
Glycine Betaine Relieves Lead-Induced Hepatic and Renal Toxicity in Albino Rats

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals and Experiment Design
- The control (G1): rats received basal diet and drinking water.
- Pb-acetate treated(G2): rats have ingested Pb-acetate in drinking water daily with a dose of 50 mg/kg bw daily for 6 weeks.
- Pb-acetate and GB-treated (G3): rats received Pb-acetate at 50 mg/kg bw followed by GB at 250 mg/kg bw after 4 h in drinking water daily for 6 weeks.
2.3. Serum Samples Preparation
2.4. Quantitative Determination of Lipid Peroxidation
2.5. Determination of GSH-R Levels
2.6. Serum Biochemistry
2.6.1. Liver Function
2.6.2. Kidney Function
2.7. Serum Antioxidant Enzyme Assay
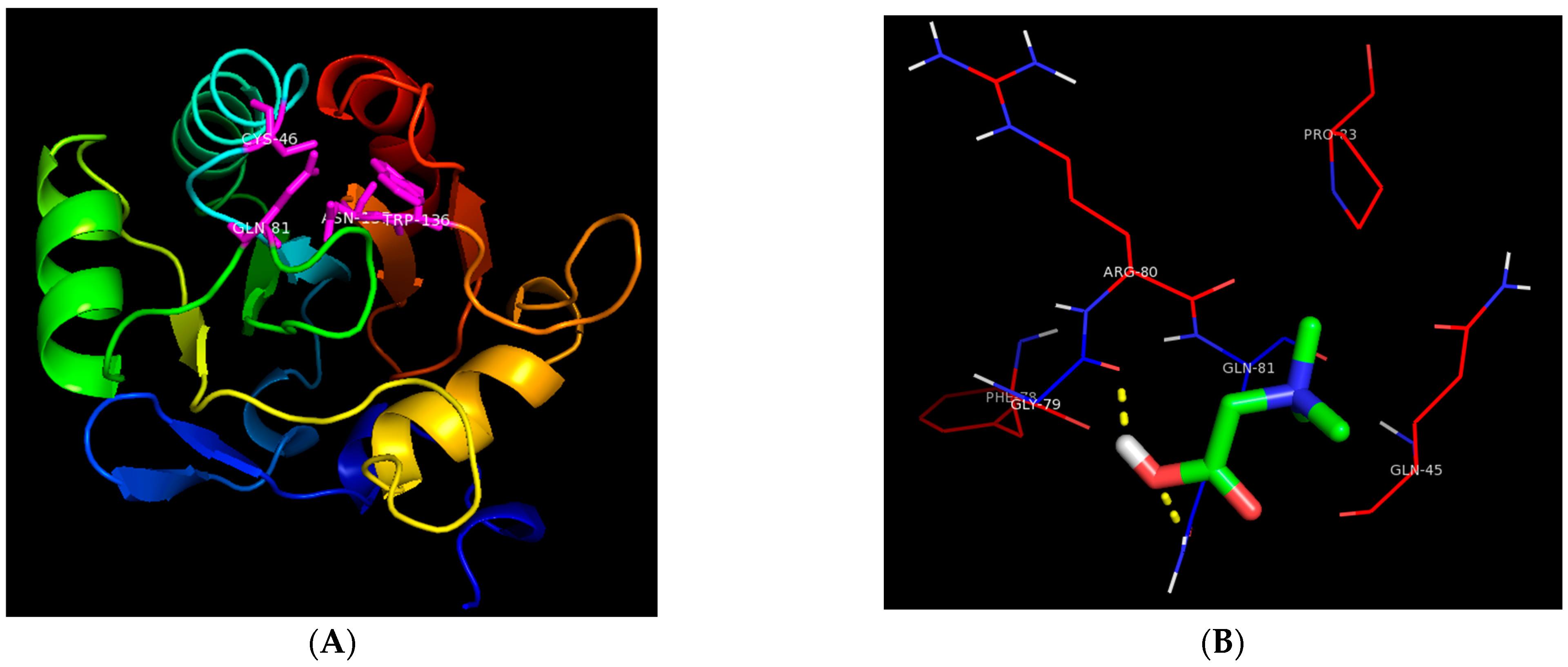
2.8. Molecular Docking of Glutathione Peroxidase
2.9. Statistical Analysis
3. Results
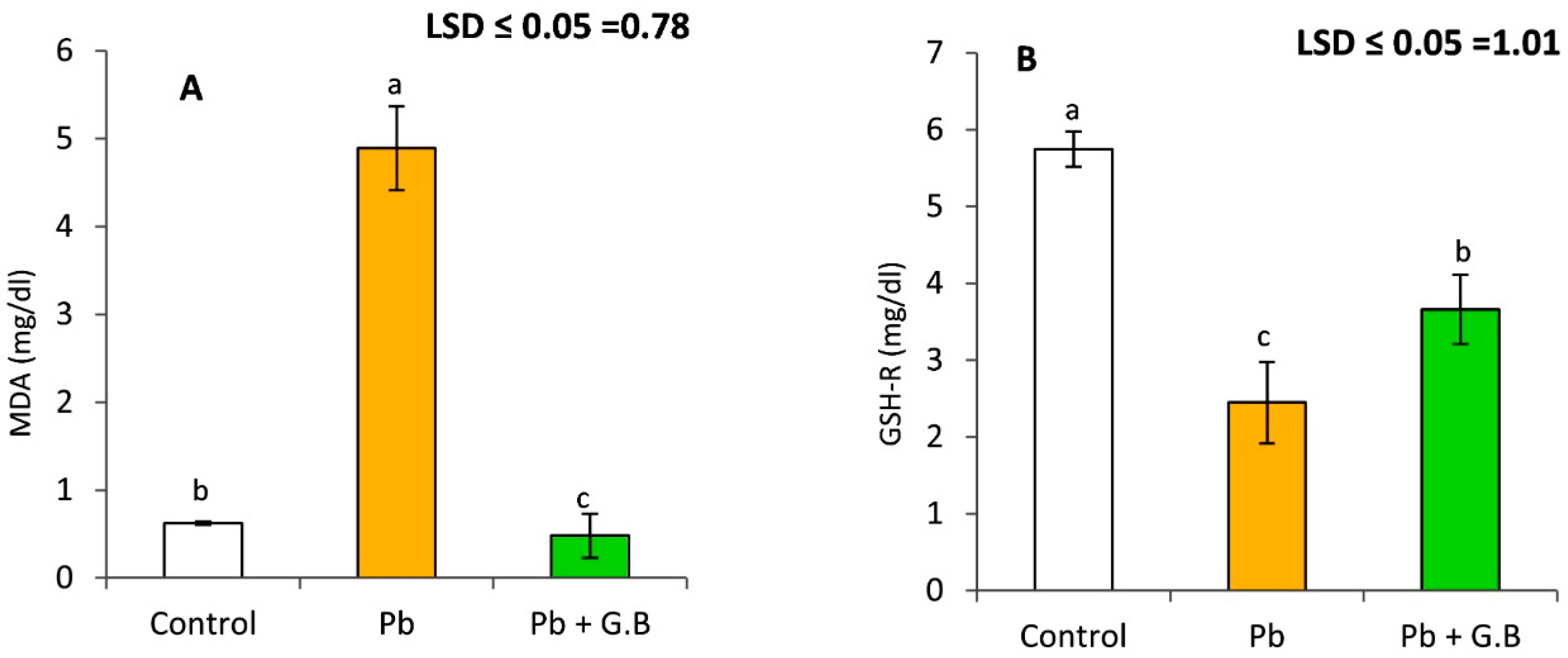
3.1. GB Mitigates Lipid Peroxidation and GSH-R in Pb-Exposed Albino Rats
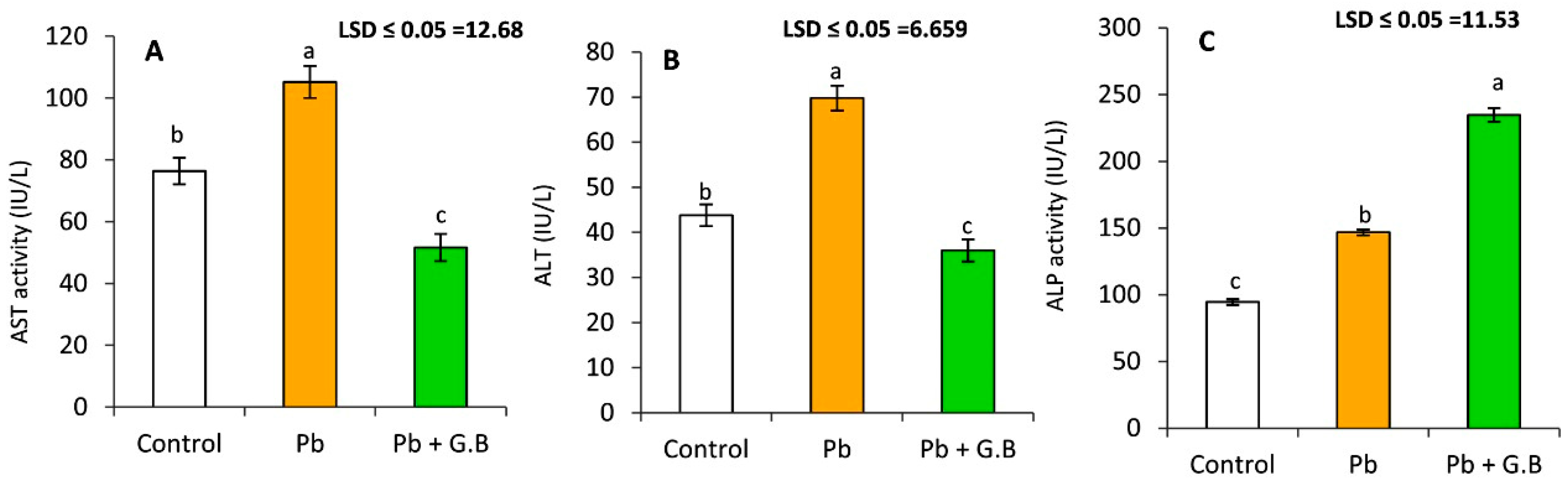
3.2. GB Protects the Liver against Pb-Induced Dysfunction
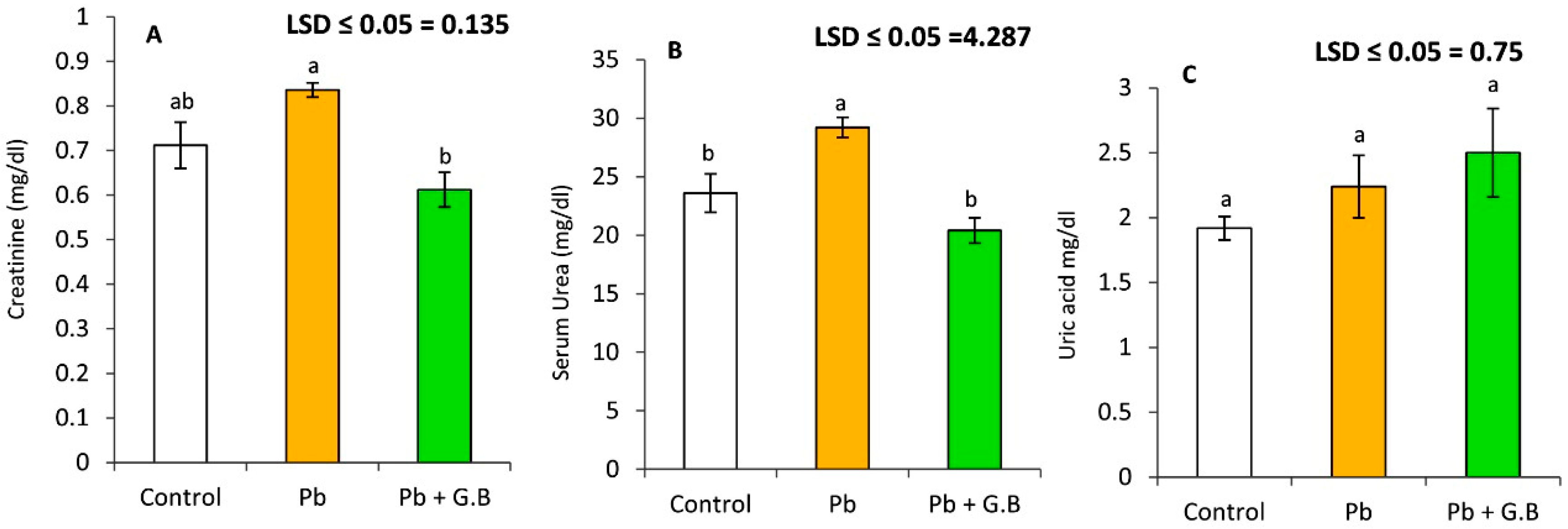
3.3. GB Improves the Kidney Function of Pb-Intoxicated Rats
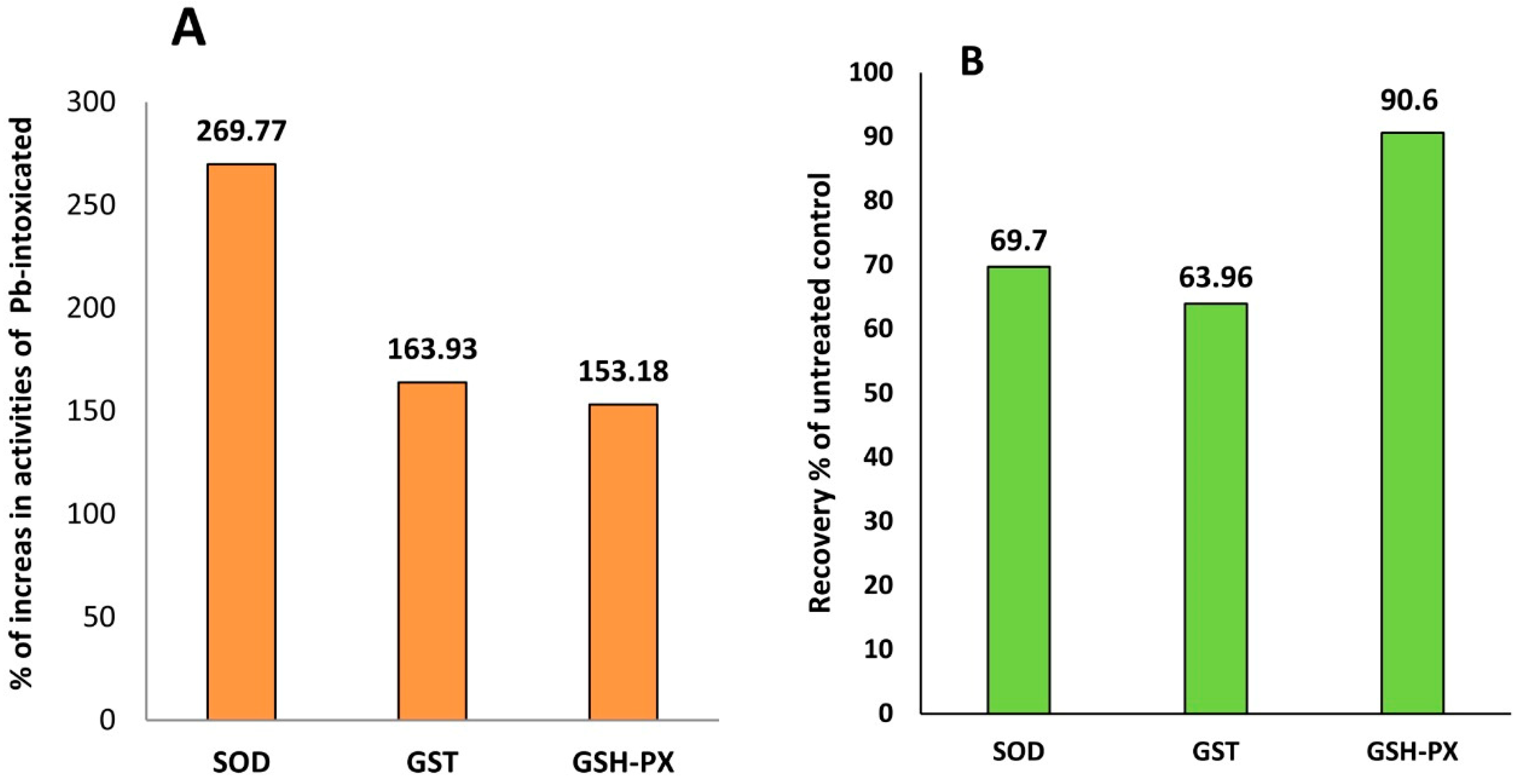
3.4. GB Enhances the Activity of Antioxidant Defense Enzymes in Pb-Intoxicated Rats
3.5. Molecular Interaction between GB and GSH-PX
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skerfving, I.S.; Bergdahl, A. Lead. In Handbook on the Toxicology of Metals; Nordberg, G.F., Fowler, B.A., Nordberg, M., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 2, pp. 911–967. [Google Scholar]
- Flora, S.J.S. Role of free radicals and antioxidants in health and disease. Cell. Mol. Biol. 2007, 53, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhusaini, A.; Fadda, L.; Hasan, I.H.; Zakaria, E.; Alenazi, A.M.; Mahmoud, A.M. Curcumin Ameliorates Lead-Induced Hepatotoxicity by Suppressing Oxidative Stress and Inflammation, and Modulating Akt/GSK-3β Signaling Pathway. Biomolecules 2019, 9, 703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B.; Gutteridge, J.M. The antioxidants of human extracellular fluids. Arch. Biochem. Biophys. 1990, 280, 1–8. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Pereira, B.; Bechara, E.J.H. Are free radicals involved in lead poisoning? Xenobiotica 1991, 21, 1085–1090. [Google Scholar] [CrossRef]
- Monteiro, H.P.; Abdalla, D.S.; Arcuri, A.S.; Bechara, E. Oxygen toxicity related to exposure to lead. Clin. Chem. 1995, 31, 1673–1676. [Google Scholar] [CrossRef]
- Monteiro, H.P.; Bechara, E.; Abdalla, D.S.P. Free radicals involvement in neurological porphyrias and lead poisoning. Mol. Cell. Biochem. 1991, 103, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Ribarov, S.R.; Bochev, P.G. Lead-hemoglobin interaction as a possible source of reactive oxygen species—A chemiluminescent study. Arch. Biochem. Biophys. 1982, 213, 288–292. [Google Scholar] [CrossRef]
- Hsu, P.-C.; Liu, M.-Y.; Hsu, C.-C.; Chen, L.-Y.; Guo, Y.L. Lead exposure causes generation of reactive oxygen species and functional impairment in rat sperm. Toxicology 1997, 122, 133–143. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abdel-Rahman, H.G.; Dessouki, A.A.; El-Far, A.H.; Khodeer, D.M.; Bin-Jumah, M.; Alhader, M.S.; Alkahtani, S.; Aleya, L. Impact of Garlic (Allium Sativum) Oil on Cisplatin-Induced Hepatorenal Biochemical and Histopathological Alterations in Rats. Sci. Total Environ. 2020, 710, 136338. [Google Scholar] [CrossRef]
- Lawton, L.J.; Donaldson, W.E. Lead-induced tissue fatty acid alterations and lipid peroxidation. Biol. Trace Elem. Res. 1991, 28, 83–97. [Google Scholar] [CrossRef]
- Sandhir, R.; Julka, D.; Gill, K. Lipoperoxidative Damage on Lead Exposure in Rat Brain and its Implications on Membrane Bound Enzymes. Pharmacol. Toxicol. 1994, 74, 66–71. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.A.; Ibrahim, A.K.; Elfaky, M.A.; Habib, E.S.; Mahamed, M.I.; Mehanna, E.T.; Darwish, K.M.; Khodeer, D.M.; Ahmed, S.A.; Elhady, S.S. Antioxidant and Anti-Inflammatory Activity of Cynanchum acutum L. Isolated Flavonoids Using Experimentally Induced Type 2 Diabetes Mellitus: Biological and In Silico Investigation for NF-ΚB Pathway/MiR-146a Expression Modulation. Antioxidants 2021, 10, 1713. [Google Scholar] [CrossRef]
- Elkazzaz, S.K.; Khodeer, D.M.; El Fayoumi, H.M.; Moustafa, Y.M. Role of Sodium Glucose Cotransporter Type 2 Inhibitors Dapagliflozin on Diabetic Nephropathy in Rats; Inflammation, Angiogenesis and Apoptosis. Life Sci. 2021, 280, 119018. [Google Scholar] [CrossRef] [PubMed]
- Mamdouh, A.M.; Khodeer, D.M.; Tantawy, M.A.; Moustafa, Y.M. In-Vitro and In-Vivo Investigation of Amygdalin, Metformin, and Combination of Both against Doxorubicin on Hepatocellular Carcinoma. Life Sci. 2021, 285, 119961. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Dogan, Z.; Kanak, E.G.; Atli, G.; Canli, M. Effects of Heavy Metals (Cd, Cu, Cr, Pb, Zn) on Fish Glutathione Metabolism. Environ. Sci. Pollut. Res. 2015, 22, 3229–3237. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Kraemer, R. Coping with osmotic challenges: Osmoregulation through accumulation and release of compatible solutes in bacteria. In Bacterial Stress Responses; Storz, G., Hengge-Aronis, R.Ž., Eds.; ASM Press: Washington, DC, USA, 2000; pp. 79–97. [Google Scholar]
- De Zwart, F.; Slow, S.; Payne, R.; Lever, M.; George, P.; Gerrard, J.; Chambers, S. Glycine betaine and glycine betaine analogues in common foods. Food Chem. 2003, 83, 197–204. [Google Scholar] [CrossRef]
- Filipčev, B.; Kojić, J.; Krulj, J.; Bodroža-Solarov, M.; Ilić, N. Betaine in Cereal Grains and Grain-Based Products. Foods 2018, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Craig, S.A.S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Burg, M.B.; Ferraris, J.D. Intracellular Organic Osmolytes: Function and Regulation. J. Biol. Chem. 2008, 283, 7309–7313. [Google Scholar] [CrossRef] [Green Version]
- Ueland, P.M. Choline and betaine in health and disease. J. Inherit. Metab. Dis. 2011, 34, 3–15. [Google Scholar] [CrossRef]
- Go, E.K.; Jung, K.J.; Kim, J.M.; Lim, H.; Lim, H.K.; Yu, B.P.; Chung, H.Y. Betaine Modulates Age-Related NF-κB by Thiol-Enhancing Action. Biol. Pharm. Bull. 2007, 30, 2244–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harisa, G.I. Oxidative Stress and Paraoxonase Activity in Experimental Selenosis: Effects of Betaine Administration. Biol. Trace Elem. Res. 2013, 152, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Kim, S.J.; Kwon, D.Y.; Ahn, C.W.; Kim, Y.S.; Choi, D.W.; Kim, Y.C. Alleviation of alcoholic liver injury by betaine involves an enhancement of antioxidant defense via regulation of sulfur amino acid metabolism. Food Chem. Toxicol. 2013, 62, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Mcconnachie, L.A.; Mohar, I.; Hudson, F.N.; Ware, C.B.; Ladiges, W.C.; Fernandwz, C.; Charrerton-Kirchmeir, S.; White, C.C.; Pierce, R.H.; Kavanagh, T.J. Glutamate cysteine ligase modifier subunit deficiency and gender as determinants of acetaminophen-induced hepatotoxicity in mice. Toxicol. Sci. 2007, 99, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Shengc, Y.; Jiang, P.; Ji, L.; Xia, Y.; Min, Y.; Wang, Z. The gender-dependent difference of liver GSH antioxidant system in mice and its influence on isoline- induced liver injury. Toxicology 2011, 280, 61–69. [Google Scholar] [CrossRef]
- Tiidus, P.M. Estrogen and gender effects on muscle damage, inflammation, and oxidative stress. Can. J. Appl. Physiol. 2000, 25, 274–287. [Google Scholar] [CrossRef]
- Jegede, A.I.; Offor, U.; Azu, O.O.; Azu Akinloye, O. Red Palm Oil attenuates lead acetate induced testicular damage in adult male Sprague-Dawley rats. Evid. Based Complement. Altern. Med. 2015, 2015, 130261. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Crowley, C.; Gillham, B.; Thorn, M.B. A direct enzymic method for the determination of reduced glutathione in blood and other tissues. Biochem. Med. 1975, 13, 287–292. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Tietz, N.W.; Burtis, C.A.; Duncan, P.; Ervin, K.; Petitclerc, C.J.; Rinker, A.D.; Zygowicz, E.R. A reference method for measurement of alkaline phosphatase activity in human serum. Clin. Chem. 1983, 29, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Wybenga, D.R.; Di Giorgio, J.; Pileggi, V.J. Manual and automated methods for urea nitrogen measurement in whole serum. Clin. Chem. 1971, 17, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Artiss, J.D.; Entwistle, W.M. The application of a sensitive uricase-peroxidase coupled reaction to a centrifugal fast analyser for the determination of uric acid. Clin. Chim. Acta 1981, 116, 301–309. [Google Scholar] [CrossRef]
- Bowers, L.D.; Wong, E.T. Kinetic serum creatinine assays. II. A critical evaluation and review. Clin. Chem. 1980, 26, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K. Serum lipid peroxide in cerebrovascular disorders determined by a new colorimetric method. Clin. Chim. Acta 1978, 90, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wilce MC, J.; Parker, M.W. Structure and function of glutathione S-transferases. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1994, 1205, 1–18. [Google Scholar] [CrossRef]
- Wendel, A. Glutathione peroxidase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1981; Volume 77, pp. 325–333. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Janowski, R.; Scanu, S.; Niessing, D.; Madl, T. Crystal and solution structural studies of mouse phospholipid hydroperoxide glutathione peroxidase 4. Acta Crystallogr. 2016, 72, 743–749. [Google Scholar] [CrossRef] [Green Version]
- Okada, T.; Kawakami, S.; Nakamura, Y.; Han, K.-H.; Ohba, K.; Aritsuka, T.; Uchino Shimada, K.-I.; Sekikawa, M.; Ishii, H.; Fukushima, M. Amelioration of D- galactosamine-induced acute liver injury in rats by dietary supplementation with betaine derived from sugar beet molasses. Biosci. Biotechnol. Biochem. 2011, 75, 1335–1341. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yang, Z.; Lin, L.; Zhao, Z.; Liu, Z.; Liu, X. Protective effect of naringenin against lead-induced oxidative stress in rats. Biol. Trace Elem. Res. 2012, 146, 354–359. [Google Scholar] [CrossRef]
- Gürer, H.; Özgünes, H.; Neal, R.; Spitz, D.R.; Erçal, N. Antioxidant effects of N-acetylcysteine and succimer in red blood cells from lead-exposed rats. Toxicology 1998, 128, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Ercal, N.; Treeratphan, P.; Hammond, T.C.; Matthews, R.H.; Grannemann, N.H.; Spitz, D.R. In vivo indices of oxidative stress in lead-exposed C57BL/6 mice are reduced by treatment with meso-2, 3-dimercaptosuccinic acid or N-acetylcysteine. Free Radic. Biol. Med. 1996, 21, 157–161. [Google Scholar] [CrossRef]
- Sivaprasad, R.; Nagaraj, M.; Varalakshmi, P. Combined efficacies of lipoic acid and 2,3-dimercaptosuccinic acid against lead-induced lipid peroxidation in rat liver. J. Nutr. Biochem. 2004, 15, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, M.; Siddiqui, M.K.J. Low Level Lead Exposure and Oxidative Stress: Current Opinions. Clin. Chim. Acta 2007, 383, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Adebisi, O.A.; Agbaje, W.B.; Adewale, O.O. Modulatory efficacy of Punica granatum L. powder ethanol extract (PLEE) on lead acetate-induced hepatic and renal toxicity. Clin. Phytosci. 2022, 8, 6. [Google Scholar] [CrossRef]
- Ahamed, M.; Verma, S.; Kumar, A.; Siddiqui, M.K.J. Environmental exposure to lead and its correlation with biochemical indices in children. Sci. Total Environ. 2005, 346, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.C.; Hsu, C.C.; Liu, M.Y.; Chen, L.Y.; Guo, Y.L. Lead-induced changes in spermatozoa function and metabolism. J. Toxicol. Environ. Health 1998, 55, 45–64. [Google Scholar] [CrossRef]
- Tandon, S.K.; Chatterjee, M.; Bhargava, A.; Shukla, V.; Bihari, V. Lead poisoning in Indian silver refiners. Sci. Total Environ. 2001, 281, 177–182. [Google Scholar] [CrossRef]
- Adonaylo, V.N.; Oteiza, P.I. Pb2+ promotes lipid peroxidation and alteration in membrane physical properties. Toxicology 1999, 132, 19–32. [Google Scholar] [CrossRef]
- Mabrouk, A.; Cheikh, H.B. Thymoquinone ameliorates lead-induced suppression of the antioxidant system in rat kidneys. Libyan J. Med. 2016, 11, 31018. [Google Scholar] [CrossRef]
- Tsai, M.; Chen, C.; Pan, Y.; Wang, S.; Mersmann, H.; Ding, S. Alleviation of carbon- tetrachloride-induced liver injury and fibrosis by betaine supplementation in chickens. Evid. Based Complement. Altern. Med. 2015, 2015, 725379. [Google Scholar] [CrossRef] [Green Version]
- Bidanchi, R.M.; Lalrindika, L.; Khushboo, M.; Bhanushree, B.; Dinata, R.; Das, M.; Gurusubramanian, G. Antioxidative, anti-inflammatory and anti-apoptotic action of ellagic acid against lead acetate induced testicular and hepato-renal oxidative damages and pathophysiological changes in male Long Evans rats. Environ. Pollut. 2022, 302, 119048. [Google Scholar] [CrossRef] [PubMed]
- Balkan, J.; Öztezcan, S.; Küçük, M.; Çevikbaş, U.; Koçak-Toker, N.; Uysal, M. The effect of betaine treatment on triglyceride levels and oxidative stress in the liver of ethanol-treated guinea pigs. Exp. Toxicol. Pathol. 2004, 55, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Erman, F.; Balkan, J.; Cevikbaş, U.; Koçak-Toker, N.; Uysal, M. Betaine or taurine administration prevents fibrosis and lipid peroxidation induced by rat liver by ethanol plus carbon tetrachloride intoxication. Amino Acids 2004, 27, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.F.H.; El-Sayed, N.M.; Khodeer, D.M.; Ahmed, A.A.M.; Hanna, P.A.; Moustafa, Y.M.A. Nano Selenium Ameliorates Oxidative Stress and Inflammatory Response Associated with Cypermethrin-Induced Neurotoxicity in Rats. Ecotoxicol. Environ. Saf. 2020, 195, 110479. [Google Scholar] [CrossRef]
- Day, C.; Kempson, S. Betaine chemistry, roles, and potential use in liver disease. Biochim. Biophys. Acta 2016, 1860, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K. Dose-dependent Effects of Betaine on Hepatic Metabolism of Sulfur Amino Acids in Mice. Yakhak Hoeji 2009, 53, 69–73. [Google Scholar]
- Wang, X.L.; Rainwater, D.L.; Mahaney, M.C.; Stocker, R. Co-supplementation with vitamin E and coenzyme Q10 reduces circulating markers of inflammation in baboons. Am. J. Clin. Nutr. 2004, 80, 649–655. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | SOD | GSTs | GSH-Px |
|---|---|---|---|
| G1 (Control) | 3.147 ± 0.23 a | 4.074 ± 0.16 a | 6.794 ± 0.19 a |
| G2 (lead acetate) | 0.814 ± 0.03 d | 0.610 ± 0.01 d | 4.020 ± 0.71 b |
| G3 (lead acetate + GB) | 2.196 ± 0.09 b | 2.606 ± 0.27 b | 6.158 ± 0.07 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelrazek, F.; Salama, D.A.; Alharthi, A.; Asiri, S.A.; Khodeer, D.M.; Qarmush, M.M.; Mobasher, M.A.; Ibrahim, M. Glycine Betaine Relieves Lead-Induced Hepatic and Renal Toxicity in Albino Rats. Toxics 2022, 10, 271. https://doi.org/10.3390/toxics10050271
Abdelrazek F, Salama DA, Alharthi A, Asiri SA, Khodeer DM, Qarmush MM, Mobasher MA, Ibrahim M. Glycine Betaine Relieves Lead-Induced Hepatic and Renal Toxicity in Albino Rats. Toxics. 2022; 10(5):271. https://doi.org/10.3390/toxics10050271
Chicago/Turabian StyleAbdelrazek, Farid, Dawlat A. Salama, Afaf Alharthi, Saeed A. Asiri, Dina M. Khodeer, Moath M. Qarmush, Maysa A. Mobasher, and Mervat Ibrahim. 2022. "Glycine Betaine Relieves Lead-Induced Hepatic and Renal Toxicity in Albino Rats" Toxics 10, no. 5: 271. https://doi.org/10.3390/toxics10050271
APA StyleAbdelrazek, F., Salama, D. A., Alharthi, A., Asiri, S. A., Khodeer, D. M., Qarmush, M. M., Mobasher, M. A., & Ibrahim, M. (2022). Glycine Betaine Relieves Lead-Induced Hepatic and Renal Toxicity in Albino Rats. Toxics, 10(5), 271. https://doi.org/10.3390/toxics10050271