
Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants

Abstract
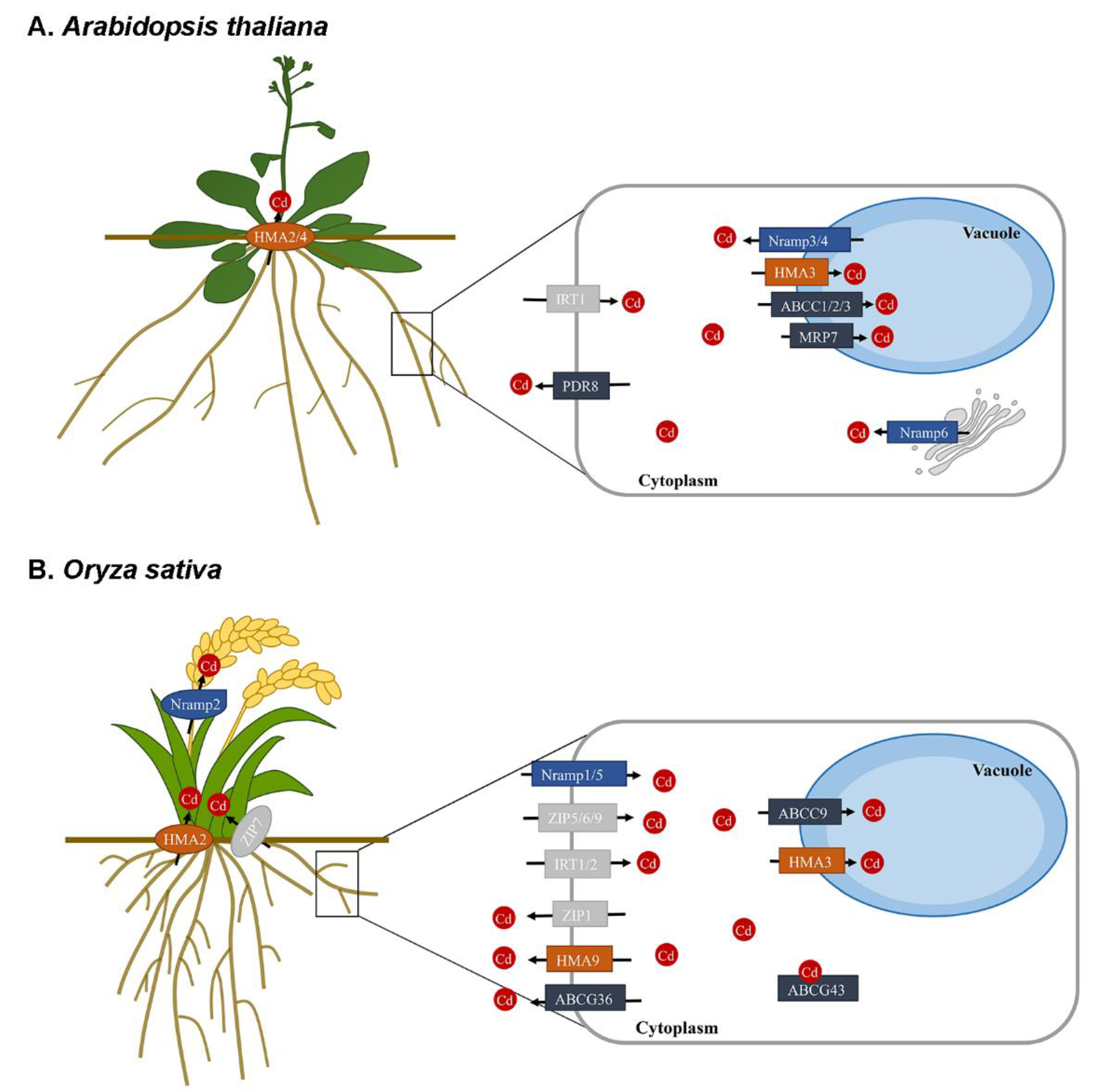
:1. Introduction
2. Natural Resistance-Associated Macrophage Proteins
3. Heavy Metal Transporting ATPases

{kind=link}
| Plant Species | Genes | Expression Sites | Subcellular Location | Function | References |
|---|---|---|---|---|---|
| Arabidopsis thaliana | AtHMA2 | - | Plasma membrane | Cd translocation | [61,62] |
| AtHMA3 | Root apex | Tonoplast | Cd sequestration | [63,64] | |
| AtHMA4 | tissues surrounding the root vascular vessels | Plasma membrane | Cd translocation | [61,65,66,67] | |
| Oryza sativa L | OsHMA2 | in the mature zone of the roots at the vegetative stage | Plasma membrane | Cd translocation | [68,69,70] |
| OsHMA3 | Roots | Tonoplast | Cd sequestration | [71,72,73,74,75,76] | |
| OsHMA9 | vascular bundles and anthers | Plasma membrane | Cd efflux | [77] | |
| Triticum aestivum L. | TaHMA2 | Nodes | Plasma membrane | Cd translocation | [78] |
| Glycine max | GmHAM3w | Roots | Endoplasmic reticulum (ER) | Cd sequestration | [79] |
| Sedum plumbizincicola | SpHMA1 | Leaves | Chloroplast envelope | Cd efflux | [80] |
| SpHMA3 | Leaves | Tonoplast | Cd sequestration | [81] | |
| Sedum alfredii Hance | SaHMA3 | Shoots | Tonoplast | Cd sequestration | [82] |
| Thlaspi caerulescens | TcHMA3 | Roots and shoots | Tonoplast | Cd sequestration | [83] |
| TcHMA4 | Roots | - | [84] | ||
| Brassica juncea | BjHMA4 | Roots, stems and leaves | Cytosol | Cd translocation | [85] |
| Iris lactea | IlHMA2 | Roots | Plasma membrane | Cd translocation | [86] |
| Populus tomentosa Carr. | PtoHMA5 | - | - | Cd translocation | [87] |
4. ATP-Binding Cassette
5. Zinc- and Iron-Regulated Transporter Proteins
6. Yellow Stripe-Like Proteins
7. Conclusions and Further Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sarwar, N.; Saifullah; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Sanita, L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Li, Y.; Kuang, H.; Hu, C.; Ge, G. Source Apportionment of Heavy Metal Pollution in Agricultural Soils around the Poyang Lake Region Using UNMIX Model. Sustainability 2021, 13, 5272. [Google Scholar] [CrossRef]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress and responses in plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Edelstein, M.; Ben-Hur, M. Heavy metals and metalloids: Sources, risks and strategies to reduce their accumulation in horticultural crops. Sci. Hortic. 2018, 234, 431–444. [Google Scholar] [CrossRef]
- Inaba, T.; Kobayashi, E.; Suwazono, Y.; Uetani, M.; Oishi, M.; Nakagawa, H.; Nogawa, K. Estimation of cumulative cadmium intake causing Itai-itai disease. Toxicol. Lett. 2005, 159, 192–201. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, S.; Khan, A.; Alam, M. Soil contamination with cadmium, consequences and remediation using organic amendments. Sci. Total Environ. 2017, 601–602, 1591–1605. [Google Scholar] [CrossRef]
- Yang, Q.; Li, Z.; Lu, X.; Duan, Q.; Huang, L.; Bi, J. A review of soil heavy metal pollution from industrial and agricultural regions in China: Pollution and risk assessment. Sci. Total Environ. 2018, 642, 690–700. [Google Scholar] [CrossRef]
- Zhao, F.J.; Ma, Y.; Zhu, Y.G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef]
- Wang, Y.; Duan, X.; Wang, L. Spatial distribution and source analysis of heavy metals in soils influenced by industrial enterprise distribution: Case study in Jiangsu Province. Sci. Total Environ. 2020, 710, 134953. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, H.; Tao, S. The Challenges and Solutions for Cadmium-contaminated Rice in China: A Critical Review. Environ. Int. 2016, 92–93, 515–532. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Kopittke, P.M.; Zhao, F.J. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar] [CrossRef]
- Zheng, S.; Wang, Q.; Yuan, Y.; Sun, W. Human health risk assessment of heavy metals in soil and food crops in the Pearl River Delta urban agglomeration of China. Food Chem. 2020, 316, 126213. [Google Scholar] [CrossRef]
- Song, Y.; Jin, L.; Wang, X. Cadmium absorption and transportation pathways in plants. Int. J. Phytoremediat. 2017, 19, 133–141. [Google Scholar] [CrossRef]
- Li, H.; Luo, N.; Li, Y.W.; Cai, Q.Y.; Li, H.Y.; Mo, C.H.; Wong, M.H. Cadmium in rice: Transport mechanisms, influencing factors, and minimizing measures. Environ. Pollut. 2017, 224, 622–630. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculik, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Z.; Tu, C.; Wu, L.H.; Peijnenburg, W.J.; Ebbs, S.; Luo, Y.M. Pathways of root uptake and membrane transport of Cd(2+) in the zinc/cadmium hyperaccumulating Plant Sedum plumbizincicola. Environ. Toxicol. Chem. 2017, 36, 1038–1046. [Google Scholar] [CrossRef]
- Chen, X.; Ouyang, Y.; Fan, Y.; Qiu, B.; Zhang, G.; Zeng, F. The pathway of transmembrane cadmium influx via calcium-permeable channels and its spatial characteristics along rice root. J. Exp. Bot. 2018, 69, 5279–5291. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.L.; Tian, S.K.; Yang, X.E.; Li, T.Q.; He, Z.L. Cadmium uptake and xylem loading are active processes in the hyperaccumulator Sedum alfredii. J. Plant Physiol. 2009, 166, 579–587. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Roles of root and shoot tissues in transport and accumulation of cadmium, lead, nickel, and strontium. Russ. J. Plant Physiol. 2011, 55, 1–22. [Google Scholar] [CrossRef]
- Mendoza-Cozatl, D.G.; Butko, E.; Springer, F.; Torpey, J.W.; Komives, E.A.; Kehr, J.; Schroeder, J.I. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation. Plant J. 2008, 54, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Tian, S.; Foyer, C.H.; Hou, D.; Wang, H.; Zhou, W.; Liu, T.; Ge, J.; Lu, L.; Lin, X. Efficient phloem transport significantly remobilizes cadmium from old to young organs in a hyperaccumulator Sedum alfredii. J. Hazard. Mater. 2019, 365, 421–429. [Google Scholar] [CrossRef]
- Hall, J.L.; Williams, L.E. Transition metal transporters in plants. J. Exp. Bot. 2003, 54, 2601–2613. [Google Scholar] [CrossRef] [PubMed]
- Socha, A.L.; Guerinot, M.L. Mn-euvering manganese: The role of transporter gene family members in manganese uptake and mobilization in plants. Front. Plant Sci. 2014, 5, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomine, S.; Wang, R.; Ward, J.M.; Crawford, N.M.; Crawford, J.I. Cadmium and iron transport by members of a Plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc. Natl. Acad. Sci. USA 2000, 97, 4991–4996. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, L.; Zheng, L.; Wang, Y.; Chen, X.; Zhang, W. A Functional Study Identifying Critical Residues Involving Metal Transport Activity and Selectivity in Natural Resistance-Associated Macrophage Protein 3 in Arabidopsis thaliana. Int. J. Mol. Sci. 2018, 19, 1430. [Google Scholar] [CrossRef] [Green Version]
- Thomine, S.; Lelievre, F.; Debarbieux, E.; Schroeder, J.I.; Barbier-Brygoo, H. AtNRAMP3, a multispecic vacuolar metal transporter involved in Plant responses to iron deciency. Plant J. 2003, 34, 685–695. [Google Scholar] [CrossRef]
- Pottier, M.; Oomen, R.; Picco, C.; Giraudat, J.; Scholz-Starke, J.; Richaud, P.; Carpaneto, A.; Thomine, S. Identification of mutations allowing Natural Resistance Associated Macrophage Proteins (NRAMP) to discriminate against cadmium. Plant J. 2015, 83, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Lanquar, V.; Lelièvre, F.; Barbier-Brygoo, H.; Thomine, S. Regulation and function of AtNRAMP4 metal transporter protein. Soil Sci. Plant Nutr. 2004, 50, 1141–1150. [Google Scholar] [CrossRef]
- Cailliatte, R.; Lapeyre, B.; Briat, J.F.; Mari, S.; Curie, C. The NRAMP6 metal transporter contributes to cadmium toxicity. Biochem. J. 2009, 422, 217–228. [Google Scholar] [CrossRef]
- Tiwari, M.; Sharma, D.; Dwivedi, S.; Singh, M.; Tripathi, R.D.; Trivedi, P.K. Expression in Arabidopsis and cellular localization reveal involvement of rice NRAMP, OsNRAMP1, in arsenic transport and tolerance. Plant Cell Environ. 2014, 37, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.D.; Huang, S.; Yamaji, N.; Zhang, W.; Ma, J.F.; Zhao, F.J. OsNRAMP1 transporter contributes to cadmium and manganese uptake in rice. Plant Cell Environ. 2020, 43, 2476–2491. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, W.; Zhang, S.; Yang, T.; Liu, Q.; Dong, J.; Fu, H.; Mao, X.; Liu, B. Genome-wide association study and candidate gene analysis of rice cadmium accumulation in grain in a diverse rice collection. Rice 2018, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.-D.; Xie, Y.; Zhang, H.; Zhang, S.; Zhao, F.-J. The vacuolar transporter OsNRAMP2 mediates Fe remobilization during germination and affects Cd distribution to rice grain. Plant Soil 2022, 1–17. [Google Scholar] [CrossRef]
- Chang, J.D.; Huang, S.; Konishi, N.; Wang, P.; Chen, J.; Huang, X.Y.; Ma, J.F.; Zhao, F.J. Overexpression of the manganese/cadmium transporter OsNRAMP5 reduces cadmium accumulation in rice grain. J. Exp. Bot. 2020, 71, 5705–5715. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; et al. Characterizing the role of rice NRAMP5 in Manganese, Iron and Cadmium Transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Wang, C.; Cheng, Y.; Kang, H.; Fan, X.; Sha, L.; Zhang, H.; Zeng, J.; Zhou, Y.; Wang, Y. Cloning and Characterization of TpNRAMP3, a Metal Transporter from Polish Wheat (Triticum polonicum L.). Front. Plant Sci. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Peng, F.; Wang, C.; Zhu, J.; Zeng, J.; Kang, H.; Fan, X.; Sha, L.; Zhang, H.; Zhou, Y.; Wang, Y. Expression of TpNRAMP5, a metal transporter from Polish wheat (Triticum polonicum L.), enhances the accumulation of Cd, Co and Mn in transgenic Arabidopsis plants. Planta 2018, 247, 1395–1406. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, X.; Yao, Q.; Long, D.; Fan, X.; Kang, H.; Zeng, J.; Sha, L.; Zhang, H.; Zhou, Y.; et al. Overexpression of TtNRAMP6 enhances the accumulation of Cd in Arabidopsis. Gene 2019, 696, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Feng Ma, J. The HvNramp5 Transporter Mediates Uptake of Cadmium and Manganese, But Not Iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokosho, K.; Yamaji, N.; Ma, J.F. Buckwheat FeNramp5 Mediates High Manganese Uptake in Roots. Plant Cell Physiol. 2021, 62, 600–609. [Google Scholar] [CrossRef]
- Meng, J.G.; Zhang, X.D.; Tan, S.K.; Zhao, K.X.; Yang, Z.M. Genome-wide identification of Cd-responsive NRAMP transporter genes and analyzing expression of NRAMP 1 mediated by miR167 in Brassica napus. Biometals 2017, 30, 917–931. [Google Scholar] [CrossRef]
- Yue, X.; Song, J.; Fang, B.; Wang, L.; Zou, J.; Su, N.; Cui, J. BcNRAMP1 promotes the absorption of cadmium and manganese in Arabidopsis. Chemosphere 2021, 283, 131113. [Google Scholar] [CrossRef]
- Milner, M.J.; Mitani-Ueno, N.; Yamaji, N.; Yokosho, K.; Craft, E.; Fei, Z.; Ebbs, S.; Clemencia Zambrano, M.; Ma, J.F.; Kochian, L.V. Root and shoot transcriptome analysis of two ecotypes of Noccaea caerulescens uncovers the role of NcNramp1 in Cd hyperaccumulation. Plant J. 2014, 78, 398–410. [Google Scholar] [CrossRef]
- Oomen, R.J.; Wu, J.; Lelievre, F.; Blanchet, S.; Richaud, P.; Barbier-Brygoo, H.; Aarts, M.G.; Thomine, S. Functional characterization of NRAMP3 and NRAMP4 from the metal hyperaccumulator Thlaspi caerulescens. New Phytol. 2009, 181, 637–650. [Google Scholar] [CrossRef]
- Wei, W.; Chai, T.; Zhang, Y.; Han, L.; Xu, J.; Guan, Z. The Thlaspi caerulescens NRAMP homologue TcNRAMP3 is capable of divalent cation transport. Mol. Biotechnol. 2009, 41, 15–21. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Song, H.; Zhao, J.; Shabala, S.; Tian, S.; Yang, X. A novel plasma membrane-based NRAMP transporter contributes to Cd and Zn hyperaccumulation in Sedum alfredii Hance. Environ. Exp. Bot. 2020, 176. [Google Scholar] [CrossRef]
- Feng, Y.; Wu, Y.; Zhang, J.; Meng, Q.; Wang, Q.; Ma, L.; Ma, X.; Yang, X. Ectopic expression of SaNRAMP3 from Sedum alfredii enhanced cadmium root-to-shoot transport in Brassica juncea. Ecotoxicol. Environ. Saf. 2018, 156, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Han, X.; Fang, J.; Lu, Z.; Qiu, W.; Liu, M.; Sang, J.; Jiang, J.; Zhuo, R. Sedum alfredii SaNramp6 Metal Transporter Contributes to Cadmium Accumulation in Transgenic Arabidopsis thaliana. Sci. Rep. 2017, 7, 13318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Chen, S.; Han, X.; Zhang, J.; Qiao, G.; Jiang, Y.; Zhuo, R.; Qiu, W. A Single Amino Acid Change in Nramp6 from Sedum Alfredii Hance Affects Cadmium Accumulation. Int. J. Mol. Sci. 2020, 21, 3169. [Google Scholar] [CrossRef] [PubMed]
- Zha, Q.; Xiao, Z.; Zhang, X.; Han, Z.; Wang, Y. Cloning and functional analysis of MxNRAMP1 and MxNRAMP3, two genes related to high metal tolerance of Malus xiaojinensis. S. Afr. J. Bot. 2016, 102, 75–80. [Google Scholar] [CrossRef]
- Zhang, W.; Yue, S.; Song, J.; Xun, M.; Han, M.; Yang, H. MhNRAMP1 from Malus hupehensis Exacerbates Cell Death by Accelerating Cd Uptake in Tobacco and Apple Calli. Front. Plant Sci. 2020, 11, 957. [Google Scholar] [CrossRef]
- Chen, Y.; Li, G.; Yang, J.; Zhao, X.; Sun, Z.; Hou, H. Role of Nramp transporter genes of Spirodela polyrhiza in cadmium accumulation. Ecotoxicol. Environ. Saf. 2021, 227, 112907. [Google Scholar] [CrossRef]
- Nakanishi-Masuno, T.; Shitan, N.; Sugiyama, A.; Takanashi, K.; Inaba, S.; Kaneko, S.; Yazaki, K. The Crotalaria juncea metal transporter CjNRAMP1 has a high Fe uptake activity, even in an environment with high Cd contamination. Int. J. Phytoremediat. 2018, 20, 1427–1437. [Google Scholar] [CrossRef]
- Liu, W.; Huo, C.; He, L.; Ji, X.; Yu, T.; Yuan, J.; Zhou, Z.; Song, L.; Yu, Q.; Chen, J.; et al. The NtNRAMP1 transporter is involved in cadmium and iron transport in tobacco (Nicotiana tabacum). Plant Physiol. Biochem. 2022, 173, 59–67. [Google Scholar] [CrossRef]
- Jia, H.; Yin, Z.; Xuan, D.; Lian, W.; Han, D.; Zhu, Z.; Li, C.; Li, C.; Song, Z. Mutation of NtNRAMP3 improves cadmium tolerance and its accumulation in tobacco leaves by regulating the subcellular distribution of cadmium. J. Hazard. Mater. 2022, 432, 128701. [Google Scholar] [CrossRef]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.E.; Jarvis, R.S.; Sherson, S.M.; Cobbett, C.S. Functional analysis of the heavy metal binding domains of the Zn/Cd-transporting ATPase, HMA2, in Arabidopsis thaliana. New Phytol. 2009, 181, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Gravot, A.; Lieutaud, A.; Verret, F.; Auroy, P.; Vavasseur, A.; Richaud, P. AtHMA3, a Plant P1B-ATPase, functions as a Cd/Pb transporter in yeast. FEBS Lett. 2004, 561, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/Co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verret, F.; Gravot, A.; Auroy, P.; Leonhardt, N.; David, P.; Nussaume, L.; Vavasseur, A.; Richaud, P. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and Plant metal tolerance. FEBS Lett. 2004, 576, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Ceasar, S.A.; Lekeux, G.; Motte, P.; Xiao, Z.; Galleni, M.; Hanikenne, M. di-Cysteine Residues of the Arabidopsis thaliana HMA4 C-Terminus Are Only Partially Required for Cadmium Transport. Front. Plant Sci. 2020, 11, 560. [Google Scholar] [CrossRef]
- Siemianowski, O.; Barabasz, A.; Kendziorek, M.; Ruszczynska, A.; Bulska, E.; Williams, L.E.; Antosiewicz, D.M. HMA4 expression in tobacco reduces Cd accumulation due to the induction of the apoplastic barrier. J. Exp. Bot. 2014, 65, 1125–1139. [Google Scholar] [CrossRef] [Green Version]
- Satoh-Nagasawa, N.; Mori, M.; Nakazawa, N.; Kawamoto, T.; Nagato, Y.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant Cell Physiol. 2012, 53, 213–224. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Feng Ma, J. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.Q.; Xuan, W.; Huang, X.Y.; Mao, C.; Zhao, F.J. Cadmium Inhibits Lateral Root Emergence in Rice by Disrupting OsPIN-Mediated Auxin Distribution and the Protective Effect of OsHMA3. Plant Cell Physiol. 2021, 62, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, L.; Tang, Z.; Huang, X.Y.; Ma, J.F.; Zhao, F.J. Producing cadmium-free Indica rice by overexpressing OsHMA3. Environ. Int. 2019, 126, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S.; Suzuki, T.; Tezuka, K.; Satoh-Nagasawa, N.; Takahashi, H.; Sakurai, K.; Watanabe, A.; Fujimura, T.; Akagi, H. Functional analysis of the C-terminal region of the vacuolar cadmium-transporting rice OsHMA3. FEBS Lett. 2014, 588, 789–794. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, Y.Y.; Lee, Y.; An, G. Rice P1B-type heavy-metal ATPase, OsHMA9, is a metal efflux protein. Plant Physiol. 2007, 145, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Wang, J.; Chai, T.; Zhang, Y.; Feng, S.; Li, Y.; Zhao, H.; Liu, H.; Chai, X. Functional analyses of TaHMA2, a P(1B)-type ATPase in wheat. Plant Biotechnol. J. 2013, 11, 420–431. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Liu, Y.; Yu, K.; Zhou, Y. GmHMA3 sequesters Cd to the root endoplasmic reticulum to limit translocation to the stems in soybean. Plant Sci. 2018, 270, 23–29. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, L.; Zhao, F.J.; Wu, L.; Liu, A.; Xu, W. SpHMA1 is a chloroplast cadmium exporter protecting photochemical reactions in the Cd hyperaccumulator Sedum plumbizincicola. Plant Cell Environ. 2019, 42, 1112–1124. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, M.; Shohag, M.J.; Tian, S.; Song, H.; Feng, Y.; Yang, X. Enhanced expression of SaHMA3 plays critical roles in Cd hyperaccumulation and hypertolerance in Cd hyperaccumulator Sedum alfredii Hance. Planta 2016, 243, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Milner, M.J.; Yamaji, N.; Yokosho, K.; Koyama, E.; Clemencia Zambrano, M.; Kaskie, M.; Ebbs, S.; Kochian, L.V.; Ma, J.F. Elevated expression of TcHMA3 plays a key role in the extreme Cd tolerance in a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Plant J. 2011, 66, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Papoyan, A.; Kochian, L.V. Identification of Thlaspi caerulescens genes that may be involved in heavy metal hyperaccumulation and tolerance. Characterization of a novel heavy metal transporting ATPase. Plant Physiol. 2004, 136, 3814–3823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Liang, S.; Xiang, W.; Dai, H.; Duan, Y.; Kang, F.; Chai, T. A repeat region from the Brassica juncea HMA4 gene BjHMA4R is specifically involved in Cd(2+) binding in the cytosol under low heavy metal concentrations. BMC Plant Biol. 2019, 19, 89. [Google Scholar] [CrossRef]
- Guo, Q.; Tian, X.; Mao, P.; Meng, L. Functional characterization of IlHMA2, a P1B2-ATPase in Iris lactea response to Cd. Environ. Exp. Bot. 2019, 157, 131–139. [Google Scholar] [CrossRef]
- Wang, X.; Zhi, J.; Liu, X.; Zhang, H.; Liu, H.; Xu, J. Transgenic tobacco plants expressing a P1B-ATPase gene from Populus tomentosa Carr. (PtoHMA5) demonstrate improved cadmium transport. Int. J. Biol. Macromol. 2018, 113, 655–661. [Google Scholar] [CrossRef]
- Grafe, K.; Schmitt, L. The ABC transporter G subfamily in Arabidopsis thaliana. J. Exp. Bot. 2021, 72, 92–106. [Google Scholar] [CrossRef]
- Verrier, P.J.; Bird, D.; Burla, B.; Dassa, E.; Forestier, C.; Geisler, M.; Klein, M.; Kolukisaoglu, U.; Lee, Y.; Martinoia, E.; et al. Plant ABC proteins–a unified nomenclature and updated inventory. Trends Plant Sci. 2008, 13, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Bovet, L.; Kushnir, S.; Noh, E.W.; Martinoia, E.; Lee, Y. AtATM3 is involved in heavy metal resistance in Arabidopsis. Plant Physiol. 2006, 140, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Rea, P.A. Plant ATP-binding cassette transporters. Annu. Rev. Plant Biol. 2007, 58, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, P.; Zanella, L.; De Paolis, A.; Di Litta, D.; Cecchetti, V.; Falasca, G.; Barbieri, M.; Altamura, M.M.; Costantino, P.; Cardarelli, M. Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin-mediated cadmium tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 3815–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaillard, S.; Jacquet, H.; Vavasseur, A.; Leonhardt, N.; Forestier, C. AtMRP6/AtABCC6, an ATP-binding cassette transporter gene expressed during early steps of seedling development and up-regulated by cadmium in Arabidopsis thaliana. BMC Plant Biol. 2008, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Wojas, S.; Hennig, J.; Plaza, S.; Geisler, M.; Siemianowski, O.; Sklodowska, A.; Ruszczynska, A.; Bulska, E.; Antosiewicz, D.M. Ectopic expression of Arabidopsis ABC transporter MRP7 modifies cadmium root-to-shoot transport and accumulation. Environ. Pollut. 2009, 157, 2781–2789. [Google Scholar] [CrossRef]
- Bhuiyan, M.S.U.; Min, S.R.; Jeong, W.J.; Sultana, S.; Choi, K.S.; Lee, Y.; Liu, J.R. Overexpression of AtATM3 in Brassica juncea confers enhanced heavy metal tolerance and accumulation. Plant Cell Tissue Organ. Cult. 2011, 107, 69–77. [Google Scholar] [CrossRef]
- Yang, G.; Fu, S.; Huang, J.; Li, L.; Long, Y.; Wei, Q.; Wang, Z.; Chen, Z.; Xia, J. The tonoplast-localized transporter OsABCC9 is involved in cadmium tolerance and accumulation in rice. Plant Sci. 2021, 307, 110894. [Google Scholar] [CrossRef]
- Fu, S.; Lu, Y.; Zhang, X.; Yang, G.; Chao, D.; Wang, Z.; Shi, M.; Chen, J.; Chao, D.Y.; Li, R.; et al. The ABC transporter ABCG36 is required for cadmium tolerance in rice. J. Exp. Bot. 2019, 70, 5909–5918. [Google Scholar] [CrossRef]
- Oda, K.; Otani, M.; Uraguchi, S.; Akihiro, T.; Fujiwara, T. Rice ABCG43 is Cd inducible and confers Cd tolerance on yeast. Biosci. Biotechnol. Biochem. 2011, 75, 1211–1213. [Google Scholar] [CrossRef]
- Cai, X.; Wang, M.; Jiang, Y.; Wang, C.; Ow, D.W. Overexpression of OsABCG48 Lowers Cadmium in Rice (Oryza sativa L.). Agronomy 2021, 11, 918. [Google Scholar] [CrossRef]
- Bhati, K.K.; Alok, A.; Kumar, A.; Kaur, J.; Tiwari, S.; Pandey, A.K. Silencing of ABCC13 transporter in wheat reveals its involvement in grain development, phytic acid accumulation and lateral root formation. J. Exp. Bot. 2016, 67, 4379–4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.H.; Wang, C.J.; Li, R.F.; Yi, Y.J.; Zeng, L.; Yang, H.; Zhang, C.F.; Song, K.Y.; Guo, S.J. Transcriptome-based identification and expression characterization of RgABCC transp.porters in Rehmannia glutinosa. PLoS ONE 2021, 16, e0253188. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Peng, Z.; Li, J.; Huang, W.; Liu, Y.; Wang, X.; Xie, S.; Sun, L.; Han, E.; et al. Ectopic Expression of Poplar ABC Transporter PtoABCG36 Confers Cd Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 3293. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Pence, N.S.; Larsen, P.B.; Ebbs, S.D.; Letham, D.L.D.; Lasat, M.M.; Garvin, D.F.; Eide, D.; Kochian, L.V. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc. Natl. Acad. Sci. USA 2000, 97, 4956–4960. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.F.; Hassan, Z.; Talukdar, S.; Schat, H.; Aarts, M.G. Expression of the ZNT1 Zinc Transporter from the Metal Hyperaccumulator Noccaea caerulescens Confers Enhanced Zinc and Cadmium Tolerance and Accumulation to Arabidopsis thaliana. PLoS ONE 2016, 11, e0149750. [Google Scholar] [CrossRef]
- Connolly, E.L.; Fett, J.P.; Guerinot, M.L. Expression of the IRT1 metal transporter is controlled by metals at the levels of transcript and protein accumulation. Plant Cell 2002, 14, 1347–1357. [Google Scholar] [CrossRef] [Green Version]
- Korshunova, Y.O.; Eide, D.; Clark, W.G.; Guerinot, M.L.; Pakrasi, H.B. The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant Mol. Biol. 1999, 40, 37–44. [Google Scholar] [CrossRef]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Qu, M.; Zhu, Y.; Peng, C.; Wang, J.; Gao, D.; Chen, C. ZINC TRANSPORTER5 and ZINC TRANSPORTER9 Function Synergistically in Zinc/Cadmium Uptake. Plant Physiol. 2020, 183, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, P.G.; Kuruvilla, S.; Mathew, M.K. Functional characterization of a transition metal ion transporter, OsZIP6 from rice (Oryza sativa L.). Plant Physiol. Biochem. 2015, 97, 165–174. [Google Scholar] [CrossRef]
- Tan, L.; Zhu, Y.; Fan, T.; Peng, C.; Wang, J.; Sun, L.; Chen, C. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochem. Biophys. Res. Commun. 2019, 512, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, Y.; Liu, Z.; Tian, J.; Liang, L.; Qiu, Y.; Wang, G.; Du, Q.; Cheng, D.; Cai, H.; et al. A high activity zinc transporter OsZIP9 mediates zinc uptake in rice. Plant J. 2020, 103, 1695–1709. [Google Scholar] [CrossRef]
- Barabasz, A.; Palusinska, M.; Papierniak, A.; Kendziorek, M.; Kozak, K.; Williams, L.E.; Antosiewicz, D.M. Functional Analysis of NtZIP4B and Zn Status-Dependent Expression Pattern of Tobacco ZIP Genes. Front. Plant Sci. 2018, 9, 1984. [Google Scholar] [CrossRef] [Green Version]
- Maslinska-Gromadka, K.; Barabasz, A.; Palusinska, M.; Kozak, K.; Antosiewicz, D.M. Suppression of NtZIP4A/B Changes Zn and Cd Root-to-Shoot Translocation in a Zn/Cd Status-Dependent Manner. Int. J. Mol. Sci. 2021, 22, 5355. [Google Scholar] [CrossRef]
- Fan, W.; Liu, C.; Cao, B.; Qin, M.; Long, D.; Xiang, Z.; Zhao, A. Genome-Wide Identification and Characterization of Four Gene Families Putatively Involved in Cadmium Uptake, Translocation and Sequestration in Mulberry. Front. Plant Sci. 2018, 9, 879. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, C.; Guo, H.; Hu, Y.; He, Y.; Jiang, D. Overexpression of a Miscanthus sacchariflorus yellow stripe-like transporter MsYSL1 enhances resistance of Arabidopsis to cadmium by mediating metal ion reallocation. Plant Growth Regul. 2018, 85, 101–111. [Google Scholar] [CrossRef]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2016, 36, 281–296. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Y.; Deng, Y.; Liu, Y.; Dong, M.; Tian, Y.; Sun, H.; Li, Y. Cloning and functional analysis of the VcCXIP4 and VcYSL6 genes as Cd-regulating genes in blueberry. Gene 2019, 686, 104–117. [Google Scholar] [CrossRef]
- Wang, J.W.; Li, Y.; Zhang, Y.X.; Chai, T.Y. Molecular cloning and characterization of a Brassica juncea yellow stripe-like gene, BjYSL7, whose overexpression increases heavy metal tolerance of tobacco. Plant Cell Rep. 2013, 32, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.K.; Garvin, D.F.; Kochian, L.V. Kinetic properties of a micronutrient transporter from Pisum sativum indicate a primary function in Fe uptake from the soil. Planta 2004, 218, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, X.; Chai, S.; Sheng, H.; Sha, L.; Fan, X.; Zeng, J.; Kang, H.; Zhang, H.; Xiao, X.; et al. TpIRT1 from Polish wheat (Triticum polonicum L.) enhances the accumulation of Fe, Mn, Co, and Cd in Arabidopsis. Plant Sci. 2021, 312, 111058. [Google Scholar] [CrossRef]
- Lee, S.; An, G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009, 32, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Pedas, P.; Ytting, C.K.; Fuglsang, A.T.; Jahn, T.P.; Schjoerring, J.K.; Husted, S. Manganese efficiency in barley: Identification and characterization of the metal ion transporter HvIRT1. Plant Physiol. 2008, 148, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Zang, J.; Huo, Y.; Liu, J.; Zhang, H.; Liu, J.; Chen, H. Maize YSL2 is required for iron distribution and development in kernels. J. Exp. Bot. 2020, 71, 5896–5910. [Google Scholar] [CrossRef]

| Plant Species | Genes | Expression Sites | Subcellular Location | Function | References |
|---|---|---|---|---|---|
| Arabidopsis thaliana | AtNramp1 | Roots | Plasma membrane | - | [25] |
| AtNramp3 | Roots and aerial parts | Tonoplast | Cd transport | [25,26,27,28] | |
| AtNramp4 | Roots and aerial parts | Tonoplast | Cd transport | [25,28,29] | |
| AtNramp6 | Seed embryo, lateral roots and young leaves | Golgi/trans-Golgi network | Cd transport | [30] | |
| Oryza sativa L. | OsNramp1 | Roots and leaves | Plasma membrane | Cd uptake and translocation | [31,32,33] |
| OsNramp2 | Embryo of germinating seeds, roots, leaf sheaths and leaf blades | Tonoplast | Cd retranslocation | [34,35] | |
| OsNramp5 | Roots epidermis, exodermis, outer layers of cortex and tissues around xylem | Plasma membrane | Cd uptake | [36,37,38,39] | |
| Triticum polonicum L. | TpNramp3 | leaf blades and roots at the jointing and booting stages, first nodes at the grain filling stage | Plasma membrane | Cd accumulation | [40] |
| TpNramp5 | Roots and basal stems of DPW | Plasma membrane | Cd accumulation | [41] | |
| Triticum turgidum L. | TtNramp6 | Roots | Plasma membrane | Cd accumulation | [42] |
| Hordeum vulgare | HvNramp5 | Roots | Plasma membrane | Cd uptake | [43] |
| Fagopyrum esculentum Moench | FeNramp5 | Roots | Plasma membrane | Cd uptake | [44] |
| Brassica napus | BnNramp1b | Vegetative tissue, flowers and siliques | - | - | [45] |
| Brassica rapa L. | BcNramp1 | Roots | Plasma membrane | Cd uptake | [46] |
| Noccaea caerulescens (Thlaspi caeulescens) | NcNramp1 | Roots and shoots | Plasma membrane | - | [47] |
| TcNramp3 | - | Tonoplast | - | [48,49] | |
| TcNramp4 | - | Tonoplast | - | [48] | |
| Sedum alfredii Hance | SaNramp1 | Young tissues of the shoots | Plasma membrane | Cd translocation | [50] |
| SaNramp3 | - | - | Cd translocation | [51] | |
| SaNramp6 | Roots | Plasma membrane | Cd uptake or translocation | [52,53] | |
| Malus xiaojinensis | MxNramp1 | Roots | Plasma membrane | Cd uptake and translocation | [54] |
| MxNramp3 | Roots and leaves | Tonoplast | Cd uptake and translocation | [54] | |
| Malus hupehensis | MhNramp1 | Roots | Cell membrane | Cd uptake | [55] |
| Spirodela polyrhiza | SpNramp1 | - | Plasma membrane | Cd accumulation | [56] |
| SpNramp2 | - | Plasma membrane | Cd accumulation | [56] | |
| SpNramp3 | - | Plasma membrane | - | [56] | |
| Crotalaria juncea | CjNramp1 | Leaves, stems, and roots | Plasma membrane | Cd uptake and translocation | [57] |
| Nicotiana tabacum | NtNRAMP1 | Roots | - | Cd uptake | [58] |
| NtNRAMP3 | Conductive tissue of leaves | Tonoplast | Cd efflux | [59] |
| Plant Species | Genes | Expression Sites | Subcellular Location | Function | References |
|---|---|---|---|---|---|
| Arabidopsis thaliana | AtABCC1 | - | Tonoplast | Cd sequestration | [92] |
| AtABCC2 | - | Tonoplast | Cd sequestration | [92] | |
| AtABCC3 | - | - | Cd sequestration | [94] | |
| AtMRP6/AtABCC6 | Xylem-opposite pericycle cells where lateral roots initiate | - | [95] | ||
| AtMRP7 | - | Plasma membrane and tonoplast | Cd sequestration | [96] | |
| AtPDR8 | Root epidermal cells | Plasma membrane | Cd efflux | [91] | |
| AtATM3 | Roots | Mitochondrial membrane | - | [90,97] | |
| Oryza sativa L. | OsABCC9 | Root stele | Tonoplast | Cd sequestration | [98] |
| OsABCG36 | Roots | Plasma membrane | Cd efflux | [99] | |
| OsABCG43 | Roots | - | Cd sequestration | [100] | |
| OsABCG48 | - | - | - | [101] | |
| Triticum aestivum | TaABCC13 | - | - | Cd uptake and transport | [102] |
| Rehmannia glutinosa | RgABCC1 | Roots | - | - | [103] |
| Populus tomentosa | PtoABCG36 | Roots | Plasma membrane | Cd efflux | [104] |
| Plant Species | Genes | Expression Sites | Subcellular Location | Function | References |
|---|---|---|---|---|---|
| Genes encoding Zinc and Iron regulated transporter Protein (ZIP) | |||||
| Noccaea caerulescens L. | NcZNT1 | roots and shoots | - | - | [106,107] |
| Arabidopsis thaliana | AtIRT1 | Roots | Plasma membrane | Cd uptake | [108,109] |
| Oryza sativa L. | OsIRT1 | Roots | Plasma membrane | Cd uptake | [108,109,110] |
| OsIRT2 | Roots | Plasma membrane | Cd uptake | [110] | |
| OsZIP1 | Roots | Endoplasmic reticulum (ER) and plasma membrane | Cd efflux | [111] | |
| OsZIP5 | Roots | Plasma membrane | Cd uptake | [112] | |
| OsZIP6 | Shoots and roots | - | Cd uptake | [113] | |
| OsZIP7 | parenchyma cells of vascular bundles in roots and nodes | Plasma membrane | Cd translocation | [114] | |
| OsZIP9 | Roots | Plasma membrane | Cd uptake | [112,115] | |
| Nicotiana tabacum var Xanthi | NtZIP4A/B | Leaves and roots | Plasma membrane | Cd translocation | [116,117] |
| Morus alba | MaZIP4 | - | Plasma membrane | - | [118] |
| Genes encoding Yellow Stripe-Like proteins (YSL) | |||||
| Miscanthus sacchariflorus | MsYSL1 | Stems | Plasma membrane | Cd translocation | [119] |
| Solanum nigrum | SnYSL3 | Vascular tissues and epidermal cells of the roots and stems | Plasma membrane | Cd translocation | [120] |
| Vaccinium ssp. | VcYSL6 | - | Chloroplast | - | [121] |
| Brassica juncea | BjYSL7 | Stems | Plasma membrane | Cd translocation | [122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.; Lu, L. Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants. Toxics 2022, 10, 411. https://doi.org/10.3390/toxics10080411
Tao J, Lu L. Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants. Toxics. 2022; 10(8):411. https://doi.org/10.3390/toxics10080411
Chicago/Turabian StyleTao, Jingyu, and Lingli Lu. 2022. "Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants" Toxics 10, no. 8: 411. https://doi.org/10.3390/toxics10080411
APA StyleTao, J., & Lu, L. (2022). Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants. Toxics, 10(8), 411. https://doi.org/10.3390/toxics10080411