



Modeling DNA Methylation Profiles and Epigenetic Analysis of Safflower (Carthamus tinctorius L.) Seedlings Exposed to Copper Heavy Metal



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Hydroponic Growth of Safflower Seedlings and Cu Stress Treatments
2.2. DNA Isolation and PCR Amplification Protocols
2.3. CRED-RA Assay
2.4. Electrophoresis
2.5. Statistical Analysis
2.6. Detection of Methylation Patterns by CRED-RA Analysis
3. Results
3.1. Data Analysis of RAPD-PCR Results
3.2. CRED-RA Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, P.H. Flora of Turkey and the East Aegeans Islands; The University Press: Edinburg, UK, 1975; Volume 5. [Google Scholar]
- Singh, R.J.; Jauhar, P.P. Genetic Resources, Chromosome Engineering, and Crop Improvement: Cereals; CRC Press: Boca Raton, FL, USA, 2006; Volume 2, pp. 167–194. [Google Scholar] [CrossRef]
- Babaoglu, M. Safflower and Its Cultivation; Trakya Agricultural Research Institute: Edirne, Türkiye, 2007. [Google Scholar]
- Delshad, E.; Yousefi, M.; Sasannezhad, P.; Rakhshandeh, H.; Ayati, Z. Medical uses of Carthamus tinctorius L. (Safflower): A comprehensive review from traditional medicine to modern medicine. Electron. Physician 2018, 10, 6672–6681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anonymous. Available online: https://www.agroscope.admin.ch/agroscope/en/home/topics/plantproduction/fieldcrops/kulturarten/alternativekulturpflanzen/saflor.html (accessed on 26 July 2021).
- Buyuk, I.; Bolukbasi, E.; Aras, E.S. Expression of CtFAD2 gene for early selection in safflower oleic linoleic oil content. J. Anim. Plant Sci. 2016, 26, 1383–1388. Available online: https://www.thejaps.org.pk/docs/v-26-05/27.pdf (accessed on 16 August 2022).
- Khurana, N.; Chatterjee, C. Influence of variable zinc on yield, oil content, and physiology of sunflower. Commun. Soil Sci. Plant Anal. 2001, 32, 3023–3030. [Google Scholar] [CrossRef]
- Yarsan, E.; Bilgili, A.; Türel, I. Heavy metal levels in mussels (Unio stevenianus Krynicki) obtained from Van Lake. Turk. J. Vet. Anim. Sci. 2000, 24, 93–96. Available online: https://www.journals.tubitak.gov.tr/veterinary/vol24/iss1/13 (accessed on 27 December 2021).
- Hu, H. Heavy metal poisoning. In Harrison’s Principles of Internal Medicine, 16th ed.; Kasper, D.L., Ed.; McGraw-Hill: New York, NY, USA, 2005; Volume 4, pp. 2577–2580. [Google Scholar]
- Kumar, N.; Soni, H.; Kumar, R.N.; Bhatt, I. Hyper accumulation and mobility of heavy metals in vegetable crops in India. J. Agric. Environ. 2009, 10, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Bolukbasi, E.; Aras, E.S. Determination of DNA methylation levels with CRED-RA technique in the genome of sunflower seedlings (Helianthus annuus L.) subjected to zinc stress. Int. J. Environ. Agric. Biotechnol. 2016, 1, 438–444. [Google Scholar] [CrossRef]
- Yang, M.G.; Lin, X.Y.; Yang, X.E. Impact of Cd on growth and nutrient accumulation of different plant species. Chin. J. Appl. Ecol. 1998, 19, 89–94. Available online: https://www.cjae.net/EN/abstract/abstract5053.shtml (accessed on 25 March 2022).
- Kosnett, M.J. Heavy metal intoxication and chelators. In Basic and Clinical Pharmacology, 10th ed.; Katzung, B.G., Ed.; McGraw-Hill: New York, NY, USA, 2007; pp. 945–957. [Google Scholar]
- Waisberg, M.P.; Joseph, B.; Beyersmann, D. Molecular and cellular mechanisms of cadmium carcinogenesis. Toxicology 2003, 192, 95–117. [Google Scholar] [CrossRef]
- Jia, X.; Fu, T.; Hu, B.; Shi, Z.; Zhou, L.; Zhu., Y. Identification of the potential risk areas for soil heavy metal pollution based on the source-sink theory. J. Hazard. Mater. 2020, 393, 122424. [Google Scholar] [CrossRef]
- Dash, S.; Borah, S.S.; Kalamdhad, A.S. Heavy metal pollution and potential ecological risk assessment for surficial sediments of Deepor Beel, India. Ecol. Indic. 2021, 122, 107265. [Google Scholar] [CrossRef]
- Bolukbasi, E. Analysis of genetic and epigenetic effects of sunflower (Helianthus Annuus L.) seedlings in response to copper stress. Fresenius Environ. Bull. 2022, 31, 4596–4602. [Google Scholar]
- Yalcin, I.E.; Ozyigit, I.I.; Dogan, I.; Demir, G.; Yarci, C. Using the Turkish red pine tree to monitor heavy metal pollution. Pol. J. Environ. Stud. 2020, 29, 3881–3889. [Google Scholar] [CrossRef]
- Alirzayeva, E.G.; Shirvani, T.S.; Yazici, M.A.; Alverdiyeva, S.; Shukurov, E.S.; Ozturk, L.; Ali-Zade, V.M.; Cakmak, I. Heavy metal accumulation in Artemisia and foliaceous lichen species from the Azerbaijan flora. For. Snow Landsc. Res. 2006, 80, 339–348. Available online: https://:www.research.sabanciuniv.edu/id/eprint/114 (accessed on 5 September 2022).
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Kirbag, F.; Munzuroglu, O. Toxic effects of cadmium (Cd+2) on metabolism of sunflower (Helianthus annuus L.) seedlings. Acta Agric. Scand. Sect. B Soil Plant Sci. 2006, 56, 224–229. [Google Scholar] [CrossRef]
- Schützendübel, A.; Polle, A. Plant responses to abiotic stresses: Heavy metal induced oxidative stress and protection by mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef]
- Soydam, A.S.; Gokce, E.; Buyuk, I.; Aras, S. Characterization of stress induced by copper and zinc on cucumber (Cucumis sativus L.) seedlings by means of molecular and population parameters. Mutat. Res. 2012, 746, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I.; Stout, P.R. The essentiality of certain elements in minute quantity for plants with special reference to copper. Plant Physiol. 1939, 14, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolukbasi, E.; Aras, E.S. Determination of physiological biochemical and molecular effects of zinc stress on the growth of sunflower seedlings (Helianthus annuus L.). Int. J. Environ. Agric. Biotechnol. 2018, 3, 530–536. [Google Scholar] [CrossRef]
- Martin, C.; Zhang, Y. Mechanisms of epigenetic inheritance. Curr. Opin. Cell Biol. 2007, 19, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Leljak-Levanic, D.; Bauer, N.; Mihaljevic, S.; Jelaska, S. Changes in DNA methylation during somatic embryogenesis in Cucurbita pepo L. Plant Cell Rep. 2004, 23, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Rein, T.; DePamphilis, M.L.; Zorbas, H. Identifying 5-methylcytosine and related modifications in DNA genomes. Nucleic Acids Res. 1998, 26, 2255–2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Li, P.; Qi, X.; Zhou, Q.; Zheng, L.; Sun, T.; Yang, Y. DNA changes in barley (Hordeum vulgare) seedlings induced by cadmium pollution using RAPD analysis. Chemosphere 2005, 61, 158–167. [Google Scholar] [CrossRef]
- Wang, W.S.; Pan, Y.J.; Zhao, X.Q.; Dwivedi, D.; Zhu, L.H.; Ali, J.; Fu, B.Y.; Li, Z.K. Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J. Exp. Bot. 2011, 62, 1951–1960. [Google Scholar] [CrossRef]
- Shams, M.; Yildirim, E.; Arslan, E.; Agar, G. Salinity induced alteration in DNA methylation pattern, enzyme activity, nutrient uptake and H2O2 content in pepper (Capsicum annuum L.) cultivars. Acta Physiol. Plant. 2020, 42, 1–12. [Google Scholar] [CrossRef]
- Grigg, G.; Clark, S. Genes and genomes: Sequencing 5-methylcytosine residues in genomic DNA. Bioessays 1994, 16, 431–436. [Google Scholar] [CrossRef]
- Altunkaynak, E.; Büyük, I.; Aydin, S.S.; Aras, S. New insight into evaluation of DNA methylation levels with CRED–RA technique in the genome of Lycopersicum esculentum subjected to NaCl and PEG. Biol. Divers. Conserv. 2016, 9, 163–171. [Google Scholar]
- Cai, Q.; Guy, C.L.; Moore, G.A. Detection of cytosine methylation and mapping of a gene influencing cytosine methylation in the genome of Citrus. Genome 1996, 39, 235–242. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. In Circular; California Agricultural Experiment Station: Davis, CA, USA, 1950; Volume 347, p. 32. [Google Scholar]
- Taspinar, M.S.; Aydin, M.; Sigmaz, B.; Yildirim, N.; Agar, G. Protective role of humic acids against picloram-induced genomic instability and DNA methylation in Phaseolus vulgaris. Environ. Sci. Pollut. Res. 2017, 24, 22948–22953. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, W.; Zhao, X.; Zhu, L.; Fu, B.; Li, Z. DNA methylation alterations of rice in response to cold stress. Plant Omics J. 2011, 4, 364–369. Available online: https://:www.pomics.com/december2011.html (accessed on 5 January 2022).
- Karan, R.; DeLeon, T.; Biradar, H.; Subudhi, P.K. Salt stress induced variation in DNA methylation pattern and its influence on gene expression in contrasting rice genotypes. PLoS ONE 2012, 7, e40203. [Google Scholar] [CrossRef]
- Bolukbasi, E. Methylation modelling and epigenetic analysis of sunflower (Helianthus annuus L.) seedlings exposed to cadmium heavy metal stress. KSU J. Agric. Nat. 2021, 25, 467–475. [Google Scholar] [CrossRef]
- Liu, W.; Yang, Y.S.; Li, P.J.; Zhou, Q.X.; Xie, L.J.; Han, Y.P. Risk assessment of cadmium contaminated soil on plant DNA damage using RAPD and physiological indices. J. Hazard. Mater. 2009, 161, 878–883. [Google Scholar] [CrossRef]
- Mastin, B.J.; Rodgers, J.H. Toxicity and bioavailability of copper herbicides (Clearigate, Cutrine-Plus, and Copper Sulfate) to freshwater animals. Environ. Contam. Toxicol. 2014, 39, 445–451. [Google Scholar] [CrossRef]
- Tuteja, N.; Ahmad, P.; Panda, B.B.; Tuteja, R. Genotoxic stress in plants: Shedding light on DNA damage, repair and DNA repair helicases. Mutat Res. 2009, 681, 134–149. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Liu, W.; Yang, Y.S.; Zhou, Q.X.; Xie, L.J.; Li, P.J.; Sun, T.H. Impact assessment of cadmium contamination on rice (Oryza sativa L.) seedlings at molecular and population levels using multiple biomarkers. Chemosphere 2007, 67, 1155–1163. [Google Scholar] [CrossRef]
- Taspinar, M.S.; Agar, G.; Alpsoy, L.; Yildirim, N.; Bozari, S. The protective role of zinc and calcium in Vicia faba seedlings subjected to cadmium stress. Toxicol. Ind. Health 2011, 27, 73–80. [Google Scholar] [CrossRef]
- Atienzar, F.A.; Cordi, B.; Donkin, M.E.; Evenden, A.J. Comparison of ultravioletinduced genotoxicity detected by random amplified polymorphic DNA with chlorophyll fluorescence and growth in a marine macroalgae. Palmaria Palmata. Aquat Toxicol 2000, 50, 1–12. [Google Scholar] [CrossRef]
- Gallo-Franco, J.J.; Sosa, C.C.; Ghneim-Herrera, T.; Quimbaya, M. Epigenetic control of plant response to heavy metal stress: A new view on aluminum tolerance. Front. Plant Sci. 2020, 11, 602625. [Google Scholar] [CrossRef] [PubMed]
- Harshitha, K.; Nair, R.A. Evaluation of DNA methylation changes by CRED-RA analysis following prednisone treatment of endophyte, Fusarium oxysporum. Indian J. Microbiol. 2020, 60, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Savva, D. The use of arbitrarily primed PCR (AP-PCR) fingerprinting to detect exposure to genotoxic chemicals. Ecotoxicology 2000, 9, 341–353. [Google Scholar] [CrossRef]
- Shi, D.; Zhuang, K.; Xia, Y.; Zhu, C.; Chen, C.; Hu, Z.; Shen, Z. Hydrilla verticillata employs two different ways to affect DNA methylation under excess copper stress. Aquat. Toxicol. 2017, 193, 97–104. [Google Scholar] [CrossRef]
- Greco, M.; Sáez, C.A.; Contreras, R.A.; Brown, M.T. Cadmium and/or copper excess induce interdependent metal accumulation, DNA methylation, induction of metal chelators and antioxidant defences in the seagrass Zostera marina. Chemosphere 2019, 224, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boquete, M.T.; Schmid, M.W.; Wagemaker, N.C.; Carey, S.B.; Alonso, C. Molecular basis of intraspecific differentiation for heavy metal tolerance in the copper moss Scopelophila cataractae. Environ. Exp. Bot. 2022, 201, 104970. [Google Scholar] [CrossRef]
- Choi, C.S.; Sano, H. Abiotic-stress induces demethylation and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants. Mol. Genet. Genom. 2007, 277, 589–600. [Google Scholar] [CrossRef]
- Ertürk, F.A.; Sunar, S.; Agar, G. Determination of cytogenetic and epigenetic effects of manganese and copper on Zea mays L. ISPEC J. Agric. Sci. 2021, 5, 529–543. [Google Scholar] [CrossRef]
- Ou, X.; Zhang, Y.; Xu, C.; Lin, X.; Zang, Q.; Zhuang, T.; Liu, B. Transgenerational inheritance of modified DNA methylation patterns and enhanced tolerance induced by heavy metal stress in rice (Oryza sativa L.). PLoS ONE 2012, 7, e41143. [Google Scholar] [CrossRef] [Green Version]
- McKergow, M.; Nkongolo, K.K. Gene regulation and global DNA methylation changes in white spruce (Picea glauca) in response to copper contaminations. Water Air Soil Pollut. 2023, 234, 1–23. [Google Scholar] [CrossRef]
- Yagci, S.; Yildirim, E.; Yildirim, N. Nitric oxide alleviates the effects of copper-induced DNA methylation, genomic instability, LTR retrotransposon polymorphism and enzyme activity in lettuce. Plant Physiol. Rep. 2019, 24, 289–295. [Google Scholar] [CrossRef]
- Colzi, I.; Arnetoli, M.; Gallo, A.; Doumett, S.; Gonnelli, C. Copper tolerance strategies involving the root cell wall pectins in Silene paradoxa L. Environ. Exp. Bot. 2012, 78, 91–98. [Google Scholar] [CrossRef]
- Cong, W.; Miao, Y.; Xu, L. Transgenerational memory of gene expression changes induced by heavy metal stress in rice (Oryza sativa L.). BMC Plant Biol. 2019, 19, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Methylation Status | HpaII | MspI | Score of Band Profile | ||||
|---|---|---|---|---|---|---|---|---|
| x | y | z | ||||||
| Type I | CCGG GGCC | digestion | digestion | −/1 | +/0 | +/0 | Non-methylation | |
| Type II | CCGG GGCC | CCGG GGCC | digestion | undigestion | −/1 | +/0 | −/1 | Semi-methylation |
| Type III | CCGG GGCC | undigestion | digestion | −/1 | −/1 | +/0 | Full-methylation | |
| Type IV | CCGG GGCC | undigestion | undigestion | −/1 | −/1 | −/1 | Full-methylation | |
| Primers’ Name | Polymorphism Rates (%) | Sequences (5′→3′) |
|---|---|---|
| OPC-01 * | 16.90 | TTCGAGCCAG |
| OPC-02 * | 14.40 | GTGAGGCGTC |
| OPC-04 * | 36.40 | CCGCATCTAC |
| OPC-06 | 52.90 | GAACGGACTC |
| OPC-07 * | 56.80 | GTCCCGACGA |
| OPC-08 | 53.00 | TGGACCGGTG |
| OPC-09 * | 59.70 | CTCACCGTCC |
| OPC-10 * | 54.10 | TGTCTGGGTG |
| OPC-11 | 55.90 | AAAGCTGCGG |
| OPA-08 | 32.50 | GTGACGTAGG |
| Samples | GTS Rate (%) |
|---|---|
| 20 mg L−1 | 82.75 |
| 40 mg L−1 | 88.90 |
| 80 mg L−1 | 86.26 |
| 160 mg L−1 | 83.90 |
| 320 mg L−1 | 82.25 |
| 640 mg L−1 | 84.60 |
| 1280 mg L−1 | 83.70 |
| Control | 20 mg L−1 | 40 mg L−1 | 80 mg L−1 | 160 mg L−1 | 320 mg L−1 | 640 mg L−1 | 1280 mg L−1 | |
|---|---|---|---|---|---|---|---|---|
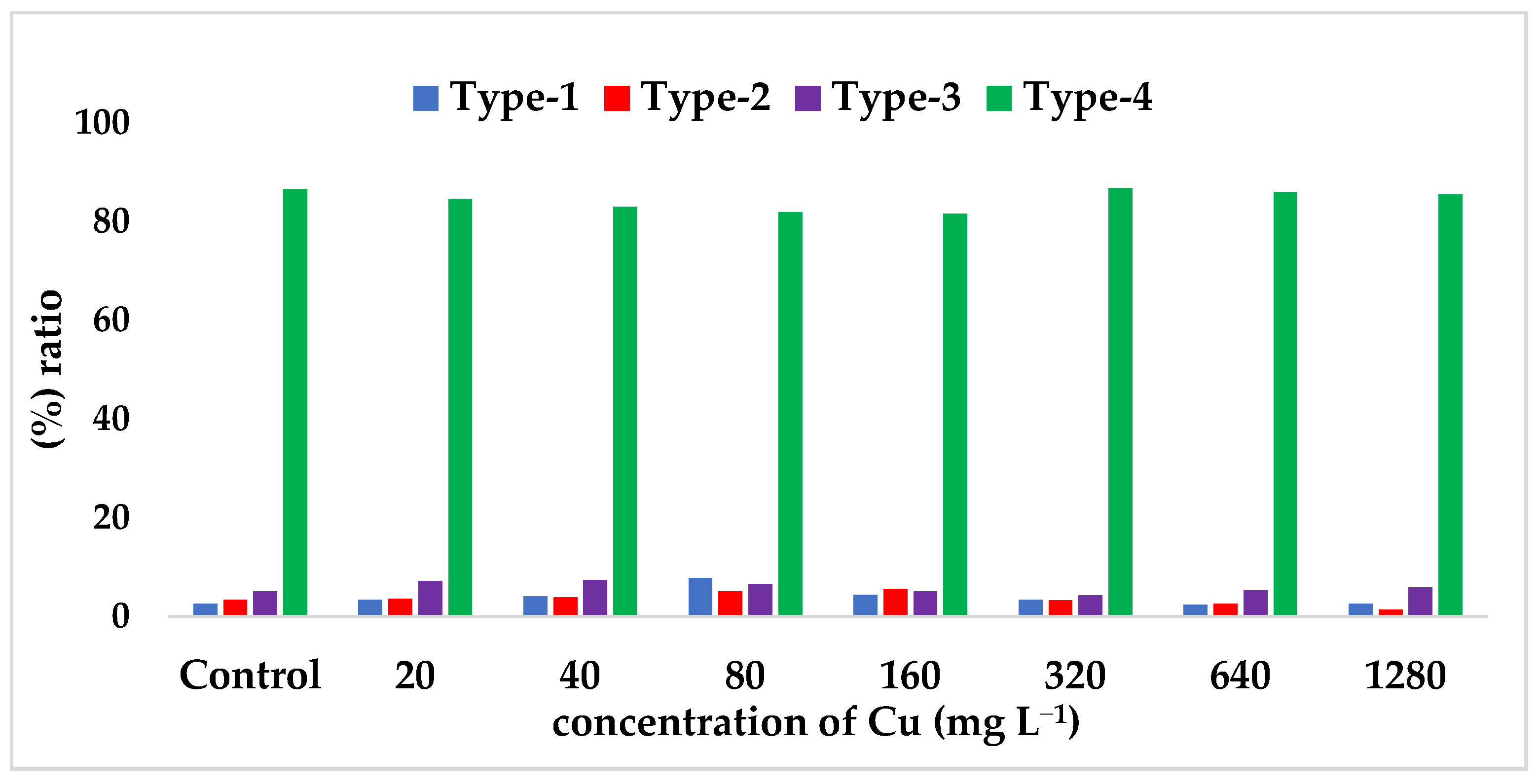
| Type-I (%) (Non-methylation) | 2.60 | 3.40 | 4.10 | 7.80 * | 4.40 | 3.40 | 2.40 * | 2.60 |
| Type-II (%) | 3.40 | 3.60 | 3.90 | 5.10 | 5.60 * | 3.30 | 2.60 | 1.40 * |
| Type-III (%) | 5.10 | 7.20 | 7.40 | 6.60 | 5.10 | 4.30 | 5.30 | 5.90 |
| Type-IV (%) | 86.60 * | 84.60 | 83.00 | 81.90 * | 81.60 * | 86.80 * | 86.00 | 85.50 |
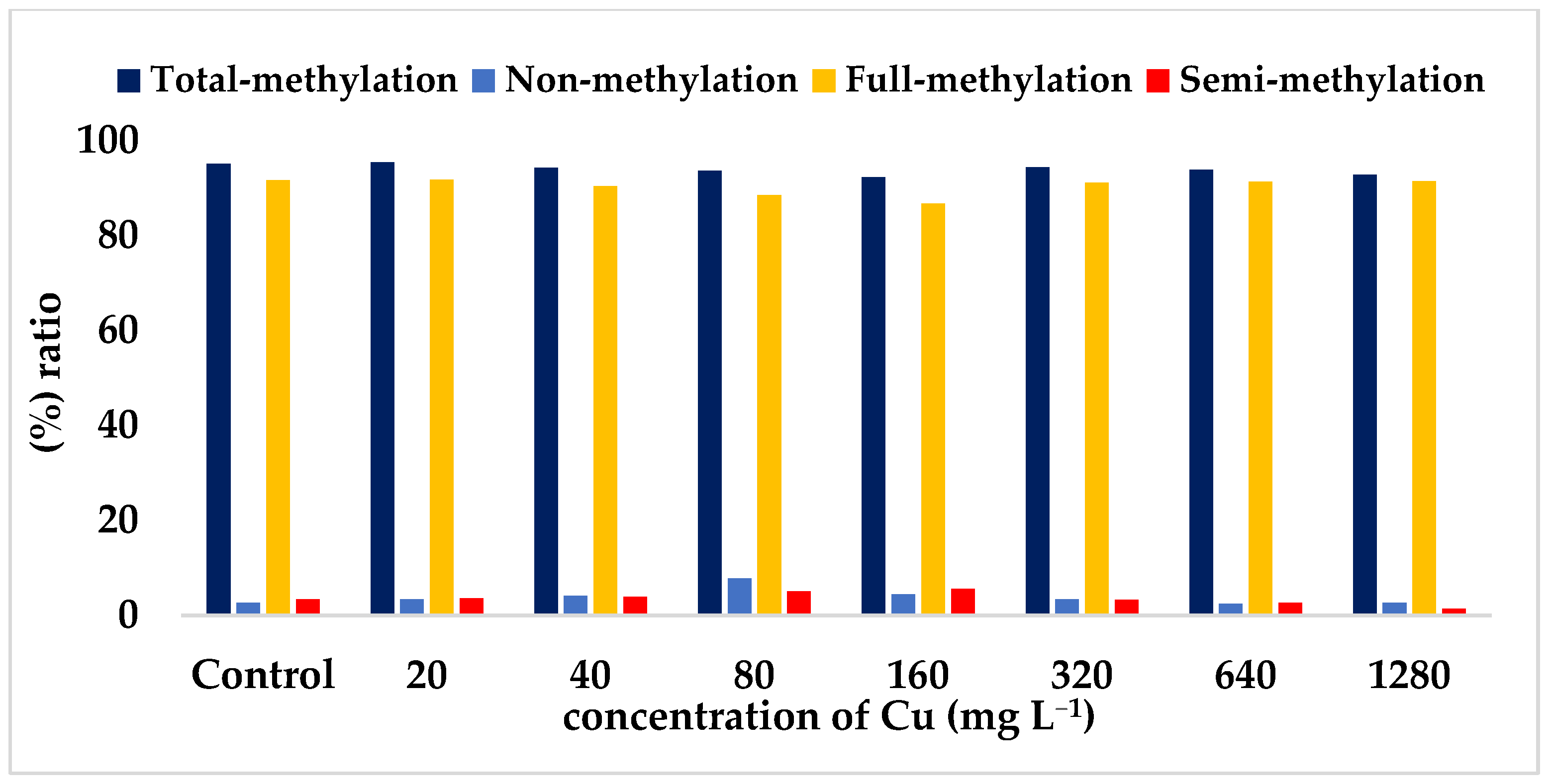
| Total methylated bands ratio (%) a | 95.10 | 95.40 * | 94.30 | 93.60 | 92.30 * | 94.40 | 93.90 | 92.80 |
| Full-methylated bands ratio (%) b | 91.70 | 91.80 * | 90.40 | 88.50 | 86.70 * | 91.10 | 91.30 | 91.40 |
| Semi-methylated bands ratio (%) c | 3.40 | 3.60 | 3.90 | 5.10 | 5.60 * | 3.30 | 2.60 * | 1.40 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bölükbaşı, E.; Karakaş, M. Modeling DNA Methylation Profiles and Epigenetic Analysis of Safflower (Carthamus tinctorius L.) Seedlings Exposed to Copper Heavy Metal. Toxics 2023, 11, 255. https://doi.org/10.3390/toxics11030255
Bölükbaşı E, Karakaş M. Modeling DNA Methylation Profiles and Epigenetic Analysis of Safflower (Carthamus tinctorius L.) Seedlings Exposed to Copper Heavy Metal. Toxics. 2023; 11(3):255. https://doi.org/10.3390/toxics11030255
Chicago/Turabian StyleBölükbaşı, Ekrem, and Mehmet Karakaş. 2023. "Modeling DNA Methylation Profiles and Epigenetic Analysis of Safflower (Carthamus tinctorius L.) Seedlings Exposed to Copper Heavy Metal" Toxics 11, no. 3: 255. https://doi.org/10.3390/toxics11030255
APA StyleBölükbaşı, E., & Karakaş, M. (2023). Modeling DNA Methylation Profiles and Epigenetic Analysis of Safflower (Carthamus tinctorius L.) Seedlings Exposed to Copper Heavy Metal. Toxics, 11(3), 255. https://doi.org/10.3390/toxics11030255