
Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella subcapitata to Cerium Oxide Nanoparticles


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of nCeO2
2.2. Preparation of Exposure Media and Concentrations
2.3. Test Organisms
2.4. Cytotoxic Effects of nCeO2 on P. subcapitata
2.5. DNA Damage and Estimation of Genomic Template Stability
2.6. Data Analysis
3. Results and Discussion
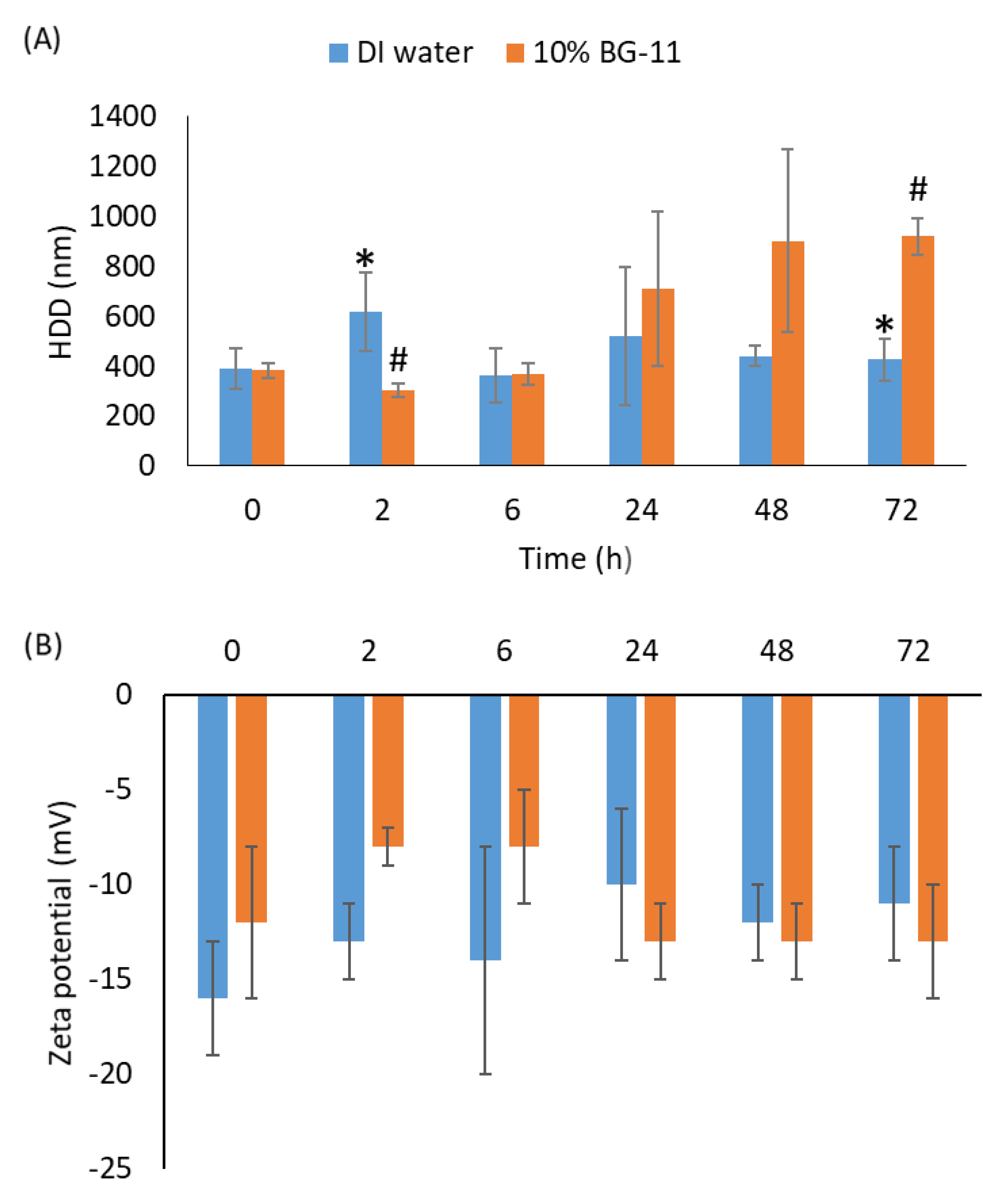
3.1. Characterization of nCeO2
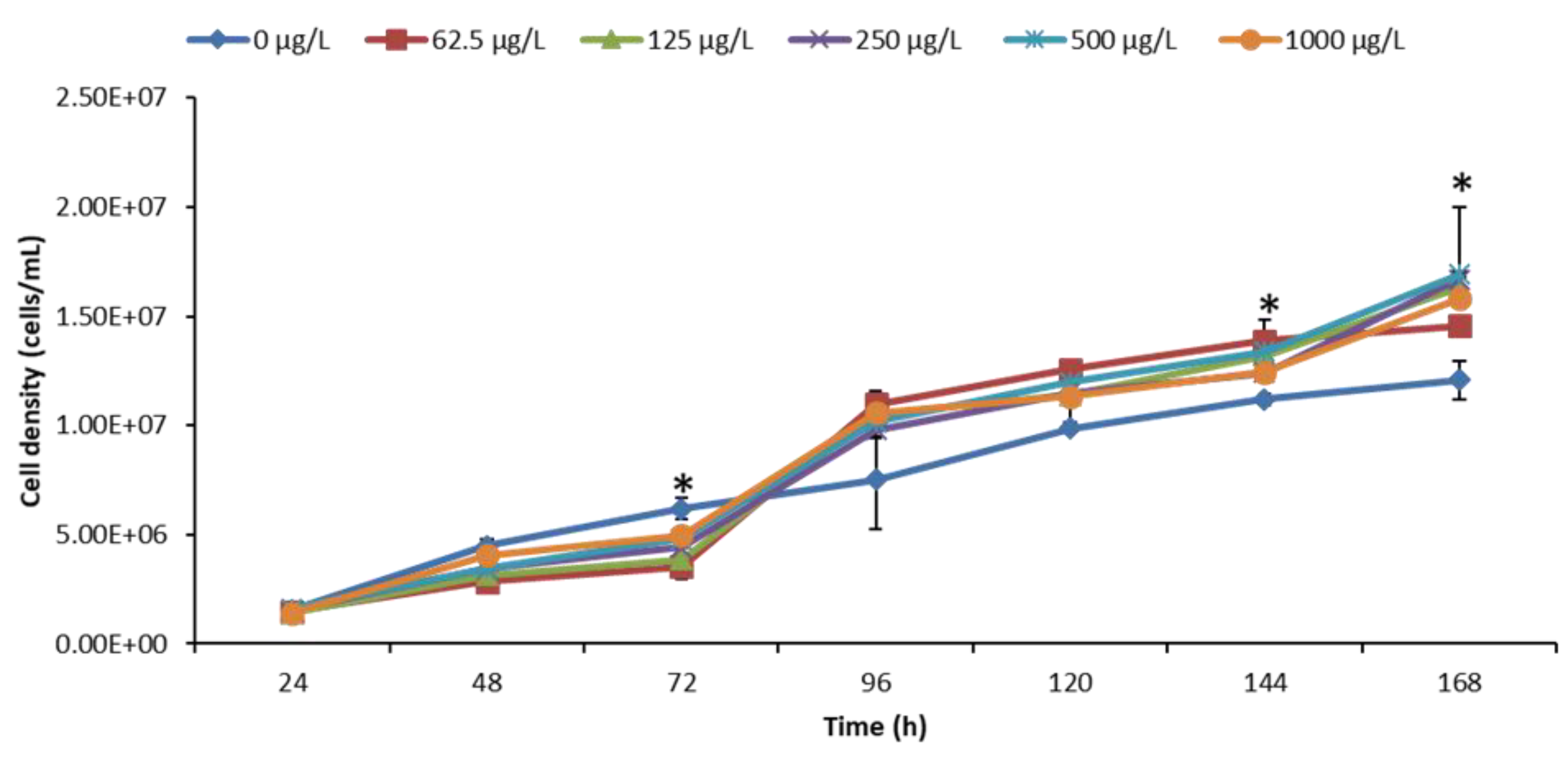
3.2. Effect of nCeO2 on Algal Growth
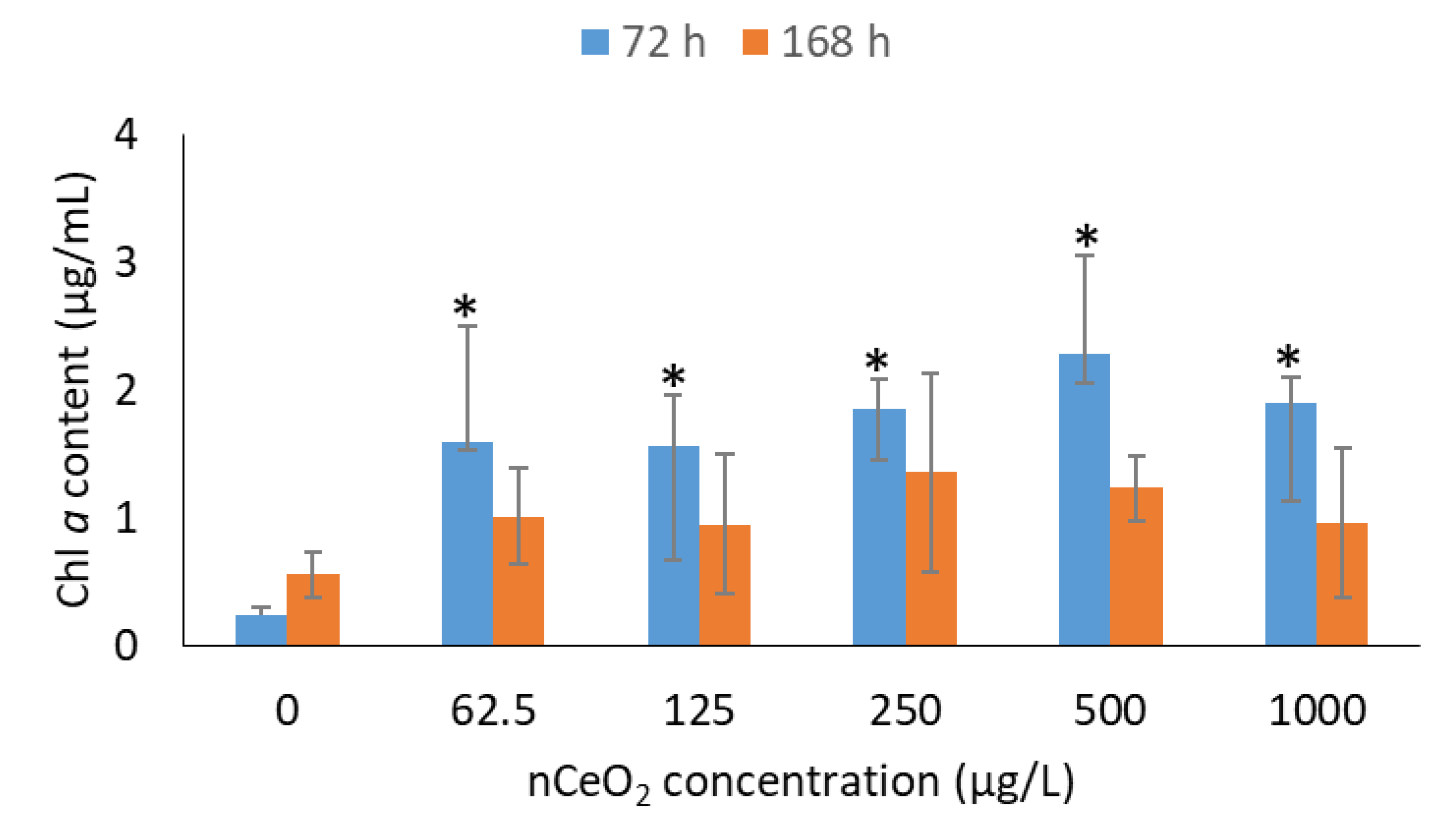
3.3. Effect of nCeO2 on Chl a Content
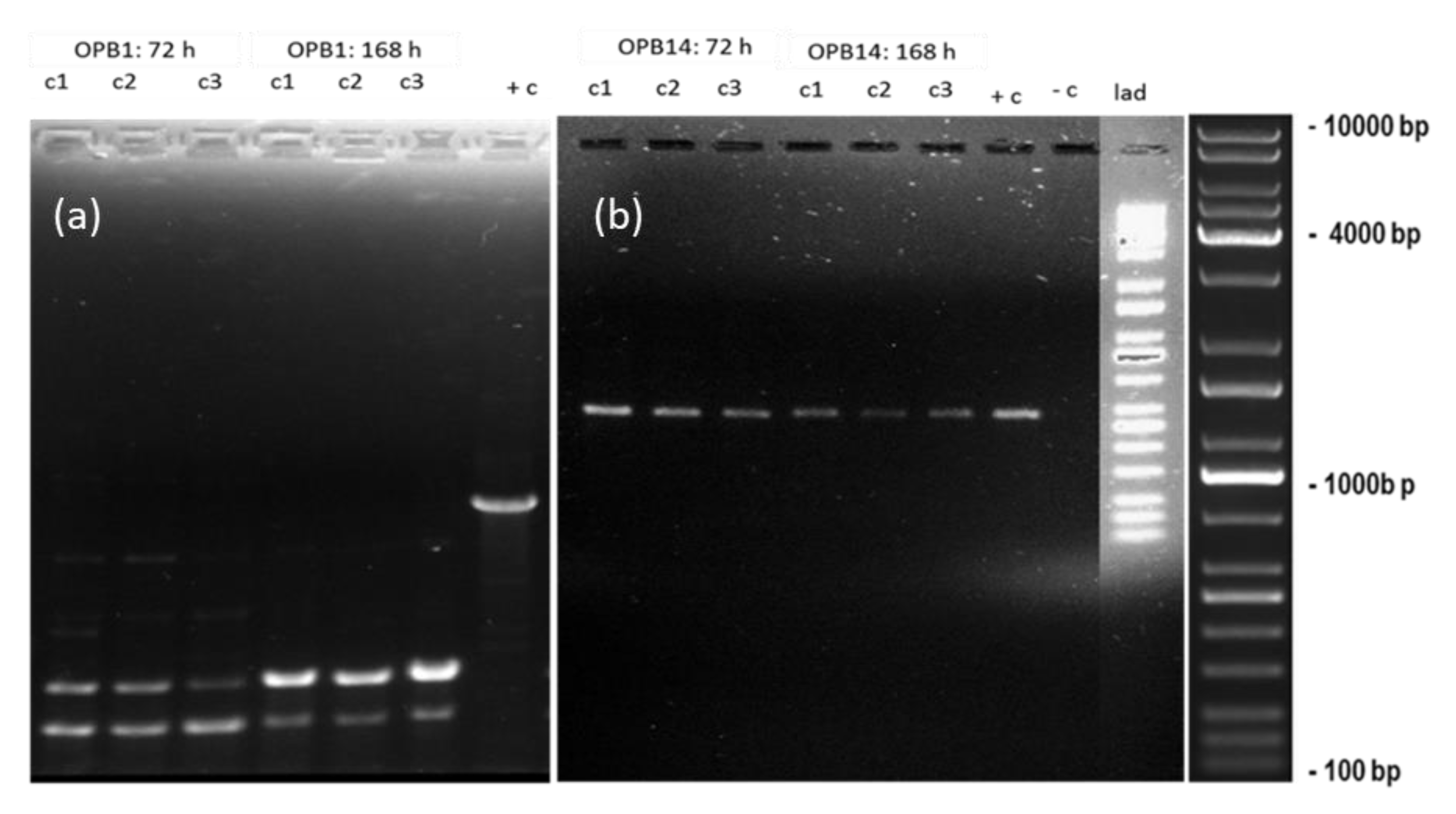
3.4. DNA Damage and Estimation of Genomic Template Stability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, K.R.; Nayak, V.; Sarkar, T.; Singh, R.P. Cerium oxide nanoparticles: Properties, biosynthesis and biomedical application. RSC Adv. 2020, 10, 27194–27214. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, S.; Liao, S.; Xiang, F.; Wu, X. Preparation of new sunscreen materials Ce1- x Zn x O2- x via solid-state re-action at room temperature and study on their properties. Rare Metals 2010, 29, 149–153. [Google Scholar] [CrossRef]
- Cassee, F.R.; Campbell, A.; Boere, A.J.F.; McLean, S.G.; Duffin, R.; Krystek, P.; Gosens, I.; Miller, M.R. The biological effects of subacute inhalation of diesel exhaust following addition of cerium oxide nanoparticles in atherosclerosis-prone mice. Environ. Res. 2012, 115, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Trovarelli, A. Catalytic Properties of Ceria and CeO2-Containing Materials. Catal. Rev. 1996, 38, 439–520. [Google Scholar] [CrossRef]
- Wang, C.-H.; Lin, S.-S. Preparing an active cerium oxide catalyst for the catalytic incineration of aromatic hydrocar-bons. Appl. Catal. A Gen. 2004, 268, 227–233. [Google Scholar] [CrossRef]
- Xu, C.; Qu, X. Cerium oxide nanoparticle: A remarkably versatile rare earth nanomaterial for biological applications. NPG Asia Mater. 2014, 6, e90. [Google Scholar] [CrossRef] [Green Version]
- Caputo, F.; De Nicola, M.; Ghibelli, L. Pharmacological potential of bioactive engineered nanomaterials. Biochem. Pharmacol. 2014, 92, 112–130. [Google Scholar] [CrossRef] [PubMed]
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nano-materials in Europe and the world. J. Nanoparticle Res. 2012, 14, 1109. [Google Scholar] [CrossRef] [Green Version]
- Collin, B.; Auffan, M.; Johnson, A.C.; Kaur, I.; Keller, A.A.; Lazareva, A.; Lead, J.R.; Ma, X.; Merrifield, R.C.; Svendsen, C.; et al. Environmental release, fate and ecotoxicological effects of manufactured ceria nanomaterials. Environ. Sci. Nano 2014, 1, 533–548. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.A.; Lazareva, A. Predicted Releases of Engineered Nanomaterials: From Global to Regional to Local. Environ. Sci. Technol. Lett. 2013, 1, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.C.; Park, B. Predicting contamination by the fuel additive cerium oxide engineered nanoparticles within the United Kingdom and the associated risks. Environ. Toxicol. Chem. 2012, 31, 2582–2587. [Google Scholar] [CrossRef]
- Gottschalk, F.; Sun, T.; Nowack, B. Environmental concentrations of engineered nanomaterials: Review of modeling and analytical studies. Environ. Pollut. 2013, 181, 287–300. [Google Scholar] [CrossRef]
- Peters, R.J.; van Bemmel, G.; Milani, N.B.; Hertog, G.C.D.; Undas, A.; van der Lee, M.; Bouwmeester, H. Detection of nanoparticles in Dutch surface waters. Sci. Total. Environ. 2017, 621, 210–218. [Google Scholar] [CrossRef]
- OECD. List of Manufactured Nanomaterials and List of Endpoints for Phase One of the Sponsorship Programme for the Testing of Manufactured Nanomaterials: Revision. 2010. Available online: https://www.oecd.org/chemicalsafety/nanosafety/testing-programme-manufactured-nanomaterials.htm (accessed on 15 January 2023).
- Abbas, Q.; Liu, G.; Yousaf, B.; Ali, M.U.; Ullah, H.; Munir, M.A.M.; Ahmed, R.; Rehman, A. Biochar-assisted transformation of engineered-cerium oxide nanoparticles: Effect on wheat growth, photosynthetic traits and cerium accumulation. Ecotoxicol. Environ. Saf. 2019, 187, 109845. [Google Scholar] [CrossRef]
- Pulido-Reyes, G.; Briffa, S.M.; Hurtado-Gallego, J.; Yudina, T.; Leganés, F.; Puntes, V.; Valsami-Jones, E.; Rosal, R.; Fernández-Piñas, F. Internalization and toxicological mechanisms of uncoated and PVP-coated cerium oxide nanoparticles in the freshwater algaChlamydomonas reinhardtii. Environ. Sci. Nano 2019, 6, 1959–1972. [Google Scholar] [CrossRef]
- Saison, C.; Perreault, F.; Daigle, J.-C.; Fortin, C.; Claverie, J.; Morin, M.; Popovic, R. Effect of core–shell copper oxide nanoparticles on cell culture morphology and photosynthesis (photosystem II energy distribution) in the green alga, Chlamydomonas reinhardtii. Aquat. Toxicol. 2010, 96, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.; Størseth, T.; Altin, D.; Fornara, A.; Ahniyaz, A.; Jungnickel, H.; Laux, P.; Luch, A.; Sørensen, L. Freshwater dispersion stability of PAA-stabilised cerium oxide nanoparticles and toxicity towards Pseudokirchneriella subcapitata. Sci. Total. Environ. 2015, 505, 596–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodea-Palomares, I.; Gonzalo, S.; Santiago-Morales, J.; Leganés, F.; García-Calvo, E.; Rosal, R.; Fernández-Piñas, F. An insight into the mechanisms of nanoceria toxicity in aquatic photosynthetic organisms. Aquat. Toxicol. 2012, 122–123, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, J.; Du, W.; Yin, Y.; Guo, H. Toxicity mechanism of cerium oxide nanoparticles on cyanobacteria Microcystis aeruginosa and their ecological risks. Environ. Sci. Pollut. Res. 2022, 29, 34010–34018. [Google Scholar] [CrossRef]
- Xie, C.; Ma, Y.; Zhang, P.; Zhang, J.; Li, X.; Zheng, K.; Li, A.; Wu, W.; Pang, Q.; He, X. Elucidating the origin of the toxicity of nano-CeO2 to Chlorella pyrenoidosa: The role of specific surface area and chemical composition. Environ. Sci. Nano 2021, 8, 1701–1712. [Google Scholar] [CrossRef]
- Mahaye, N.; Thwala, M.; Cowan, D.; Musee, N. Genotoxicity of metal based engineered nanoparticles in aquatic organisms: A review. Mutat. Res. Mol. Mech. Mutagen. 2017, 773, 134–160. [Google Scholar] [CrossRef] [PubMed]
- Milani, Z.M.; Charbgoo, F.; Darroudi, M. Impact of physicochemical properties of cerium oxide nanoparticles on their toxicity effects. Ceram. Int. 2017, 43, 14572–14581. [Google Scholar] [CrossRef]
- Lee, S.-W.; Kim, S.-M.; Choi, J. Genotoxicity and ecotoxicity assays using the freshwater crustacean Daphnia magna and the larva of the aquatic midge Chironomus riparius to screen the ecological risks of nanoparticle exposure. Environ. Toxicol. Pharmacol. 2009, 28, 86–91. [Google Scholar] [CrossRef]
- Bayat, N. Toxicity and Biological Impact of Metal and Metal Oxide Nanoparticles Focus on the Vascular Toxicity of ul-tra-Small Titanium Dioxide Nanoparticles. Department of Biochemistry and Biophysics, Stockholm University: Stockholm, Sweden, 2015. [Google Scholar]
- Taylor, N.S.; Merrifield, R.; Williams, T.D.; Chipman, J.K.; Lead, J.R.; Viant, M.R. Molecular toxicity of cerium oxide nanoparticles to the freshwater alga Chlamydomonas reinhardtii is associated with supra-environmental exposure concentra-tions. Nanotoxicology 2016, 10, 32–41. [Google Scholar]
- Mahaye, N.; Leareng, S.K.; Musee, N. Cytotoxicity and genotoxicity of coated-gold nanoparticles on freshwater algae Pseudokirchneriella subcapitata. Aquat. Toxicol. 2021, 236, 105865. [Google Scholar] [CrossRef] [PubMed]
- Mahana, A.; Guliy, O.I.; Mehta, S.K. Accumulation and cellular toxicity of engineered metallic nanoparticle in freshwater microalgae: Current status and future challenges. Ecotoxicol. Environ. Saf. 2021, 208, 111662. [Google Scholar] [CrossRef]
- Yingjun, W.; Jia, L.; Yun, L.Ü.; Hangbiao, J.; Shihuai, D.; Yunmin, Z. Effects of cerium on growth and physiological characteristics of Anabaena flosaquae. J. Rare Earths 2012, 30, 1287–1292. [Google Scholar]
- Xue, W.; Yousheng, L.; Dongwu, L.; Hengjian, X.; Tao, L.; Fengyun, Z. Cerium toxici-ty, uptake and translocation in Arabidopsis thaliana seedlings. J. Rare Earths 2012, 30, 579–585. [Google Scholar]
- Angel, B.M.; Vallotton, P.; Apte, S.C. On the mechanism of nanoparticulate CeO2 toxicity to freshwater algae. Aquat. Toxicol. 2015, 168, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Vannini, C.; Domingo, G.; Marsoni, M.; De Mattia, F.; Labra, M.; Castiglioni, S.; Bracale, M. Effects of a complex mixture of therapeutic drugs on unicellular algae Pseudokirchneriella subcapitata. Aquat. Toxicol. 2011, 101, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocco, L.; Santonastaso, M.; Nigro, M.; Mottola, F.; Costagliola, D.; Bernardeschi, M.; Guidi, P.; Lucchesi, P.; Scarcelli, V.; Corsi, I.; et al. Genomic and chromosomal damage in the marine mussel Mytilus galloprovincialis: Effects of the combined exposure to titanium dioxide nanoparticles and cadmium chloride. Mar. Environ. Res. 2015, 111, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Rocco, L.; Valentino, I.V.; Scapigliati, G.; Stingo, V. RAPD-PCR analysis for molecular characterization and genotoxic studies of a new marine fish cell line derived from Dicentrarchus labrax. Cytotechnology 2013, 66, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Rocco, L.; Santonastaso, M.; Mottola, F.; Costagliola, D.; Suero, T.; Pacifico, S.; Stingo, V. Genotoxicity assessment of TiO2 nanoparticles in the teleost Danio rerio. Ecotoxicol. Environ. Saf. 2015, 113, 223–230. [Google Scholar] [CrossRef]
- Mahaye, N. Stability of Gold and Cerium Oxide Nanoparticles in Aqueous Environments, and Their Effects on Pseudo-kirchneriella Subcapitata and Salvinia Minima. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2019. Available online: https://repository.up.ac.za/handle/2263/72778 (accessed on 15 January 2023).
- Slabbert, L. Methods for Direct Estimation of Ecological Effect Potential (DEEEP), 1st ed.; Water Research Com-mission Report No.: 1313/01/04; Water Research Commission: Pretoria, South Africa, 2004; p. 100. [Google Scholar]
- Rodrigues, L.H.R.; Arenzon, A.; Raya-Rodriguez, M.T.; Fontoura, N.F. Algal density assessed by spectrophotometry: A cali-bration curve for the unicellular algae Pseudokirchneriella subcapitata. J. Environ. Chem. Ecotoxicol. 2011, 3, 225–228. [Google Scholar]
- Miller, W.E.; Greene, J.C.; Shiroyama, T. The Selenastrum capricornutum printz algal assay bottle test: Experimental design, application, and data interpretation protocol; Environmental Protection Agency, Office of Research and Develop-ment, Corvallis Environmenal Research Laboratory: Washington, DC, USA, 1978. [Google Scholar]
- Markus, A.; Krystek, P.; Tromp, P.; Parsons, J.; Roex, E.; de Voogt, P.; Laane, R. Determination of metal-based nanoparticles in the river Dommel in the Netherlands via ultrafiltration, HR-ICP-MS and SEM. Sci. Total. Environ. 2018, 631–632, 485–495. [Google Scholar] [CrossRef] [PubMed]
- de Klein, J.J.M.; Quik, J.T.K.; Bäuerlein, P.S.; Koelmans, A.A. Towards validation of the NanoDUFLOW nanoparticle fate model for the river Dommel, The Netherlands. Environ. Sci. Nano 2016, 3, 434–441. [Google Scholar] [CrossRef]
- Giese, B.; Klaessig, F.; Park, B.; Kaegi, R.; Steinfeldt, M.; Wigger, H.; von Gleich, A.; Gottschalk, F. Risks, Release and Concentrations of Engineered Nanomaterial in the Environment. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Grillo, R.; Clemente, Z.; de Oliveira, J.L.; Campos, E.V.R.; Chalupe, V.C.; Jonsson, C.M.; de Lima, R.; Sanches, G.; Nishisaka, C.S.; Rosa, A.H.; et al. Chitosan nanoparticles loaded the herbicide paraquat: The influence of the aquatic humic substances on the colloidal stability and toxicity. J. Hazard. Mater. 2015, 286, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Ozkaleli, M.; Erdem, A. Biotoxicity of TiO2 Nanoparticles on Raphidocelis subcapitata Microalgae Exemplified by Membrane Deformation. Int. J. Environ. Res. Public Heal. 2018, 15, 416. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.H. The Chlamydomonas Sourcebook; Academic Press: San Diego, CA, USA, 1989. [Google Scholar] [CrossRef]
- A Atienzar, F.; Venier, P.; Jha, A.; Depledge, M.H. Evaluation of the random amplified polymorphic DNA (RAPD) assay for the detection of DNA damage and mutations. Mutat. Res. Toxicol. Environ. Mutagen. 2002, 521, 151–163. [Google Scholar] [CrossRef]
- Nur, Y.; Lead, J.; Baalousha, M. Evaluation of charge and agglomeration behavior of TiO2 nanoparticles in ecotoxicological media. Sci. Total. Environ. 2015, 535, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Alam, B.; Philippe, A.; Rosenfeldt, R.R.; Seitz, F.; Dey, S.; Bundschuh, M.; Schaumann, G.E.; Brenner, S.A. Synthesis, characterization, and ecotoxicity of CeO2 nanoparticles with differing properties. J. Nanoparticle Res. 2016, 18, 1–10. [Google Scholar] [CrossRef]
- Oriekhova, O.; Stoll, S. Stability of uncoated and fulvic acids coated manufactured CeO2 nanoparticles in various conditions: From ultrapure to natural Lake Geneva waters. Sci. Total. Environ. 2016, 562, 327–334. [Google Scholar] [CrossRef]
- Yang, X.; Pan, H.; Wang, P.; Zhao, F.-J. Particle-specific toxicity and bioavailability of cerium oxide (CeO2) nanoparticles to Arabidopsis thaliana. J. Hazard. Mater. 2017, 322, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Song, U.; Shin, M.; Lee, G.; Roh, J.; Kim, Y.; Lee, E.J. Functional Analysis of TiO2 Nanoparticle Toxicity in Three Plant Species. Biol. Trace Element Res. 2013, 155, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ma, Y.; Liu, S.; Wang, G.; Zhang, J.; He, X.; Zhang, J.; Rui, Y.; Zhang, Z. Phytotoxicity, uptake and transformation of nano-CeO2 in sand cultured romaine lettuce. Environ. Pollut. 2017, 220, 1400–1408. [Google Scholar] [CrossRef]
- Rogers, N.J.; Franklin, N.M.; Apte, S.C.; Batley, G.E.; Angel, B.; Lead, J.R.; Baalousha, M. Physico-chemical behaviour and algal toxicity of nanoparticulate CeO2 in freshwater. Environ. Chem. 2010, 7, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Manier, N.; Bado-Nilles, A.; Delalain, P.; Aguerre-Chariol, O.; Pandard, P. Ecotoxicity of non-aged and aged CeO2 nanomaterials towards freshwater microalgae. Environ. Pollut. 2013, 180, 63–70. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, Z.; Zhai, Y.; Zhang, F.; Vijver, M.G.; Peijnenburg, W.J. Effects of humic substances on the aqueous sta-bility of cerium dioxide nanoparticles and their toxicity to aquatic organisms. Sci. Total Environ. 2021, 781, 146583. [Google Scholar] [CrossRef]
- Hitchman, A.; Smith, G.H.S.; Ju-Nam, Y.; Sterling, M.; Lead, J.R. The effect of environmentally relevant conditions on PVP stabilised gold nanoparticles. Chemosphere 2013, 90, 410–416. [Google Scholar] [CrossRef]
- Lowry, G.V.; Hill, R.J.; Harper, S.; Rawle, A.F.; Hendren, C.O.; Klaessig, F.; Nobbmann, U.; Sayre, P.; Rumble, J. Guidance to improve the scientific value of zeta-potential measurements in nanoEHS. Environ. Sci. Nano 2016, 3, 953–965. [Google Scholar] [CrossRef]
- Rioboo, C.; Prado, R.; Herrero, C.; Cid, A. Population growth study of the rotifer Brachionus sp. fed with triazine-exposed microalgae. Aquat. Toxicol. 2007, 83, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Dubey, D.; Dutta, V. Nutrient Enrichment in Lake Ecosystem and Its Effects on Algae and Macrophytes. Environ. Concerns Sustain. Dev. Vol. 2 Biodivers. Soil Waste Manag. 2019, 81–126. [Google Scholar] [CrossRef]
- Dedman, C.J.; Rizk, M.M.I.; Christie-Oleza, J.A.; Davies, G.-L. Investigating the Impact of Cerium Oxide Nanoparticles Upon the Ecologically Significant Marine Cyanobacterium Prochlorococcus. Front. Mar. Sci. 2021, 8, 668097. [Google Scholar] [CrossRef]
- Röhder, L.A.; Brandt, T.; Sigg, L.; Behra, R. Influence of agglomeration of cerium oxide nanoparticles and speciation of cerium(III) on short term effects to the green algae Chlamydomonas reinhardtii. Aquat. Toxicol. 2014, 152, 121–130. [Google Scholar] [CrossRef]
- Zemke-White, W.L.; Clements, K.D.; Harris, P.J. Acid lysis of macroalgae by marine herbivorous fishes: Effects of acid pH on cell wall porosity. J. Exp. Mar. Biol. Ecol. 2000, 245, 57–68. [Google Scholar] [CrossRef]
- Deng, X.-Y.; Cheng, J.; Hu, X.-L.; Wang, L.; Li, D.; Gao, K. Biological effects of TiO2 and CeO2 nanoparticles on the growth, photosynthetic activity, and cellular components of a marine diatom Phaeodactylum tricornutum. Sci. Total. Environ. 2017, 575, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wu, D.; Cao, S.; Du, W.; Yin, Y.; Guo, H. Effects of CeO2 Nanoparticles on Microcystis aeruginosa Growth and Microcystin Production. Bull. Environ. Contam. Toxicol. 2020, 104, 834–839. [Google Scholar] [CrossRef]
- Du, S.; Zhang, P.; Zhang, R.; Lu, Q.; Liu, L.; Bao, X.; Liu, H. Reduced graphene oxide induces cytotoxicity and inhibits photosynthetic performance of the green alga Scenedesmus obliquus. Chemosphere 2016, 164, 499–507. [Google Scholar] [CrossRef]
- Qian, H.; Zhu, K.; Lu, H.; Lavoie, M.; Chen, S.; Zhou, Z.; Deng, Z.; Chen, J.; Fu, Z. Contrasting silver nanoparticle toxicity and detoxification strategies in Microcystis aeruginosa and Chlorella vulgaris: New insights from proteomic and physiological analyses. Sci. Total. Environ. 2016, 572, 1213–1221. [Google Scholar] [CrossRef]
- Chiu, M.-H.; Khan, Z.A.; Garcia, S.G.; Le, A.D.; Kagiri, A.; Ramos, J.; Tsai, S.-M.; Drobenaire, H.W.; Santschi, P.; Quigg, A.; et al. Effect of Engineered Nanoparticles on Exopolymeric Substances Release from Marine Phytoplankton. Nanoscale Res. Lett. 2017, 12, 620. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Lv, J.; Ma, J.; Zhang, S. Cellular internalization and intracellular biotransformation of silver nanoparticles in Chlamydomonas reinhardtii. Nanotoxicology 2016, 10, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Metzler, D.M.; Erdem, A.; Tseng, Y.H.; Huang, C.P. Responses of Algal Cells to Engineered Nanoparticles Measured as Algal Cell Population, Chlorophyll a, and Lipid Peroxidation: Effect of Particle Size and Type. J. Nanotechnol. 2012, 2012, 237284. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Liang, Z.; Zheng, X.; Zhao, W.; Wu, M.; Wang, Z. Toxicity of nano-TiO2 on algae and the site of reactive oxygen species production. Aquat. Toxicol. 2015, 158, 1–13. [Google Scholar] [CrossRef]
- Franqueira, D.; Orosa, M.; Torres, E.; Herrero, C.; Cid, A. Potential use of flow cytometry in toxicity studies with microalgae. Sci. Total. Environ. 2000, 247, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Eullaffroy, P.; Vernet, G. The F684/F735 chlorophyll fluorescence ratio: A potential tool for rapid detection and determination of herbicide phytotoxicity in algae. Water Res. 2003, 37, 1983–1990. [Google Scholar] [CrossRef]
- Hazeem, L.J.; Bououdina, M.; Rashdan, S.; Brunet, L.; Slomianny, C.; Boukherroub, R. Cumulative effect of zinc oxide and titanium oxide nanoparticles on growth and chlorophyll a content of Picochlorum sp. Environ. Sci. Pollut. Res. 2015, 23, 2821–2830. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of Nano-TiO2 on Photochemical Reaction of Chloroplasts of Spinach. Biol. Trace Element Res. 2005, 105, 269–280. [Google Scholar] [CrossRef]
- Lei, Z.; Mingyu, S.; Xiao, W.; Chao, L.; Chunxiang, Q.; Liang, C.; Hao, H.; Xiaoqing, L.; Fashui, H. Effects of Nano-anatase on Spectral Characteristics and Distribution of LHCII on the Thylakoid Membranes of Spinach. Biol. Trace Element Res. 2007, 120, 273–283. [Google Scholar] [CrossRef]
- Hartmann, N.B. Algal testing of titanium dioxide nanoparticles—Testing considerations, inhibitory effects and modification of cadmium bioavailability 8. Toxicology 2010, 269, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhou, L.; Liu, Y.; Deng, S.; Wu, H.; Wang, G. Toxicological effects of nanometer titanium dioxide (nano-TiO2) on Chlamydomonas reinhardtii. Ecotoxicol. Environ. Saf. 2012, 84, 155–162. [Google Scholar] [CrossRef]
- Gui, X.; Rui, M.; Song, Y.; Ma, Y.; Rui, Y.; Zhang, P.; He, X.; Li, Y.; Zhang, Z.; Liu, L. Phytotoxicity of CeO2 nanoparticles on radish plant (Raphanus sativus). Environ. Sci. Pollut. Res. 2017, 24, 13775–13781. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Ma, C.; Wang, K.; Hao, Y.; Chen, Q.; Mo, Y.; Rui, Y. Effects of cerium oxide on rice seed-lings as affected by co-exposure of cadmium and salt. Environ. Pollut. 2019, 252, 1087–1096. [Google Scholar] [CrossRef]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of Silver Nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef]
- Schwabe, F.; Schulin, R.; Limbach, L.K.; Stark, W.; Bürge, D.; Nowack, B. Influence of two types of organic matter on interaction of CeO2 nanoparticles with plants in hydroponic culture. Chemosphere 2013, 91, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Lukhele, L.P.; Mamba, B.; Musee, N.; Wepener, V. Acute Toxicity of Double-Walled Carbon Nanotubes to Three Aquatic Organisms. J. Nanomater. 2015, 2015, 219074. [Google Scholar] [CrossRef] [Green Version]
- Quigg, A.; Chin, W.C.; Chen, C.S.; Zhang, S.; Jiang, Y.; Miao, A.J.; Schwehr, K.A.; Xu, C.; Santschi, P.H. Direct and indirect toxic effects of engineered nanoparticles on algae: Role of natural organic matter. ACS Sustain. Chem. Eng. 2013, 1, 686–702. [Google Scholar] [CrossRef]
- Von Moos, N.; Slaveykova, V.I. Oxidative stress induced by inorganic nanoparticles in bacteria and aquatic microalgae—State of the art and knowledge gaps. Nanotoxicology 2013, 8, 605–630. [Google Scholar] [CrossRef]
- Chen, F.; Xiao, Z.; Yue, L.; Wang, J.; Feng, Y.; Zhu, X.; Wang, Z.; Xing, B. Algae response to engineered nano-particles: Current understanding, mechanisms and implications. Environ. Sci. Nano 2019, 6, 1026–1042. [Google Scholar] [CrossRef]
- Atienzar, F.A.; Jha, A.N. The random amplified polymorphic DNA (RAPD) assay and related techniques applied to genotoxicity and carcinogenesis studies: A critical review. Mutat. Res. Mol. Mech. Mutagen. 2006, 613, 76–102. [Google Scholar] [CrossRef]
- Załęska-Radziwiłł, M.; Doskocz, N. DNA changes in Pseudomonas putida induced by aluminum oxide nanoparticles using RAPD analysis. Desalination Water Treat. 2015, 57, 1573–1581. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, Y.; Yang, K.; Zhu, L.; Lin, D. Nanoparticle TiO2 size and rutile content impact bioconcentration and biomagnification from algae to daphnia. Environ. Pollut. 2019, 247, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Huang, G.; Zhang, B.; Zhou, Y. Trophic transfer potential of nTiO2, nZnO, and triclosan in an algae-algae eating fish food chain. Aquat. Toxicol. 2021, 235, 105824. [Google Scholar] [CrossRef] [PubMed]
- Matt, S.; Hofmann, T.G. The DNA damage-induced cell death response: A roadmap to kill cancer cells. Cell. Mol. Life Sci. 2016, 73, 2829–2850. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.-Q.; Cui, P.; Ru, S.; Kurade, M.B.; Patil, S.M.; Yadav, K.K.; Fallatah, A.M.; Cabral-Pinto, M.M.; Jeon, B.-H. A comprehensive review on the effects of engineered nanoparticles on microalgal treatment of pollutants from wastewater. J. Clean. Prod. 2022, 344, 131121. [Google Scholar] [CrossRef]
- Nyati, S.; Werth, S.; Honegger, R. Genetic Diversity of Sterile Cultured Trebouxia Photobionts Associated with the Lichen-Forming Fungus Xanthoria Parietina Visualized with RAPD-PCR Fingerprinting Techniques. The Lichenologist 2013, 344, 825–840. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahaye, N.; Musee, N. Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella subcapitata to Cerium Oxide Nanoparticles. Toxics 2023, 11, 283. https://doi.org/10.3390/toxics11030283
Mahaye N, Musee N. Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella subcapitata to Cerium Oxide Nanoparticles. Toxics. 2023; 11(3):283. https://doi.org/10.3390/toxics11030283
Chicago/Turabian StyleMahaye, Ntombikayise, and Ndeke Musee. 2023. "Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella subcapitata to Cerium Oxide Nanoparticles" Toxics 11, no. 3: 283. https://doi.org/10.3390/toxics11030283
APA StyleMahaye, N., & Musee, N. (2023). Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella subcapitata to Cerium Oxide Nanoparticles. Toxics, 11(3), 283. https://doi.org/10.3390/toxics11030283