigan, M.E.; et al. Sex Differences in Nucleus Accumbens Transcriptome Profiles Associated with Susceptibility versus Resilience to Subchronic Variable Stress. J. Neurosci. 2015, 35, 16362–16376. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L.; Epperson, C.N. Sex differences and stress across the lifespan. Nat. Neurosci. 2015, 18, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.M.; Zhang, J.J.; Huang, Y.B.; Zhao, Y.Z.; Sui, N. Peripubertal stress of male, but not female rats increases morphine-induced conditioned place preference and locomotion in adulthood. Dev. Psychobiol. 2019, 61, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.L.; Morano, R.L.; Ulrich-Lai, Y.M.; Myers, B.; Solomon, M.B.; Herman, J.P. Adolescent environmental enrichment prevents behavioral and physiological sequelae of adolescent chronic stress in female (but not male) rats. Stress 2018, 21, 464–473. [Google Scholar] [CrossRef]
- Kaplowitz, E.T.; Savenkova, M.; Karatsoreos, I.N.; Romeo, R.D. Somatic and Neuroendocrine Changes in Response to Chronic Corticosterone Exposure During Adolescence in Male and Female Rats. J. Neuroendocrinol. 2016, 28, 12336. [Google Scholar] [CrossRef]
- Adam, N.; Brusamonti, L.; Mhaouty-Kodja, S. Exposure of Adult Female Mice to Low Doses of di(2-ethylhexyl) Phthalate Alone or in an Environmental Phthalate Mixture: Evaluation of Reproductive Behavior and Underlying Neural Mechanisms. Environ. Health Perspect. 2021, 129, 17008. [Google Scholar] [CrossRef]
- Dombret, C.; Capela, D.; Poissenot, K.; Parmentier, C.; Bergsten, E.; Pionneau, C.; Chardonnet, S.; Hardin-Pouzet, H.; Grange-Messent, V.; Keller, M.; et al. Neural Mechanisms Underlying the Disruption of Male Courtship Behavior by Adult Exposure to Di(2-ethylhexyl) Phthalate in Mice. Environ. Health Perspect. 2017, 125, 097001. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.C.; Holley, A.M.; Crews, D. Mate choice, sexual selection, and endocrine-disrupting chemicals. Horm. Behav. 2018, 101, 3–12. [Google Scholar] [CrossRef]
- Patisaul, H.B.; Sullivan, A.W.; Radford, M.E.; Walker, D.M.; Adewale, H.B.; Winnik, B.; Coughlin, J.L.; Buckley, B.; Gore, A.C. Anxiogenic effects of developmental bisphenol A exposure are associated with gene expression changes in the juvenile rat amygdala and mitigated by soy. PLoS ONE 2012, 7, e43890. [Google Scholar] [CrossRef]
- Cao, J.; Joyner, L.; Mickens, J.A.; Leyrer, S.M.; Patisaul, H.B. Sex-specific Esr2 mRNA expression in the rat hypothalamus and amygdala is altered by neonatal bisphenol A exposure. Reproduction 2014, 147, 537–554. [Google Scholar] [CrossRef] [Green Version]
- Gillette, R.; Son, M.J.; Ton, L.; Gore, A.C.; Crews, D. Passing experiences on to future generations: Endocrine disruptors and transgenerational inheritance of epimutations in brain and sperm. Epigenetics 2018, 13, 1106–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatcher, K.M.; Willing, J.; Chiang, C.; Rattan, S.; Flaws, J.A.; Mahoney, M.M. Exposure to di-(2-ethylhexyl) phthalate transgenerationally alters anxiety-like behavior and amygdala gene expression in adult male and female mice. Physiol. Behav. 2019, 207, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Viau, V. Functional cross-talk between the hypothalamic-pituitary-gonadal and -adrenal axes. J. Neuroendocrinol. 2002, 14, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Rivest, S.; Rivier, C. Influence of the paraventricular nucleus of the hypothalamus in the alteration of neuroendocrine functions induced by intermittent footshock or interleukin. Endocrinology 1991, 129, 2049–2057. [Google Scholar] [CrossRef] [PubMed]








| Females | Males | ||||||
|---|---|---|---|---|---|---|---|
| Development and Physiology | Mean | SEM | Mean | SEM | p Value | Cohen’s d | Effect Size |
| P1 Body Weight | 6.20 | 0.07 | 6.55 | 0.07 | 0.120 | −0.50 | Medium |
| P84 Body Weight | 239 | 1.7 | 383 | 3.4 | <0.0001 | −5.42 | Large |
| P1 AGI | 1.06 | 0.01 | 2.08 | 0.01 | <0.0001 | −11.21 | Large |
| P14 AGI | 2.03 | 0.01 | 3.46 | 0.01 | <0.0001 | −12.09 | Large |
| Corticosterone | 821 | 43 | 472 | 31 | <0.0001 | 1.34 | Large |
| Light/Dark | Mean | SEM | Mean | SEM | p value | Cohen’s d | Effect Size |
| Distance | 24.35 | 0.41 | 19.76 | 0.40 | <0.0001 | 1.13 | Large |
| Freezing Episodes | 22.28 | 0.93 | 23.59 | 1.07 | 0.283 | −0.13 | Small |
| Time Freezing | 27.31 | 1.31 | 31.78 | 1.85 | 0.050 | −0.28 | Small |
| Light Time | 113 | 5.0 | 95 | 7.5 | 0.040 | 0.28 | Small |
| Light Freezing Episodes | 5.41 | 0.38 | 5.31 | 0.53 | 0.854 | 0.02 | Small |
| Light Time Freezing | 6.77 | 0.69 | 7.65 | 1.02 | 0.479 | −0.10 | Small |
| Dark Time | 186 | 5.0 | 203 | 7.5 | 0.045 | −0.27 | Small |
| Dark Freezing Episodes | 16.79 | 0.93 | 18.06 | 1.17 | 0.324 | −0.12 | Small |
| Dark Time Freezing | 20.28 | 1.21 | 23.61 | 1.80 | 0.115 | −0.22 | Small |
| Line Crossing | 16.66 | 0.59 | 11.50 | 0.79 | <0.0001 | 0.74 | Medium |
| Sociability | Mean | SEM | Mean | SEM | p value | Cohen’s d | Effect Size |
| Distance | 59.44 | 0.95 | 43.57 | 0.93 | <0.001 | 1.71 | Large |
| Time In Center | 97.52 | 2.82 | 128.16 | 5.22 | <0.001 | −0.73 | Medium |
| Time Near Social | 230 | 5.0 | 208 | 6.5 | 0.002 | 0.39 | Small |
| Avg Visit Time to Social | 16.05 | 0.60 | 18.49 | 0.71 | 0.016 | −0.38 | Small |
| Time Near Empty | 113 | 3.6 | 108 | 4.5 | 0.440 | 0.14 | Small |
| Avg Visit Time to Empty | 11.17 | 0.39 | 13.94 | 0.67 | <0.001 | −0.51 | Large |
| Social Pref Score | 0.67 | 0.01 | 0.65 | 0.01 | 0.255 | 0.14 | Small |
| Social Novelty | Mean | SEM | Mean | SEM | p value | Cohen’s d | Effect Size |
| Distance | 56.38 | 0.89 | 40.53 | 0.84 | <0.001 | 1.85 | Large |
| Time in Center | 136 | 3.8 | 172 | 6.1 | <0.001 | −0.73 | Medium |
| Time Near Familiar | 115 | 5.0 | 104 | 4.7 | 0.123 | 0.22 | Small |
| Avg Visit Time Familiar | 10.81 | 0.49 | 12.96 | 0.77 | 0.020 | −0.34 | Small |
| Time Near Novel | 164 | 5.6 | 140 | 5.2 | 0.002 | 0.45 | Small |
| Avg Visit Time Novel | 12.44 | 0.52 | 14.11 | 0.58 | 0.034 | −0.31 | Small |
| Social Novelty Score | 0.59 | 0.01 | 0.58 | 0.01 | 0.466 | 0.10 | Small |
| Mate Preference | Mean | SEM | Mean | SEM | p value | Cohen’s d | Effect Size |
| Distance | 67.36 | 0.92 | 49.42 | 0.88 | <0.001 | 2.00 | Large |
| Time Near Hormone | 185 | 5.5 | 244 | 6.7 | <0.001 | −0.97 | Large |
| Time Near NH | 115 | 3.2 | 113 | 4.0 | 0.751 | 0.05 | Small |
| Stim Explore Time Hormone | 132 | 4.7 | 190 | 6.6 | <0.001 | −1.03 | Large |
| Nose Touch Number Hormone | 11.52 | 0.73 | 17.76 | 1.01 | <0.001 | −0.71 | Medium |
| Nose Touch Time Hormone | 13.77 | 1.10 | 23.86 | 1.48 | <0.001 | −0.78 | Medium |
| Stim Explore Time NH | 74.98 | 2.93 | 76.10 | 3.39 | 0.802 | −0.04 | Small |
| Nose Touch Number NH | 6.00 | 0.53 | 9.54 | 0.72 | <0.001 | −0.56 | Medium |
| Nose Touch Time NH | 6.81 | 0.74 | 12.18 | 1.07 | <0.001 | −0.59 | Medium |
| Time Near Both Rats | 300 | 5.4 | 358 | 5.8 | <0.001 | −1.02 | Large |
| Mate Pref Score | 0.22 | 0.02 | 0.35 | 0.02 | <0.0001 | −0.59 | Medium |
| Gene Expression | Mean | SEM | Mean | SEM | p value | Cohen’s d | Effect Size |
| MeA Ar | 0.94 | 0.02 | 0.87 | 0.02 | 0.062 | 0.40 | Small |
| MeA Esr2 | 0.91 | 0.06 | 0.99 | 0.06 | 0.374 | −0.18 | Small |
| PVN Ar | 0.95 | 0.03 | 0.97 | 0.04 | 0.627 | −0.08 | Small |
| PVN Esr2 | 0.99 | 0.05 | 1.14 | 0.06 | 0.077 | −0.37 | Small |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gore, A.C.; Moore, T.; Groom, M.J.; Thompson, L.M. Prenatal Exposure to an EDC Mixture, NeuroMix: Effects on Brain, Behavior, and Stress Responsiveness in Rats. Toxics 2022, 10, 122. https://doi.org/10.3390/toxics10030122
Gore AC, Moore T, Groom MJ, Thompson LM. Prenatal Exposure to an EDC Mixture, NeuroMix: Effects on Brain, Behavior, and Stress Responsiveness in Rats. Toxics. 2022; 10(3):122. https://doi.org/10.3390/toxics10030122
Chicago/Turabian StyleGore, Andrea C., Tatum Moore, Matthew J. Groom, and Lindsay M. Thompson. 2022. "Prenatal Exposure to an EDC Mixture, NeuroMix: Effects on Brain, Behavior, and Stress Responsiveness in Rats" Toxics 10, no. 3: 122. https://doi.org/10.3390/toxics10030122
APA StyleGore, A. C., Moore, T., Groom, M. J., & Thompson, L. M. (2022). Prenatal Exposure to an EDC Mixture, NeuroMix: Effects on Brain, Behavior, and Stress Responsiveness in Rats. Toxics, 10(3), 122. https://doi.org/10.3390/toxics10030122