
Prenatal Lipopolysaccharide Exposure Alters Hepatic Drug-Metabolizing Enzyme Expression in Mouse Offspring via Histone Modifications

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
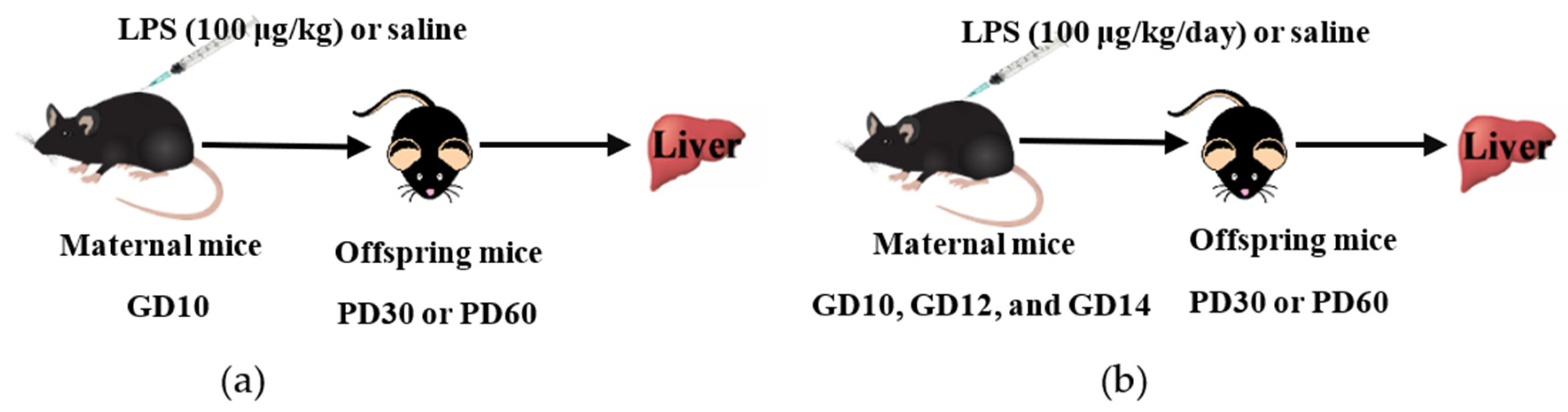
2.2. Animals and Procedures
2.3. Total RNA Isolation
2.4. Quantitative Real-Time Polymerase Chain Reaction
2.5. Western Blot Analysis
2.6. Chromatin Immunoprecipitation Analysis
2.7. Statistical Analysis
3. Results
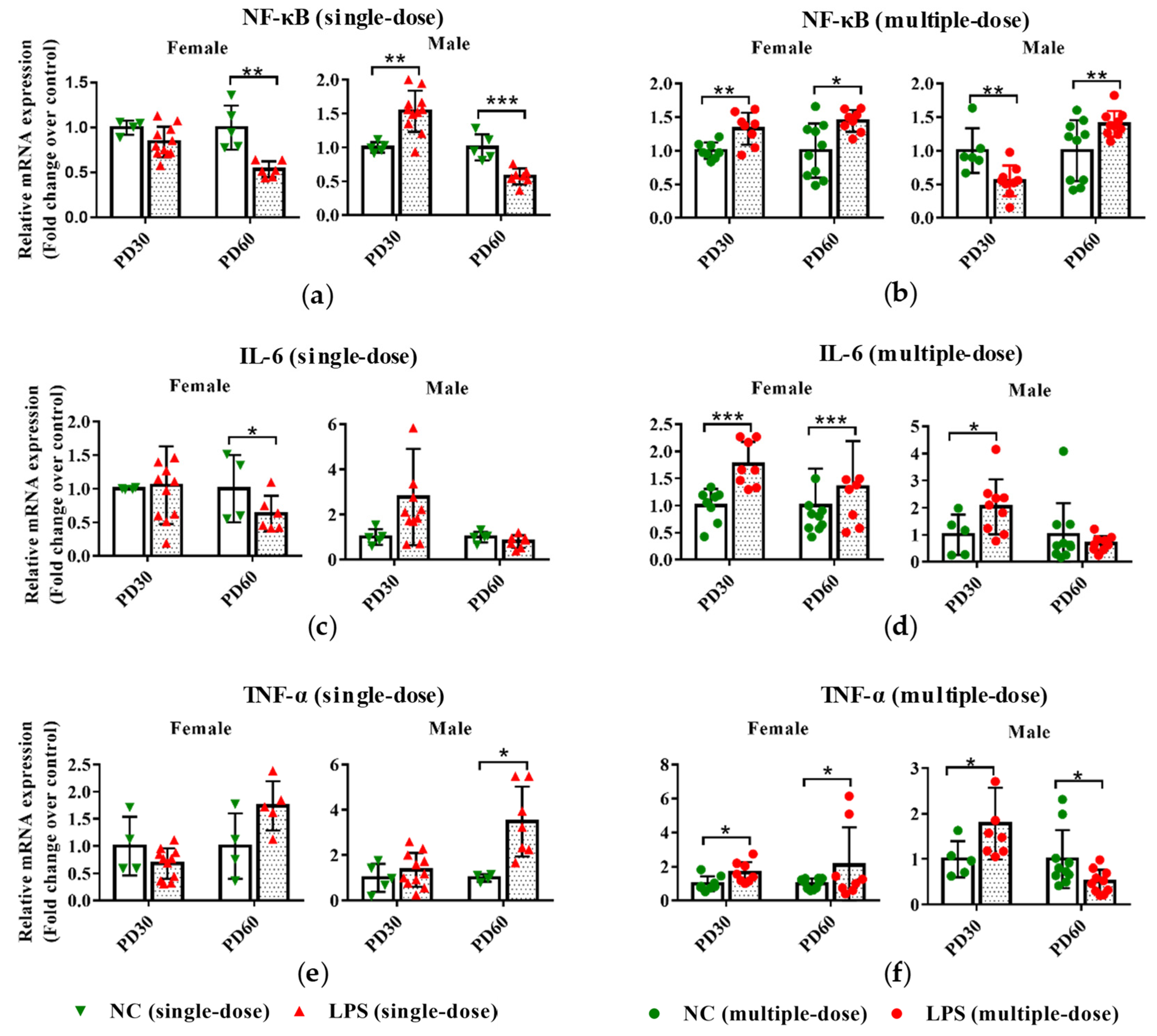
3.1. Prenatal LPS Exposure Alters the Hepatic Expression of Inflammatory-Related Genes in Offspring Mice at Different Ages
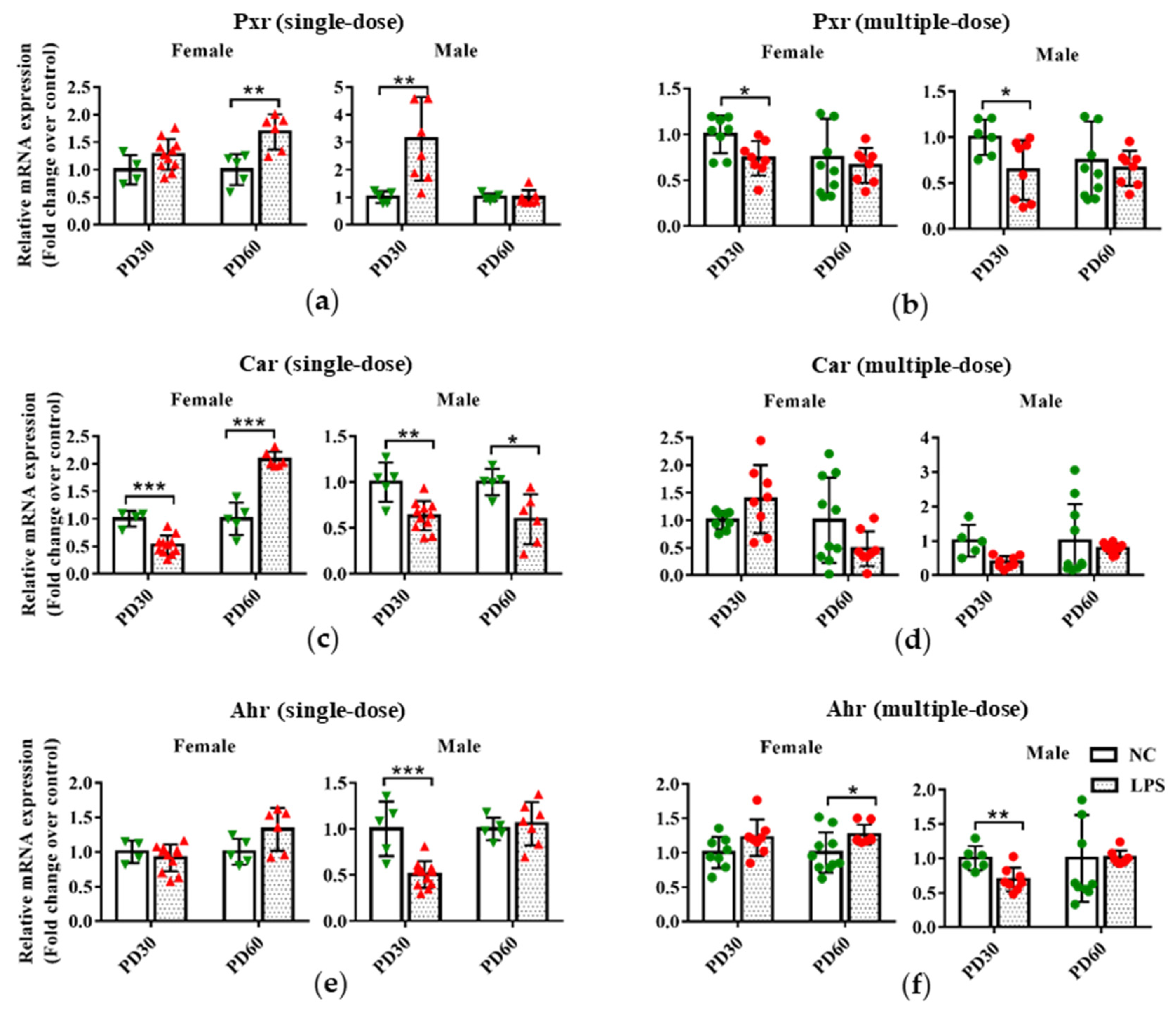
3.2. Prenatal LPS Exposure Affects the Hepatic Expression of Transcription Factors in Offspring Mice at Different Ages
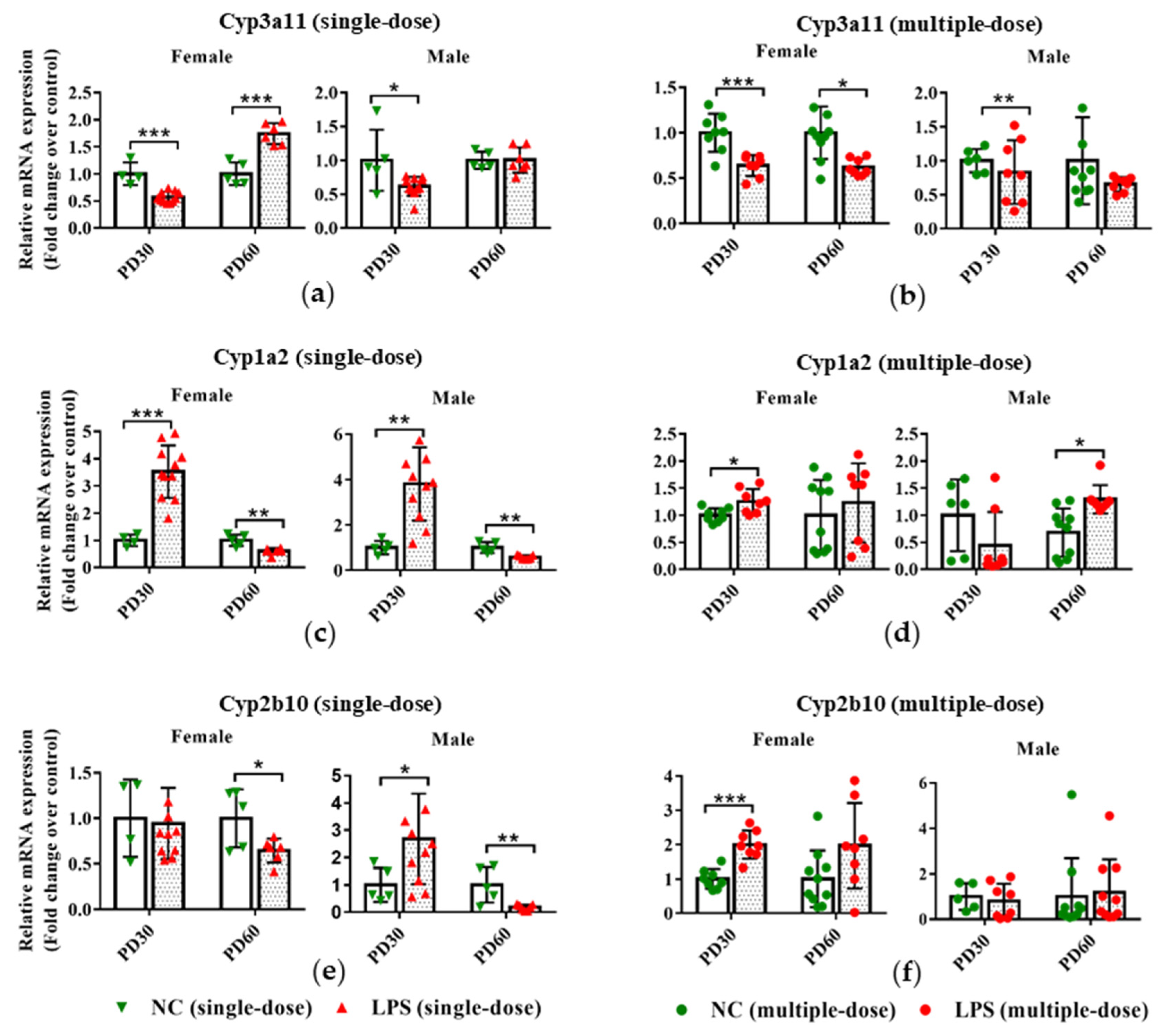
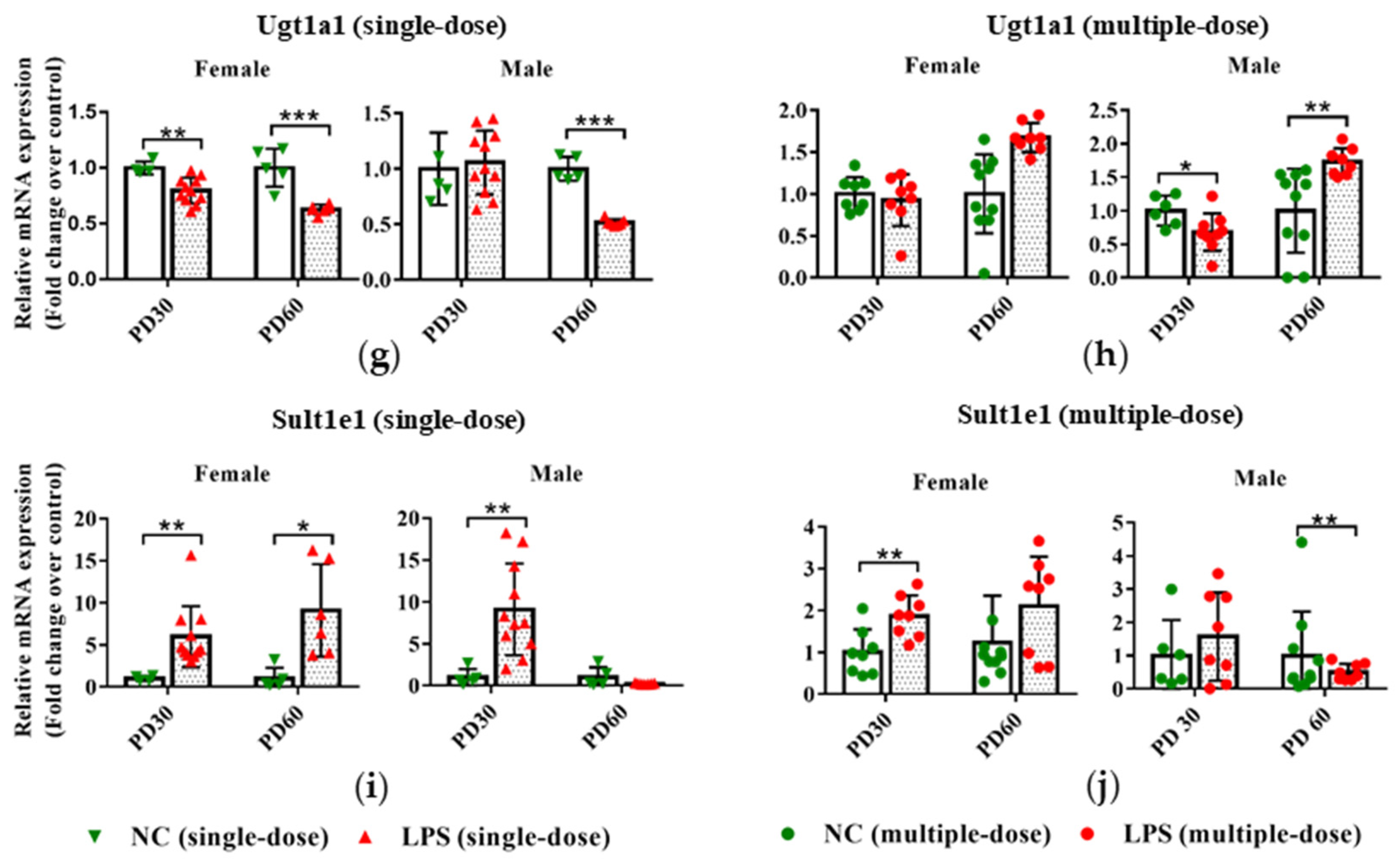
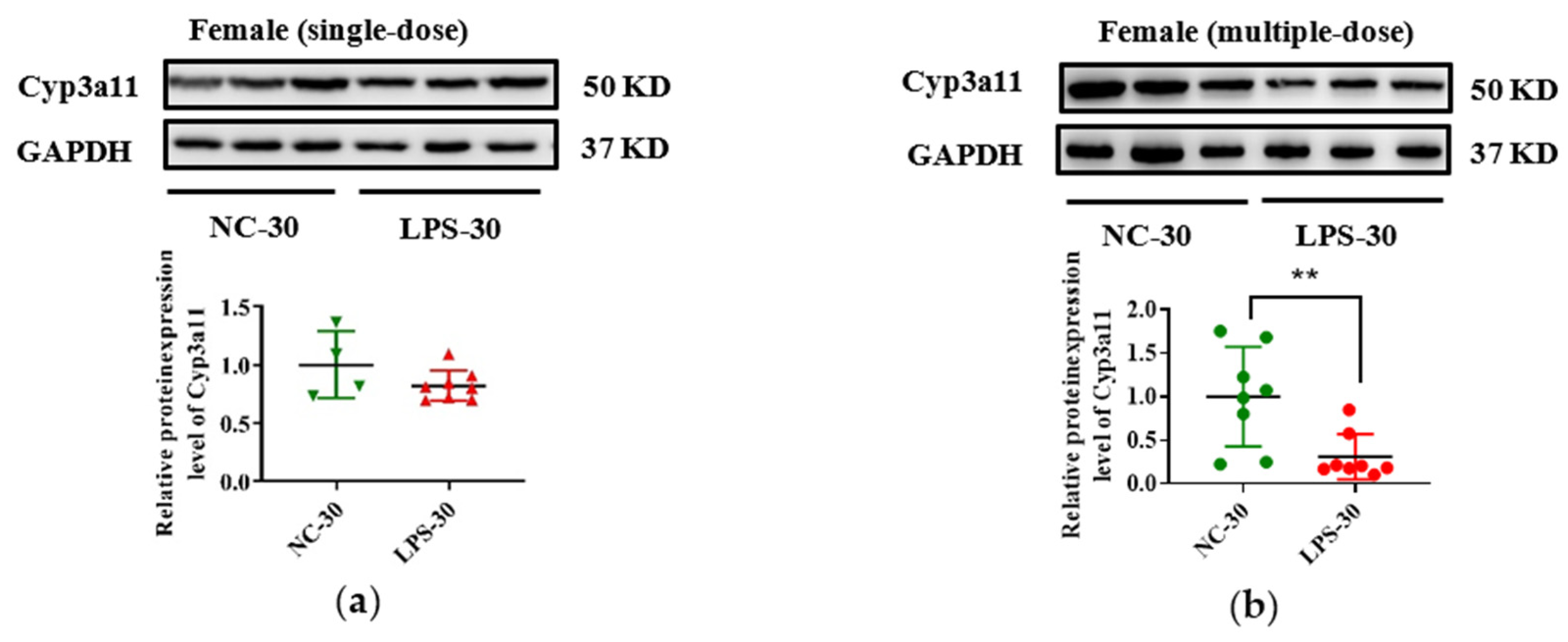
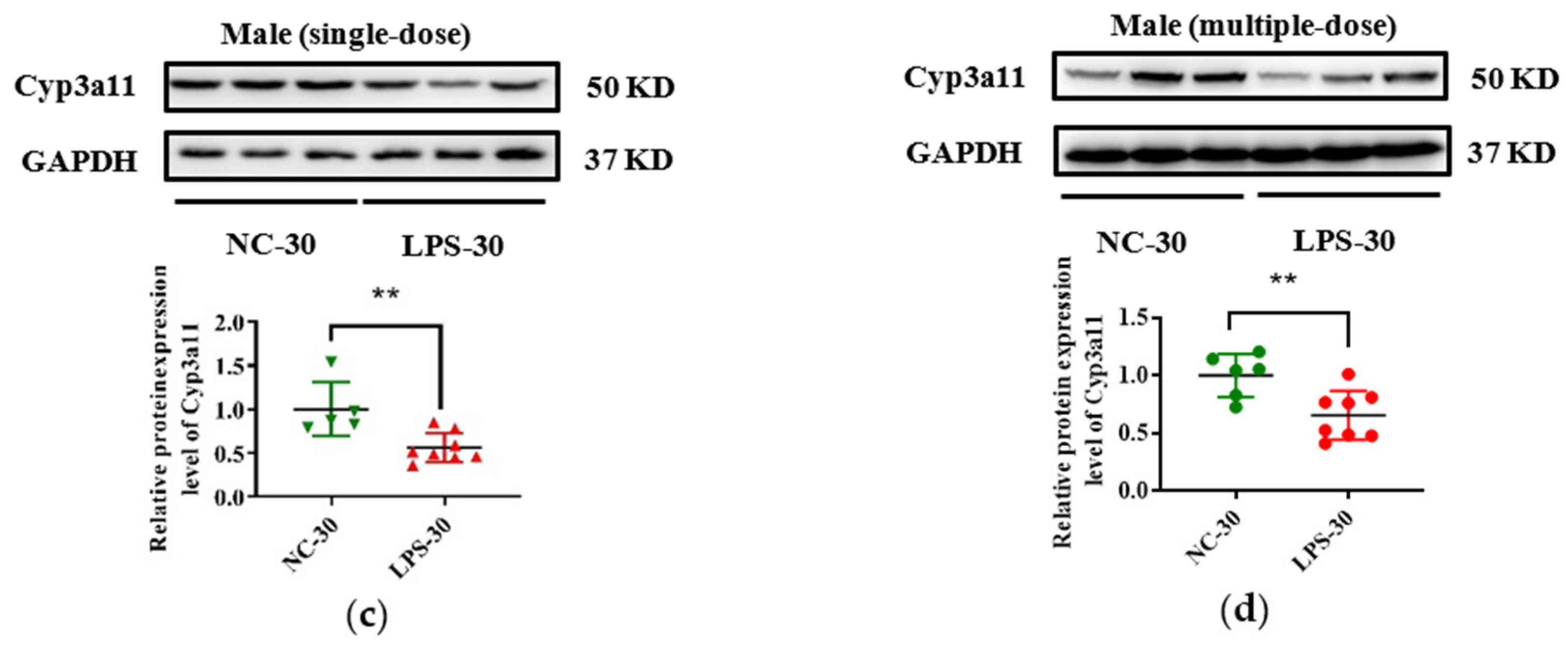
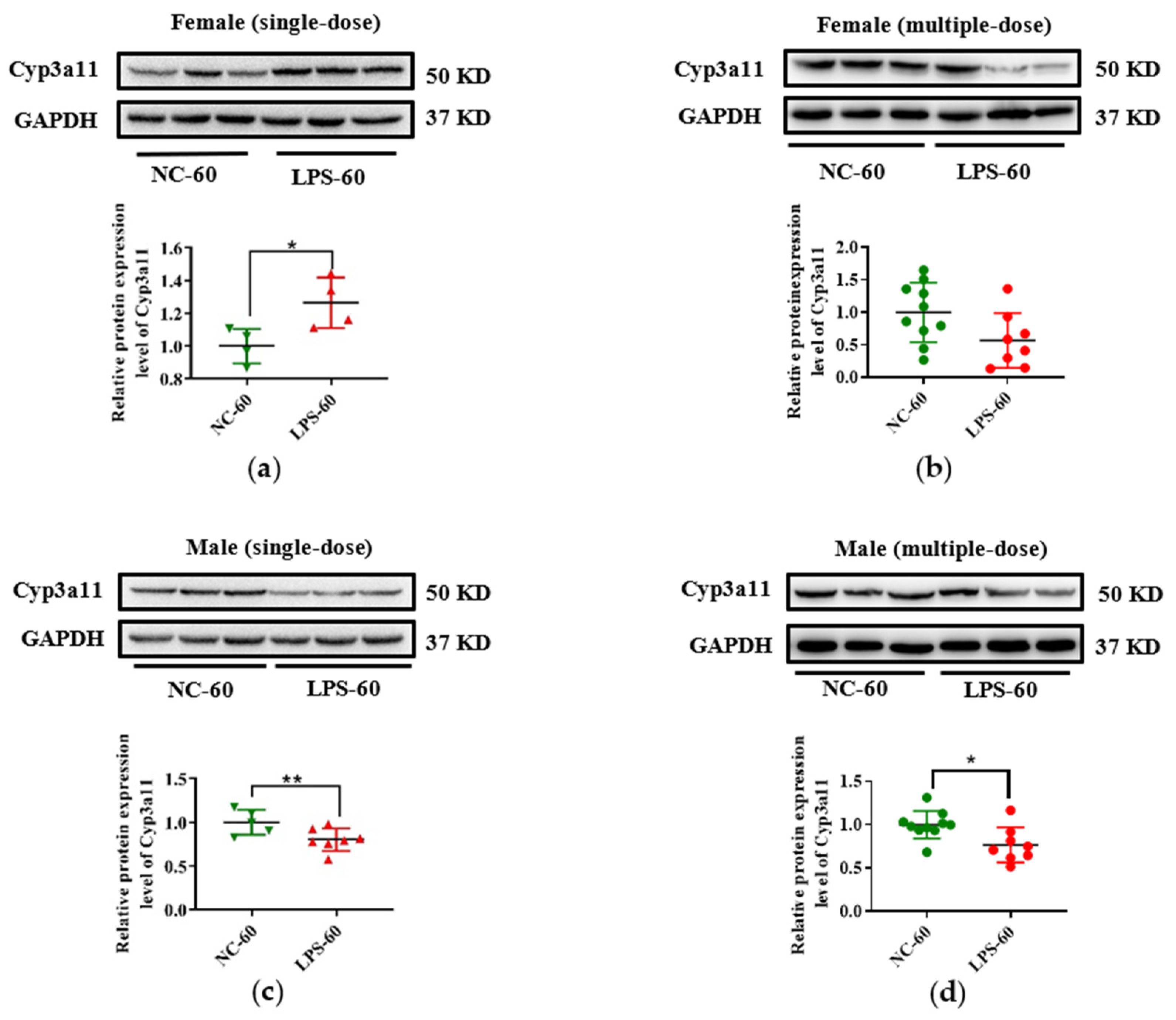
3.3. Prenatal LPS Exposure Impacts the Hepatic Expression of DMEs in Offspring Mice at Different Ages
3.4. Histone Modifications Contribute to the Decreased Cyp3a11 Expression Following Prenatal Exposure to Multiple Doses of LPS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Stavropoulou, E.; Pircalabioru, G.G.; Bezirtzoglou, E. The role of cytochromes P450 in infection. Front. Immunol. 2018, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, D.B.; Andersen, N.E.; Dunvald, A.C.D.; Pottegard, A.; Stage, T.B. Drug metabolism and drug transport of the 100 most prescribed oral drugs. Basic Clin. Pharmacol. Toxicol. 2022, 131, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Wu, J.J.; Wu, Y.; Li, H.X.; Finel, M.S.; Wang, D.D.; Ge, G.B. Optical substrates for drug-metabolizing enzymes: Recent advances and future perspectives. Acta Pharm. Sin. B 2022, 12, 1068–1099. [Google Scholar] [CrossRef]
- Tracy, T.S.; Chaudhry, A.S.; Prasad, B.; Thummel, K.E.; Schuetz, E.G.; Zhong, X.B.; Tien, Y.C.; Jeong, H.; Pan, X.; Shireman, L.M.; et al. Interindividual variability in cytochrome P450-mediated drug metabolism. Drug Metab. Dispos. 2016, 44, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Tien, Y.C.; Liu, K.; Pope, C.; Wang, P.; Ma, X.; Zhong, X.B. Dose of phenobarbital and age of treatment at early life are two key factors for the persistent induction of Cytochrome P450 enzymes in adult mouse liver. Drug. Metab. Dispos. 2015, 43, 1938–1945. [Google Scholar] [CrossRef] [Green Version]
- Dempsey, J.L.; Cui, J.Y. Regulation of hepatic long noncoding RNAs by pregnane X receptor and constitutive androstane receptor agonists in mouse liver. Drug Metab. Dispos. 2019, 47, 329–339. [Google Scholar]
- Dutta, M.; Lim, J.J.; Cui, J.Y. Pregnane X receptor and the gut-liver axis: A recent update. Drug Metab. Dispos. 2022, 50, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.N.; Greene, J.; Kenna, L.; Kissell, L.; Kuhn, M. The impact of infection and inflammation on drug metabolism, active transport, and systemic drug concentrations in veterinary species. Drug Metab. Dispos. 2020, 48, 631–644. [Google Scholar] [CrossRef]
- Lenoir, C.; Niederer, A.; Rollason, V.; Desmeules, J.A.; Daali, Y.; Samer, C.F. Prediction of cytochromes P450 3A and 2C19 modulation by both inflammation and drug interactions using physiologically based pharmacokinetics. CPT Pharmacomet. Syst. Pharm. 2022, 11, 30–43. [Google Scholar] [CrossRef]
- Wang, X.; Rao, J.; Tan, Z.; Xun, T.; Zhao, J.; Yang, X. Inflammatory signaling on cytochrome P450-mediated drug metabolism in hepatocytes. Front. Pharm. 2022, 13, 1043836. [Google Scholar] [CrossRef] [PubMed]
- White, C.M. Inflammation suppresses patients’ ability to metabolize cytochrome P450 substrate drugs. Ann. Pharm. 2022, 56, 809–819. [Google Scholar] [CrossRef]
- Lv, C.; Huang, L. Xenobiotic receptors in mediating the effect of sepsis on drug metabolism. Acta. Pharm Sin B 2020, 10, 33–41. [Google Scholar] [CrossRef]
- Hayden, M.S.; West, A.P.; Ghosh, S. NF-kappaB and the immune response. Oncogene 2006, 25, 6758–6780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Tabb, M.M.; Nelson, E.L.; Grün, F.; Verma, S.; Sadatrafiei , A.; Lin, M.; Mallick, S.; Forman, B.M.; Thummel, K.E.; et al. Mutual repression between steroid and xenobiotic receptor and NF-kappaB signaling pathways links xenobiotic metabolism and inflammation. J. Clin. Investig. 2006, 116, 2280–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Ke, S.; Liu, D. Role of NF-kappaB in regulation of PXR-mediated gene expression: A mechanism for the suppression of cytochrome P-450 3A4 by proinflammatory agents. J. Biol. Chem. 2006, 281, 17882–17889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barouki, R.; Melen, E.; Herceg, Z.; Beckers, J.; Chen, J.; Karagas, M.; Puga, A.; Xia, Y.; Chadwick, L.; Yan, W.; et al. Epigenetics as a mechanism linking developmental exposures to long-term toxicity. Environ. Int. 2018, 114, 77–86. [Google Scholar] [CrossRef]
- Kim, E.; Paik, D.; Ramirez, R.N.; Biggs, D.G.; Park, Y.; Kwon, H.K.; Choi, G.B.; Huh, J.R. Maternal gut bacteria drive intestinal inflammation in offspring with neurodevelopmental disorders by altering the chromatin landscape of CD4(+) T cells. Immunity 2022, 55, 145–158.e147. [Google Scholar] [CrossRef]
- Lim, A.I.; McFadden, T.; Link, V.M.; Han, S.J.; Karlsson, R.M.; Stacy, A.; Farley, T.K.; Lima-Junior, D.S.; Harrison, O.J.; Desai, J.V.; et al. Prenatal maternal infection promotes tissue-specific immunity and inflammation in offspring. Science 2021, 373, eabf3002. [Google Scholar] [CrossRef]
- Cao, N.; Lan, C.; Chen, C.; Xu, Z.; Luo, H.; Zheng, S.; Gong, X.; Ren, H.; Li, Z.; Qu, S.; et al. Prenatal Lipopolysaccharides Exposure Induces Transgenerational Inheritance of Hypertension. Circulation 2022, 146, 1082–1095. [Google Scholar] [CrossRef]
- Wang, P.; Liu, G.M.; Nie, Y.L.; Han, S.N.; Li, J.F.; Zhong, X.B.; Zhang, L.R. Epigenetic memory Is involved in the persistent Alterations of drug-processing genes in adult Mice due to PCN-activated PXR during early life. Toxicol. Sci. 2019, 172, 98–108. [Google Scholar] [CrossRef]
- Shin, A.; Waxman, D.J. Impact of neonatal activation of nuclear receptor CAR (Nr1i3) on Cyp2 gene expression in adult mouse liver. Toxicol. Sci. 2022, 187, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Chen, X.; Gao, M.; Zhou, L.; Li, X. Prenatal exposure to lipopolysaccharide induces PTX3 expression and results in obesity in mouse offspring. Inflammation 2017, 40, 1847–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Saadaoui, M.; Al Khodor, S. Infections and Pregnancy: effects on maternal and child health. Front. Cell. Infect. Microbiol. 2022, 12, 873253. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Tsujimoto, K.; Kawahori, K.; Hanzawa, N.; Hamaguchi, M.; Seki, T.; Nawa, M.; Ehara, T.; Kitamura, Y.; Hashimoto, K.; et al. Epigenetic modulation of Fgf21 in the perinatal mouse liver ameliorates diet-induced obesity in adulthood. Nat. Commun. 2018, 9, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Wang, J.; Xu, S.; Wang, L.; Song, M.; An, C.; Wang, X. Prenatal earthquake stress exposure in different gestational trimesters is associated with methylation changes in the glucocorticoid receptor gene (NR3C1) and long-term working memory in adulthood. Transl. Psychiatry 2022, 12, 176. [Google Scholar] [CrossRef]
- He, H.; Nie, Y.L.; Li, J.F.; Meng, X.G.; Yang, W.H.; Chen, Y.L.; Wang, S.J.; Ma, X.C.; Kan, Q.C.; Zhang, L.R.; et al. Developmental regulation of CYP3A4 and CYP3A7 in Chinese Han population. Drug Metab. Pharm. 2016, 31, 433–444. [Google Scholar] [CrossRef]
- Li, Y.; Cui, Y.; Hart, S.N.; Zhong, X.B. Dynamic patterns of histone methylation are associated with ontogenic expression of the Cyp3a genes during mouse liver maturation. Mol. Pharm. 2009, 75, 1171–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.L.; Meng, X.G.; Yan, L.; Wang, P.; Bi, H.Z.; Kan, Q.C.; Zhang, L.R. Histone modifications regulate the developmental expression of human hepatic UDP-glucuronosyltransferase 1A1. Drug Metab. Dispos. 2017, 45, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Wang, Y.T.; Liu, J.Y.; Nie, Y.L.; Zhong, X.B.; Kan, Q.; Zhang, L. Alterations of histone modifications contribute to pregnane X receptor-mediated induction of CYP3A4 by rifampicin. Mol. Pharm. 2017, 92, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Chen, W.D.; Fu, X.; Dong, B.; Wang, Y.D.; Shiah, S.; Moore, D.D.; Huang, W. Neonatal activation of the nuclear receptor CAR results in epigenetic memory and permanent change of drug metabolism in mouse liver. Hepatology 2012, 56, 1499–1509. [Google Scholar] [CrossRef]
- Hildreth, J.R.; Vickers, M.H.; Buklijas, T.; Bay, J.L. Understanding the importance of the early-life period for adult health: A systematic review. J. Dev. Orig. Health Dis. 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lodge-Tulloch, N.A.; Toews, A.J.; Atallah, A.; Cotechini, T.; Girard, S.; Graham, C.H. Cross-generational impact of innate immune memory following pregnancy complications. Cells 2022, 11, 3935. [Google Scholar] [CrossRef]
- Deng, Y.; Song, L.; Nie, X.; Shou, W.; Li, X. Prenatal inflammation exposure-programmed cardiovascular diseases and potential prevention. Pharm. Ther. 2018, 190, 159–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, D.; Liu, Y.; Chen, X.; Liu, J.; Liu, J.; Lai, W.; Li, S.; Wang, W.; Zhang, W.; Xiao, D.; et al. Activation of iNKT cells at the maternal-fetal interface predisposes offspring to cardiac injury. Circulation 2022, 145, 1032–1035. [Google Scholar] [CrossRef] [PubMed]
- van Groen, B.D.; Nicolai, J.; Kuik, A.C.; Van Cruchten, S.; van Peer, E.; Smits, A.; Schmidt, S.; de Wildt, S.N.; Allegaert, K.; De Schaepdrijver, L.; et al. Ontogeny of hepatic transporters and drug-metabolizing enzymes in humans and in nonclinical species. Pharmacol. Rev. 2021, 73, 597–567. [Google Scholar] [CrossRef]
- Peng, L.; Yoo, B.; Gunewardena, S.S.; Lu, H.; Klaassen, C.D.; Zhong, X.B. RNA sequencing reveals dynamic changes of mRNA abundance of cytochromes P450 and their alternative transcripts during mouse liver development. Drug Metab. Dispos. 2012, 40, 1198–1209. [Google Scholar] [CrossRef]
- Guan, X.; Dan, G.R.; Yang, Y.; Ji, Y.; Lai, W.J.; Wang, F.J.; Meng, M.; Mo, B.H.; Huang, P.; You, T.T.; et al. Prenatal inflammation exposure-programmed hypertension exhibits multi-generational inheritance via disrupting DNA methylome. Acta Pharm. Sin. 2022, 43, 1419–1429. [Google Scholar] [CrossRef]
- Xu, D.X.; Chen, Y.H.; Wang, J.P. Perinatal lipopolysaccharide exposure downregulates pregnane X receptor and Cyp3a11 expression in fetal mouse liver. Toxicol. Sci. 2005, 87, 38–45. [Google Scholar] [CrossRef]
- Chen, Y.H.; Wang, J.P.; Wang, H.; Sun, M.F.; Wei, L.Z.; Wei, W.; Xu, D.X. Lipopolysaccharide treatment downregulates the expression of the pregnane X receptor, cyp3a11 and mdr1a genes in mouse placenta. Toxicology 2005, 211, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Zhang, C.; Wang, S.F.; Ji, Y.L.; Wang, H.; Zhao, L.; Xu, D.X. Maternally administered lipopolysaccharide (LPS) upregulates the expression of heme oxygenase-1 in fetal liver: The role of reactive oxygen species. Toxicol. Lett. 2008, 176, 169–177. [Google Scholar] [CrossRef]
- Sachdeva, K.; Yan, B.; Chichester, C.O. Lipopolysaccharide and cecal ligation/puncture differentially affect the subcellular distribution of the pregnane X receptor but consistently cause suppression of its target genes CYP3A. Shock 2003, 19, 469–474. [Google Scholar] [CrossRef]
- Xu, D.X.; Wang, H.; Ning, H.; Zhao, L.; Chen, Y.H. Maternally administered melatonin differentially regulates lipopolysaccharide-induced proinflammatory and anti-inflammatory related genes in maternal serum, amniotic fluid, fetal liver, and fetal brain. J Pineal. Res. 2007, 43, 74–79. [Google Scholar] [CrossRef]
- Xiang, K.H.; Shen, P.; Gao, Z.Y.; Liu, Z.Y.; Hu, X.Y.; Liu, B.; Fu, Y.H. Formononetin Protects LPS-Induced Mastitis Through Suppressing Inflammation and Enhancing Blood-Milk Barrier Integrity via AhR-Induced Src Inactivation. Front. Immunol. 2022, 13, 814319. [Google Scholar] [CrossRef]
- Zheng, Z.P.; Wu, L.; Han, Y.Q.; Chen, J.; Zhu, S.; Yao, Y.Y.; Wang, B.H.; Li, L.J. Gut microbiota-controlled tryptophan metabolism improves D-Gal/LPS-induced acute liver failure in C57BL/6 mice. Engineering 2022, 14, 134–146. [Google Scholar] [CrossRef]
- Renaud, H.J.; Cui, J.Y.; Khan, M.; Klaassen, C.D. Tissue distribution and gender-divergent expression of 78 cytochrome P450 mRNAs in mice. Toxicol. Sci. 2011, 124, 261–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerges, S.H.; El-Kadi, A.O.S. Sexual dimorphism in the expression of cytochrome P450 enzymes in rat heart, liver, kidney, lung, brain, and small intestine. Drug Metab. Dispos. 2023, 51, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Meng, P.; Cheng, S.; Jiang, X.; Zhang, J.; Qin, X.; Tang, Q.; Bai, L.; Zou, Z.; Chen, C. Pregnancy exposure to carbon black nanoparticles induced neurobehavioral deficits that are associated with altered m(6)A modification in offspring. Neurotoxicology 2020, 81, 40–50. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, M.; Zhang, T.; Jiang, Y.; Zhao, P.; Liu, X.; Gao, A.; Yang, L.; Hou, J. Fear stress during pregnancy affects placental m6A-modifying enzyme expression and epigenetic modification levels. Front. Genet. 2022, 13, 92761. [Google Scholar] [CrossRef]
- Isono, M.; Nakano, M.; Fukami, T.; Nakajima, M. Adenosine N(6)-methylation upregulates the expression of human CYP2B6 by altering the chromatin status. Biochem. Pharm. 2022, 205, 115247. [Google Scholar] [CrossRef]
- Nakano, M.; Nakajima, M. A-to-I RNA editing and m(6)A modification modulating expression of drug-metabolizing enzymes. Drug Metab. Dispos. 2022, 50, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, P.; Bahal, R.; Manautou, J.E.; Zhong, X.B. Ontogenic mRNA expression of RNA modification writers, erasers, and readers in mouse liver. PLoS ONE 2019, 14, e0227102. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhou, Z.; Kang, X.; Pan, L.; Liu, C.; Liang, X.; Chu, J.; Dong, S.; Li, Y.; Liu, Q.; et al. Mettl3-mediated mRNA m(6)A modification controls postnatal liver development by modulating the transcription factor Hnf4a. Nat. Commun. 2022, 13, 4555. [Google Scholar] [CrossRef]
- Hrycay, E.G.; Bandiera, S.M. Expression, function and regulation of mouse cytochrome P450 enzymes: Comparison with human P450 enzymes. Curr. Drug Metab. 2009, 10, 1151–1183. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Liu, G.; Chang, Q.; Yan, M.; Yang, K.; Li, Y.; Nie, Y.; Li, X.; Han, S.; Wang, P.; et al. Prenatal Lipopolysaccharide Exposure Alters Hepatic Drug-Metabolizing Enzyme Expression in Mouse Offspring via Histone Modifications. Toxics 2023, 11, 82. https://doi.org/10.3390/toxics11010082
Zhu H, Liu G, Chang Q, Yan M, Yang K, Li Y, Nie Y, Li X, Han S, Wang P, et al. Prenatal Lipopolysaccharide Exposure Alters Hepatic Drug-Metabolizing Enzyme Expression in Mouse Offspring via Histone Modifications. Toxics. 2023; 11(1):82. https://doi.org/10.3390/toxics11010082
Chicago/Turabian StyleZhu, Hanhan, Guangming Liu, Qi Chang, Mengyao Yan, Kun Yang, Yanxin Li, Yali Nie, Xiaotian Li, Shengna Han, Pei Wang, and et al. 2023. "Prenatal Lipopolysaccharide Exposure Alters Hepatic Drug-Metabolizing Enzyme Expression in Mouse Offspring via Histone Modifications" Toxics 11, no. 1: 82. https://doi.org/10.3390/toxics11010082
APA StyleZhu, H., Liu, G., Chang, Q., Yan, M., Yang, K., Li, Y., Nie, Y., Li, X., Han, S., Wang, P., & Zhang, L. (2023). Prenatal Lipopolysaccharide Exposure Alters Hepatic Drug-Metabolizing Enzyme Expression in Mouse Offspring via Histone Modifications. Toxics, 11(1), 82. https://doi.org/10.3390/toxics11010082