
Rat Hepatocytes Protect against Lead–Cadmium-Triggered Apoptosis Based on Autophagy Activation

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Conditions for Cell Culture and Exposure
2.3. Cell Viability Assay
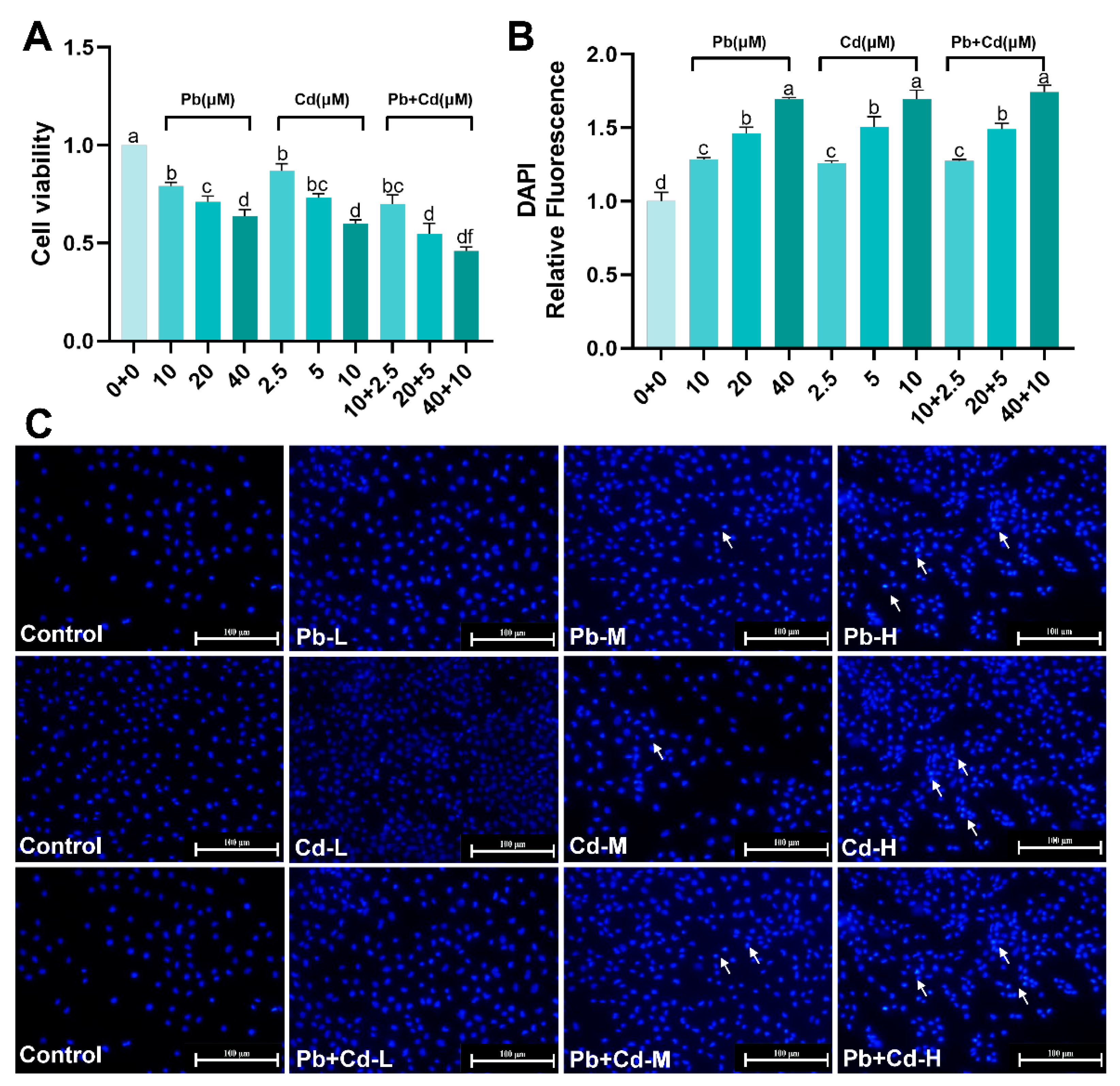
2.4. Nuclear Morphological Observations
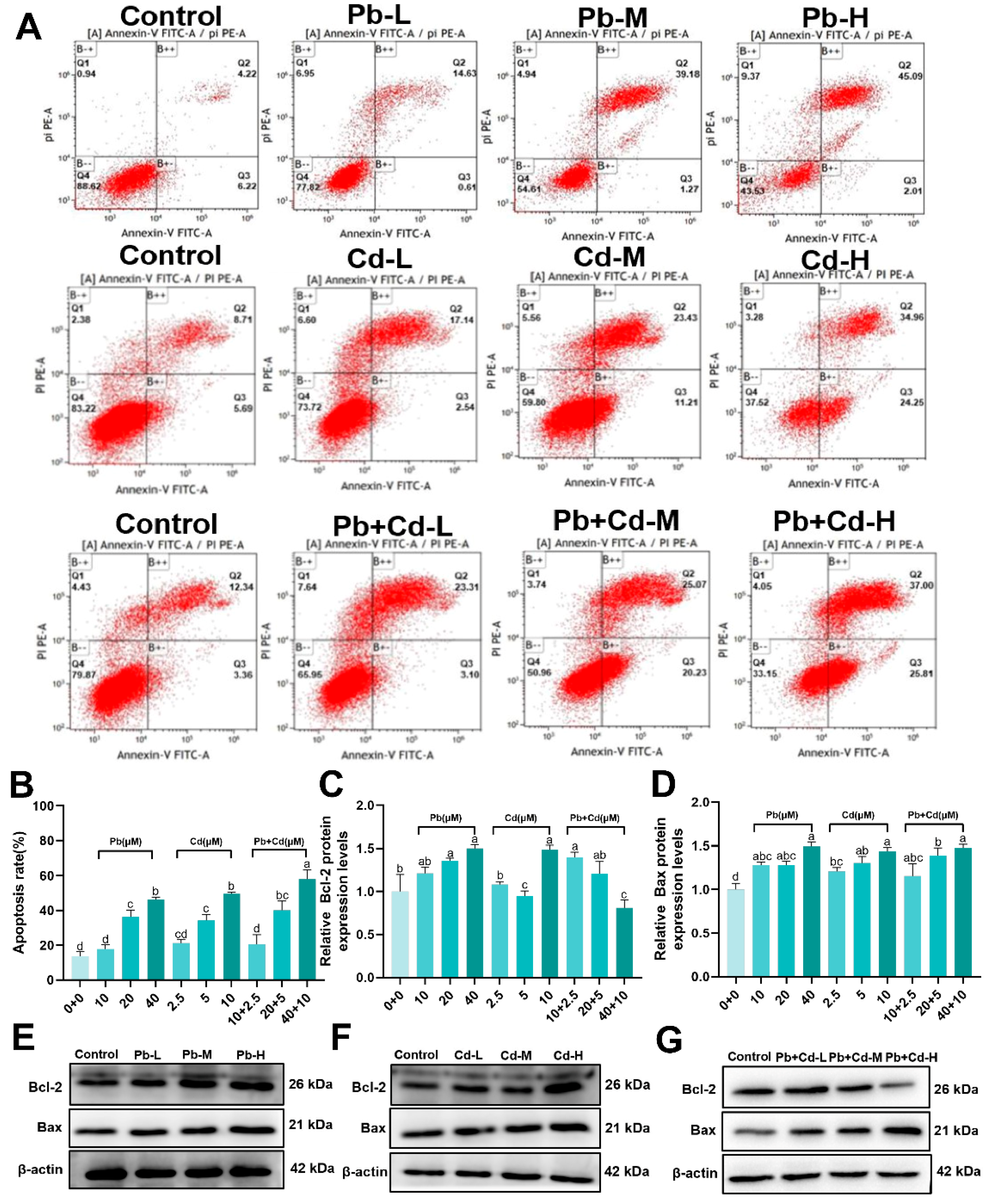
2.5. AnnexinV-FITC/PI Apoptosis Analysis
2.6. Detection of ROS Level
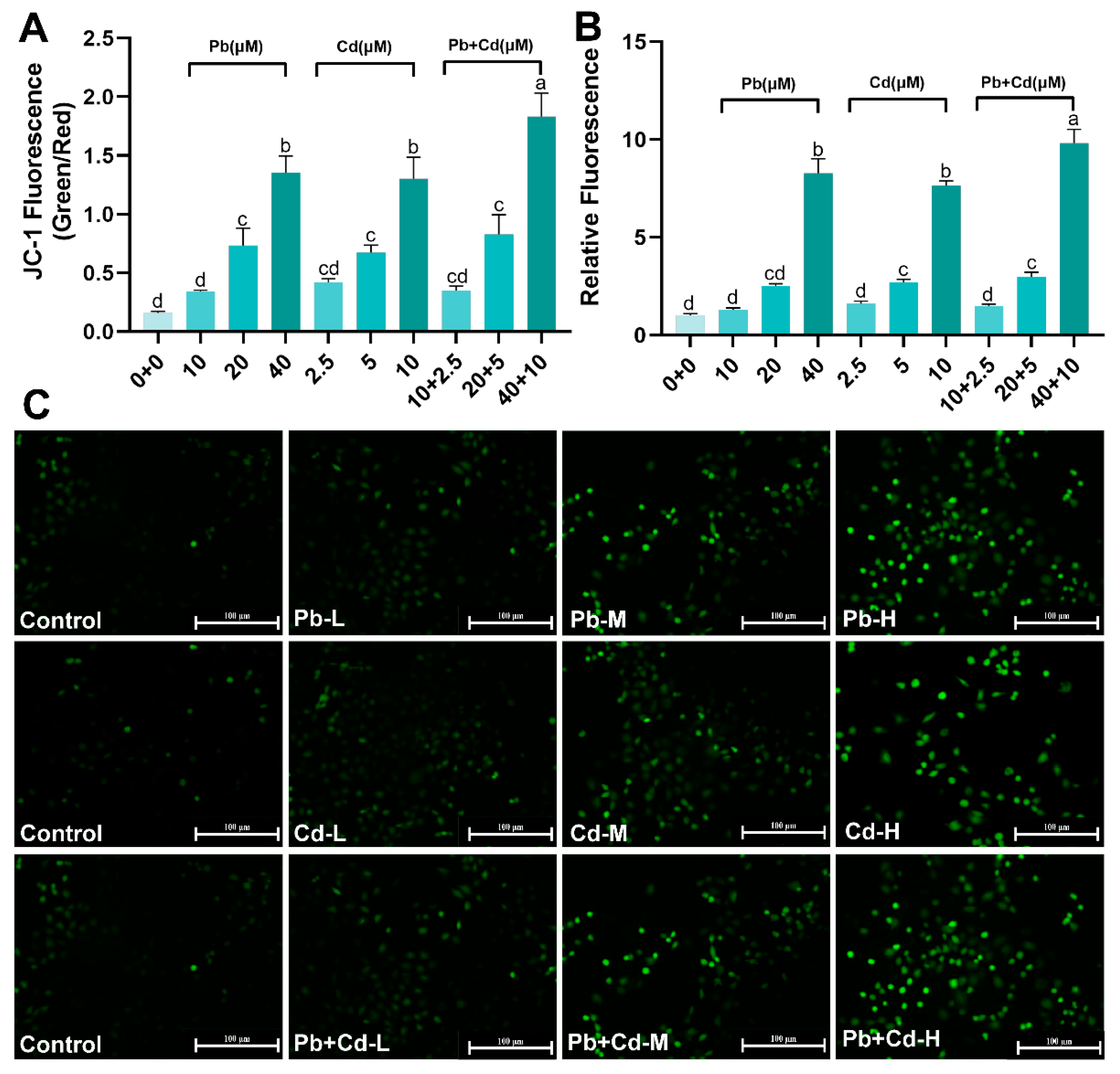
2.7. Mitochondrial Membrane Potential (MMP) Determination
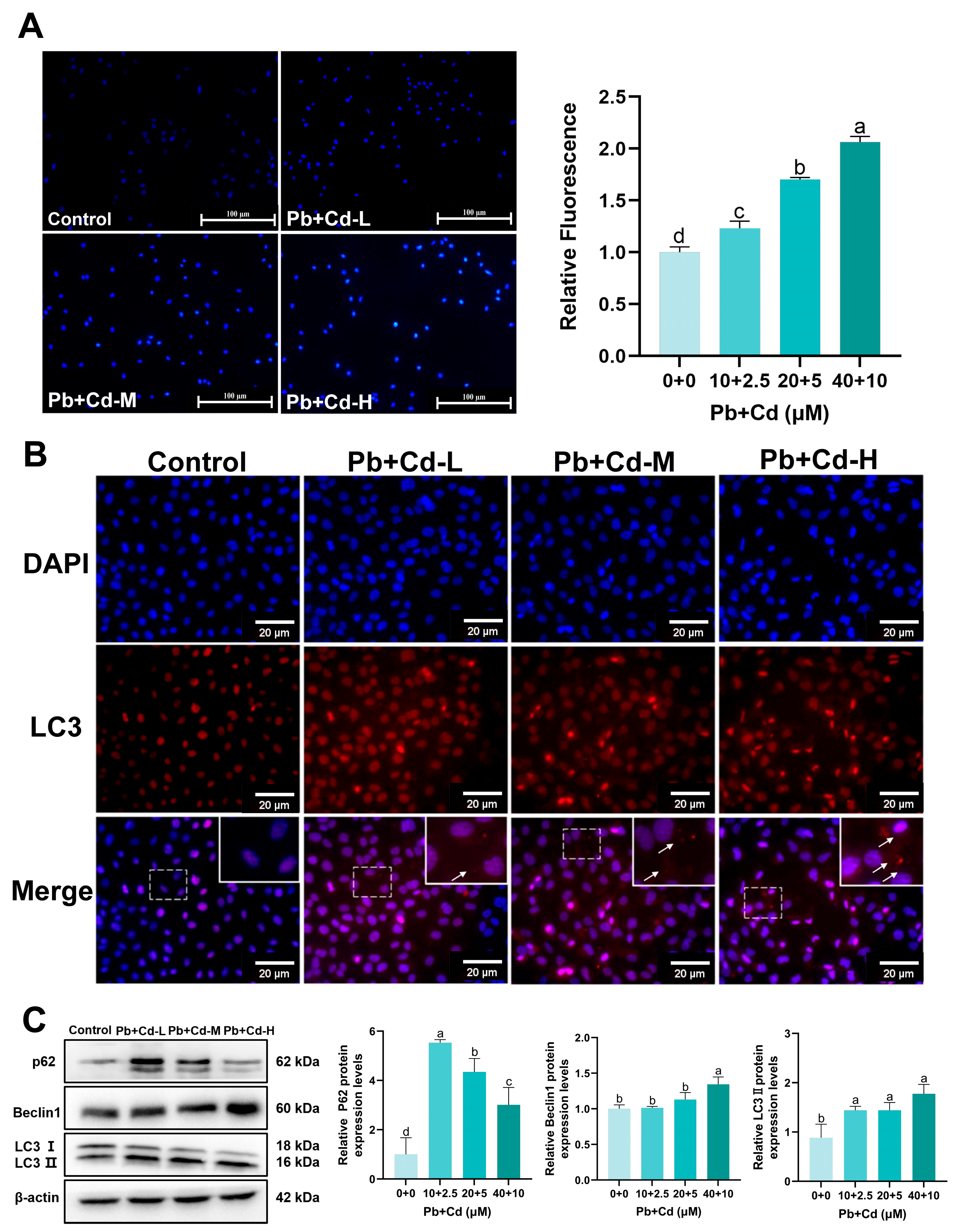
2.8. MDC-Stained Observation of Autophagy Vesicles
2.9. LC3 Immunofluorescence Aggregation Site Study
2.10. Transcriptomics
2.11. Real-Time Quantitative PCR Fluorescence Analysis
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
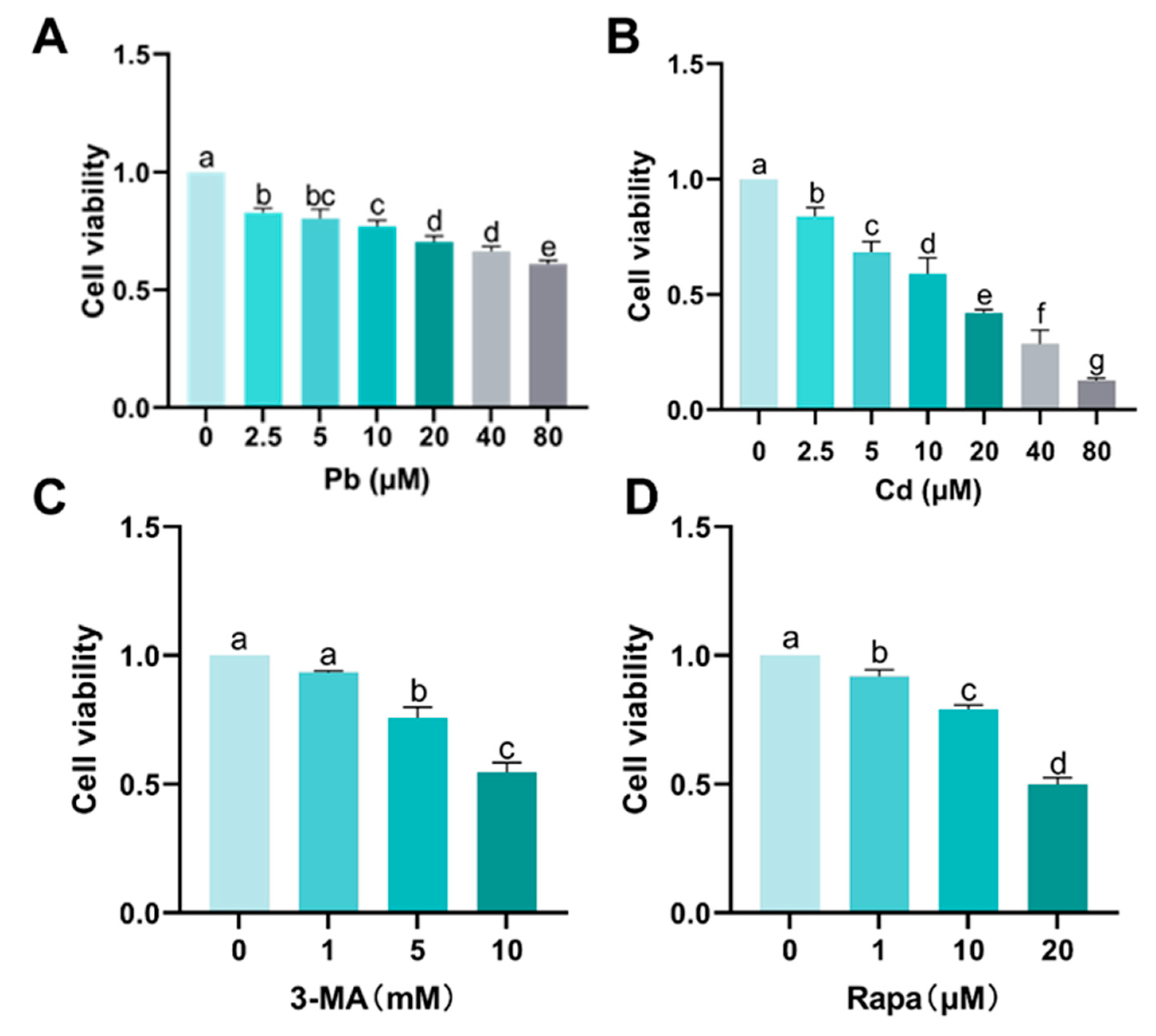
3.1. Cytotoxicity of Pb and Cd in BRL-3A Cells
3.2. Pb and Cd Cause ROS and MMP Changes in BRL-3A Cells
3.3. Pb and Cd Induce Apoptosis in BRL-3A Cells
3.4. Pb and Cd Trigger Autophagy in BRL-3A Cells
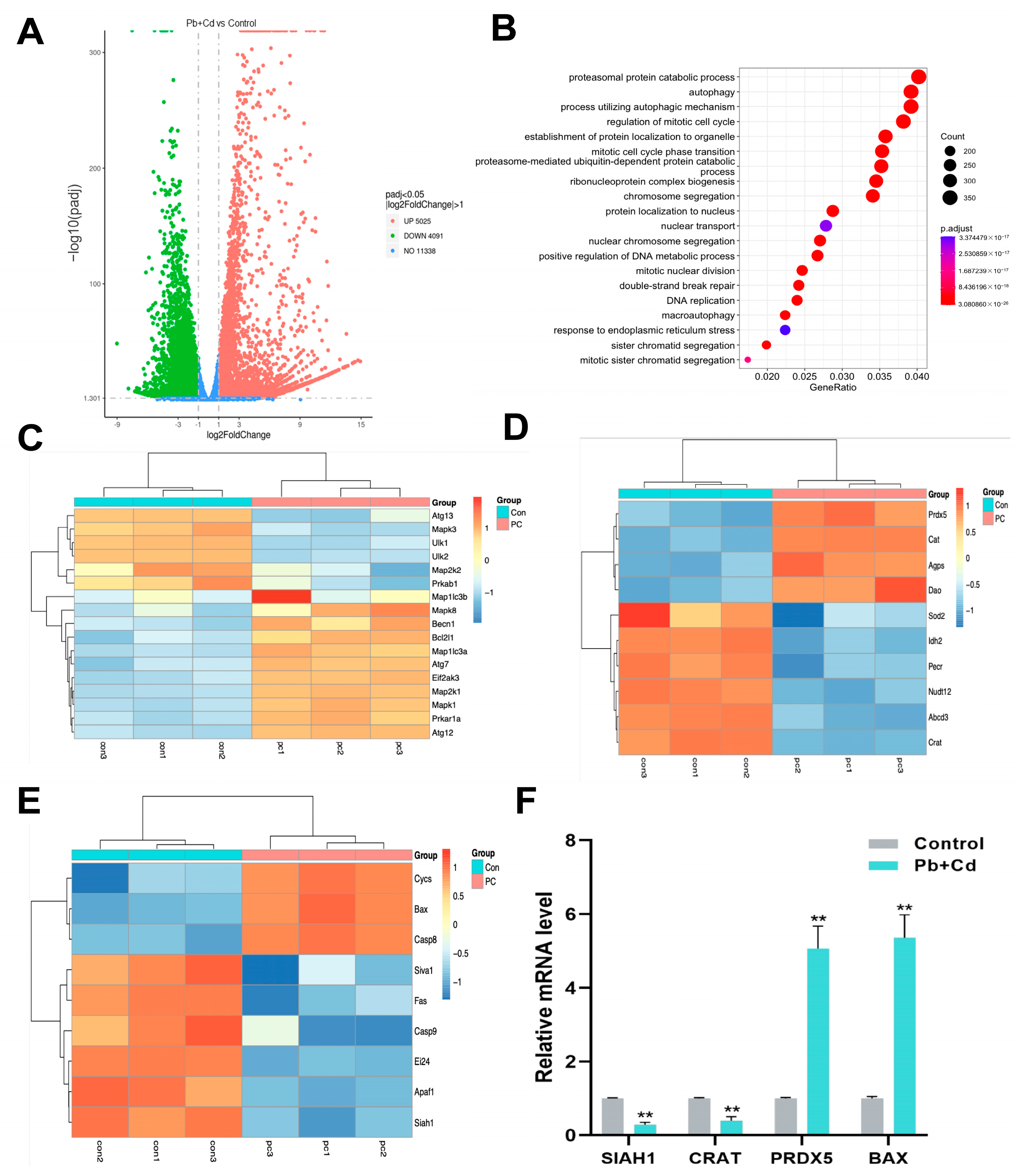
3.5. GO Enrichment Analysis and Transcriptomic Validation of DEGs
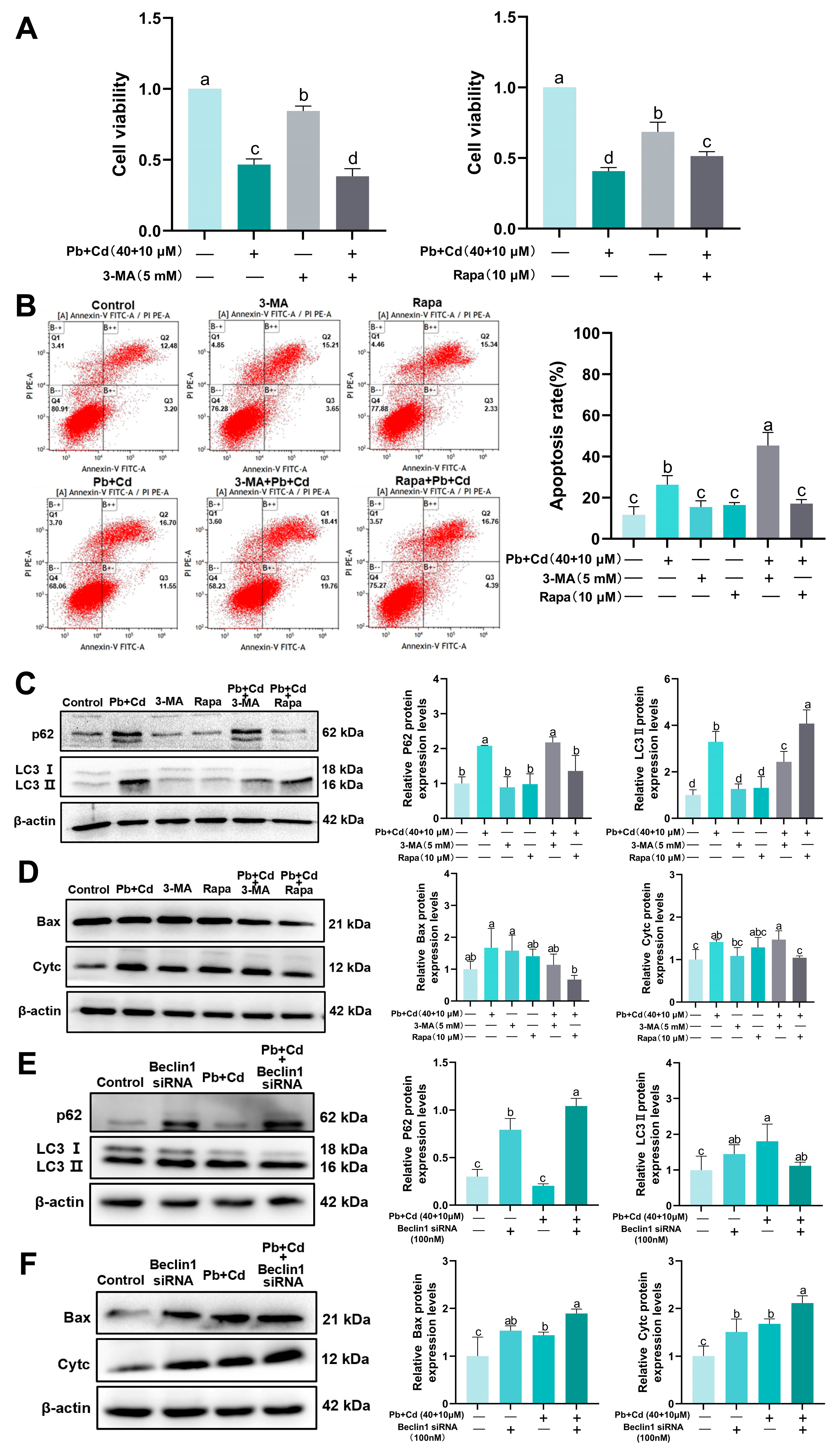
3.6. Autophagy Plays a Protective Role in BRL-3A Cells Exposed to Pb and Cd
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Screening for Pb, Cd, 3-MA, and Rapa Concentrations

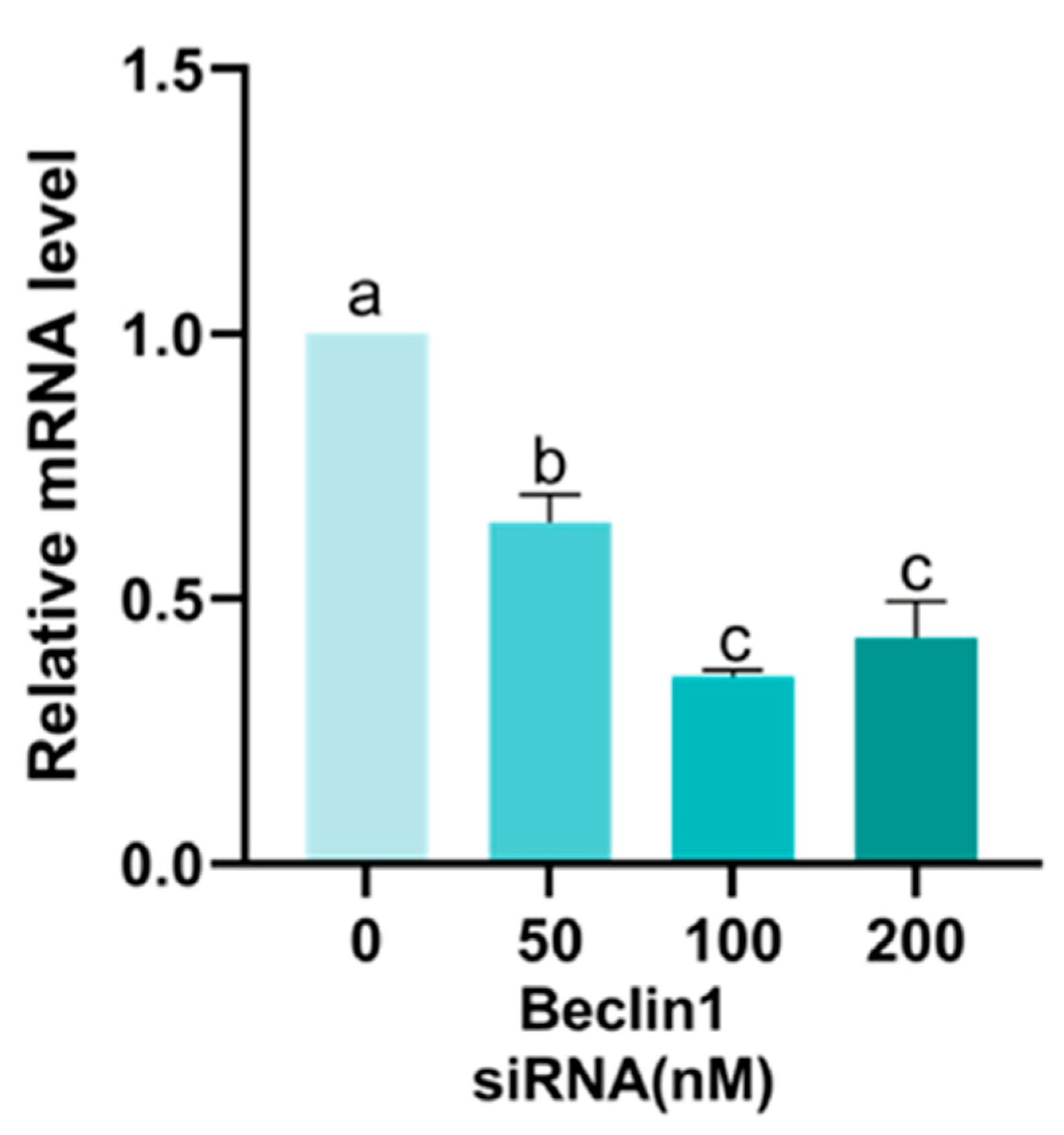
Appendix A.2. Screening of Beclin1 siRNA Concentration

References
- Guo, Y.; Yang, Y.; Li, R.; Liao, X.; Li, Y. Distribution of cadmium and Pb in soil-rice systems and their environmental driving factors at the island scale. Ecotoxicol. Environ. Saf. 2023, 265, 115530. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Jiang, S.; Yan, X.; Qin, Z.; Jia, C.; Li, Z.; Zhang, J.; Huang, R. The mobility of cadmium and Pb in the soil-mulberry-silkworm system. Chemosphere 2020, 242, 125179. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Liu, H.; Banet, T.; Wang, H.; Ippolito, J.A.; Li, L. Cadmium, copper, Pb and zinc accumulation in wild plant species near a Pb smelter. Ecotoxicol. Environ. Saf. 2020, 198, 110683. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kumar, A.; Gupta, V.K.; Sharma, B. Biochemical and Molecular Bases of Pb-Induced Toxicity in Mammalian Systems and Possible Mitigations. Chem. Res. Toxicol. 2018, 31, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Jing, D.; Huang, X.; Xiao, Y.; Shu, Z.; Luo, D.; Duan, Y.; He, M.; Xiao, S.; Chen, X. Effects of co-exposure to multiple metals on children′s behavior problems in China. Sci. Total Environ. 2022, 826, 154062. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, R.; Bhattacharya, R.; Chatterjee, M. Biochemical, haematological and histopathological study in relation to time-related cadmium-induced hepatotoxicity in mice. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2000, 13, 231. [Google Scholar]
- Renu, K.; Chakraborty, R.; Haritha, M.; Rajeshwari, K.; Abilash, V.G. Molecular mechanism of heavy metals (Pb, Chromium, Arsenic, Mercury, Nickel and Cadmium) induced hepatotoxicity—A review. Chemosphere 2021, 271, 129735. [Google Scholar] [CrossRef]
- Ma, L.; Liu, J.; Dong, J.; Xiao, Q.; Zhao, J.; Jiang, F. Toxicity of Pb2+ on rat liver mitochondria induced by oxidative stress and mitochondrial permeability transition. Toxicol. Res. 2017, 6, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Matović, V.; Buha, A.; Ðukić-Ćosić, D.; Bulat, Z. Insight into the oxidative stress induced by Pb and/or cadmium in blood, liver and kidneys. Food Chem. Toxicol. 2015, 78, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Pandya, C.D.; Pillai, P.P.; Gupta, S.S. Pb and Cadmium Co-exposure Mediated Toxic Insults on Hepatic Steroid Metabolism and Antioxidant System of Adult Male Rats. Biol. Trace Elem. Res. 2010, 134, 307. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Feng, X.; Shen, J.; Wang, Y.; Zhang, Y.; Zhou, R. Pb intoxication-induced exosomes promote autophagy and apoptosis in renal proximal tubule cells by activating the adenosine 5′-monophosphate-activated protein kinase signaling. Environ. Toxicol. 2023, 38, 1011–1021. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Jin, X.; Fan, R.; Xing, M.; Guo, J.; Zhang, Z.; Zhang, J.; Xu, S. Cadmium-mediated miR-30a-GRP78 Pbs to JNK-dependent autophagy in chicken kidney. Chemosphere 2019, 215, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, J.; Yang, J.; Cai, J.; Liu, Q.; Zhang, X.; Bao, J.; Zhang, Z. Cadmium induces apoptosis and autophagy in swine small intestine by downregulating the PI3K/Akt pathway. Environ. Sci. Pollut. Res. Int. 2022, 29, 41207–41218. [Google Scholar] [CrossRef] [PubMed]
- Vaux, D.L. Apoptosis and toxicology—What relevance? Toxicology 2002, 181–182, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Ma, T.; Lv, L.; Yu, Y.; Zhao, M.; Chen, H.; Gao, L. Endoplasmic reticulum stress mediated by ROS participates in cadmium exposure-induced MC3T3-E1 cell apoptosis. Ecotoxicol. Environ. Saf. 2023, 251, 114517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liao, J.; Hu, Z.; Li, Q.; Hu, L.; Guo, J.; Li, Y.; Zhang, H.; Pan, J.; Tang, Z. Mitochondria-mediated apoptosis and endoplasmic reticulum stress are involved in the toxicity induced by copper in the porcine spleen. Environ. Sci. Pollut. Res. Int. 2023, 30, 94928–94939. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wang, T.; Pan, T.T.; Huang, M.; Ren, W.H.; Xu, G.L.; Amin, H.K.; Kassab, R.B.; Moneim, A.E.A. Senna alexandrina extract supplementation reverses hepatic oxidative, inflammatory, and apoptotic effects of cadmium chloride administration in rats. Environ. Sci. Pollut. Res. 2020, 27, 5981–5992. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; He, Y.; Wang, H.; Zhang, Q. Protective effect of melatonin against chronic cadmium-induced hepatotoxicity by suppressing oxidative stress, inflammation, and apoptosis in mice. Ecotoxicol. Environ. Saf. 2021, 228, 112947. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Autophagy in the Pathogenesis of Disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, Y.; An, Y.; Jiao, W.; Xu, Y.; Han, Q.; Teng, X.; Teng, X. Selenium alleviates oxidative stress and autophagy in Pb-treated chicken testes. Theriogenology 2019, 131, 146–152. [Google Scholar] [CrossRef]
- Zou, H.; Sun, J.; Wu, B.; Yuan, Y.; Gu, J.; Bian, J.; Liu, X.; Liu, Z. Effects of Cadmium and/or Pb on Autophagy and Liver Injury in Rats. Biol. Trace Elem. Res. 2020, 198, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.; Lin, M.; Qi, Q.; Moore, J.T.; Levine, K.E.; Ferno, R.A.; Kumar, D. Role of Autophagy in Cadmium-Induced Hepatotoxicity and Liver Diseases. J. Toxicol. 2021, 2021, 9564297. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Zhuo, L.; Han, T.; Hu, D.; Yang, X.; Wang, Y.; Yuan, Y.; Gu, J.; Bian, J.; Liu, X.; et al. Autophagy and gap junctional intercellular communication inhibition are involved in cadmium-induced apoptosis in rat liver cells. Biochem. Biophys. Res. Commun. 2015, 459, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Lamark, T.; Sjottem, E.; Bowitz Larsen, K.; Atesoh Awuh, J.; Overvatn, A.; Mcmahon, M.; Hayes, J.D.; Johansen, T. p62/SQSTM1 Is a Target Gene for Transcription Factor NRF2 and Creates a Positive Feedback Loop by Inducing Antioxidant Response Element-driven Gene Transcription. J. Biol. Chem. 2010, 285, 22576–22591. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, S.; Ma, B.; Wang, J.; Chen, J. Di-isononyl phthalate induces apoptosis and autophagy of mouse ovarian granulosa cells via oxidative stress. Ecotoxicol. Environ. Saf. 2022, 242, 113898. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wen, X.; Lin, P.; Chen, H.; Wang, A.; Jin, Y. HERP depletion inhibits zearalenone-induced apoptosis through autophagy activation in mouse ovarian granulosa cells. Toxicol. Lett. 2018, 301, 1–10. [Google Scholar] [CrossRef]
- Yuan, G.; Dai, S.; Yin, Z.; Lu, H.; Zhang, M. Sub-chronic Pb and cadmium co-induce apoptosis protein expression in liver and kidney of rats. Int. J. Clin. Exp. Pathol. 2014, 7, 2905. [Google Scholar] [PubMed]
- Hu, D.; Zou, H.; Han, T.; Xie, J.Z.; Dai, N.N.; Zhuo, L.L.; Gu, J.H.; Bian, J.C.; Yuan, Y.; Liu, X.Z.; et al. Gap junction blockage promotes cadmium-induced apoptosis in BRL 3A derived from Buffalo rat liver cells. J. Vet. Sci. 2016, 17, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.D.; Fergusson, J.E. The concentrations, distribution and sources of cadmium, copper, Pb and zinc in the atmosphere of an urban environment. Sci. Total Environ. 1994, 144, 179. [Google Scholar] [CrossRef]
- Li, C.; Shi, L.; Peng, C.; Yu, G.; Zhang, Y.; Du, Z. Pb-induced cardiomyocytes apoptosis by inhibiting gap junction intercellular communication via autophagy activation. Chem. Biol. Interact. 2021, 337, 109331. [Google Scholar] [CrossRef] [PubMed]
- Yin, H. Research on the Role and Mechanism of Autophagy in NiCl2-Induced Apoptosis in Kidney. Ph.D. thesis, Sichuan Agricultural 481 University, Chengdu, China, 2022. [Google Scholar]
- Wang, C.; Nie, G.; Zhuang, Y.; Hu, R.; Zhang, C. Inhibition of autophagy enhances cadmium-induced apoptosis in duck renal tubular epithelial cells. Ecotoxicol. Environ. Saf. 2020, 205, 111188. [Google Scholar] [CrossRef] [PubMed]
- Siraj, M.; Khisroon, M.; Khan, A.; Zaidi, F.; Ullah, A.; Rahman, G. Bio-monitoring of Tissue Accumulation and Genotoxic Effect of Heavy Metals in Cyprinus carpio from River Kabul Khyber Pakhtunkhwa Pakistan. Bull. Environ. Contam. Toxicol. 2018, 100, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Souza-Arroyo, V.; Fabián, J.J.; Bucio-Ortiz, L.; Miranda-Labra, R.U.; Gomez-Quiroz, L.E.; Gutiérrez-Ruiz, M.C. The mechanism of the cadmium-induced toxicity and cellular response in the liver. Toxicology 2022, 480, 153339. [Google Scholar] [CrossRef] [PubMed]
- Chi, Q.; Liu, T.; Sun, Z.; Tan, S.; Li, S.; Li, S. Involvement of mitochondrial pathway in environmental metal pollutant Pb-induced apoptosis of chicken liver: Perspectives from oxidative stress and energy metabolism. Environ. Sci. Pollut. Res. 2017, 24, 28121–28131. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Cheng, J.; Bao, L.; Zhu, X.; Huang, H. Exposure to waterborne cadmium induce oxidative stress, autophagy and mitochondrial dysfunction in the liver of Procypris merus. Ecotoxicol. Environ. Saf. 2020, 204, 111051. [Google Scholar] [CrossRef] [PubMed]
- Palikaras, K.; Tavernarakis, N. Mitochondrial homeostasis: The interplay between mitophagy and mitochondrial biogenesis. Exp. Gerontol. 2014, 56, 182–188. [Google Scholar] [CrossRef]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Liu, J. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2021, 263, 128346. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Muneer, P.M.; Chandra, N.; Haorah, J. Interactions of Oxidative Stress and Neurovascular Inflammation in the Pathogenesis of Traumatic Brain Injury. Mol. Neurobiol. 2015, 51, 966. [Google Scholar] [CrossRef] [PubMed]
- Mu, M.; Zhao, H.; Wang, Y.; Liu, J.; Fei, D.; Xing, M. Arsenic trioxide or/and copper sulfate co-exposure induce glandular stomach of chicken injury via destruction of the mitochondrial dynamics and activation of apoptosis as well as autophagy. Ecotoxicol. Environ. Saf. 2019, 185, 109678. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Uson-Lopez, R.A.; Sikder, M.T.; Tan, G.; Hosokawa, T.; Saito, T.; Kurasaki, M. Ameliorative effects of selenium on arsenic-induced cytotoxicity in PC12 cells via modulating autophagy/apoptosis. Chemosphere 2018, 196, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.D.; Manon, S.; Gonçalves, J.; Saraiva, L.; Côrte-Real, M. The importance of humanized yeast to better understand the role of bcl-2 family in apoptosis: Finding of novel therapeutic opportunities. Curr. Pharm. Des. 2011, 17, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Yin, Z.; Yuan, G.; Lu, H.; Jia, R.; Xu, J.; Song, X.; Li, L.; Shu, Y.; Liang, X.; et al. Quantification of metallothionein on the liver and kidney of rats by subchronic Pb and cadmium in combination. Environ. Toxicol. Pharmacol. 2013, 36, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R. Mechanisms of p53-induced apoptosis. Biochem. Pharmacol. 1999, 58, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of Cells and Tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Dai, N.; Wang, Y.; Xu, C.; Liu, Z. Role of autophagy in cadmium-induced apoptosis of primary rat osteoblasts. Sci. Rep. 2016, 6, 20404. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Ma, S.X.; Qi, Y.M.; Wei, X.; Cai, H.; Dong, L.; Lu, Y.F.; Zhang, Y.P.; Guo, Q.J. Quercetin inhibited cadmium-induced autophagy in the mouse kidney via inhibition of oxidative stress. J. Toxicol. Pathol. 2016, 29, 247–252. [Google Scholar] [CrossRef]
- Chu, B.-X.; Fan, R.-F.; Lin, S.-Q.; Yang, D.-B.; Wang, Z.-Y.; Wang, L. Interplay between autophagy and apoptosis in Pb(II)-induced cytotoxicity of primary rat proximal tubular cells. J. Inorg. Biochem. Interdiscip. J. 2018, 182, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Pu, W.J.; Chu, X.S.; Guo, H.L.; Huang, G.; Cui, T.; Huang, B.Y.; Dai, X.Y.; Zhang, C.Y. The activated ATM/AMPK/mTOR axis promotes autophagy in response to oxidative stress-mediated DNA damage co-induced by molybdenum and cadmium in duck testes. Environ. Pollut. 2023, 316, 11. [Google Scholar] [CrossRef]
- Lv, X.H.; Zhao, D.H.; Cai, S.Z.; Luo, S.Y.; You, T.; Xu, B.L.; Chen, K. Autophagy plays a protective role in cell death of osteoblasts exposure to Pb chloride. Toxicol. Lett. 2015, 239, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Xin-Yu, W.; Heng, Y.; Min-Ge, W.; Du-Bao, Y.; Zhen-Yong, W.; Lin, W. Trehalose protects against cadmium-induced cytotoxicity in primary rat proximal tubular cells via inhibiting apoptosis and restoring autophagic flux. Cell Death Dis. 2017, 8, e3099. [Google Scholar]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.X.; Zhu, H.L.; Shi, X.T.; Nan, Y.; Wang, H. Autophagy in Sertoli cell protects against environmental cadmium-induced germ cell apoptosis in mouse testes. Environ. Pollut. 2021, 270, 116241. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Hu, D.; Han, T.; Zhao, H.; Xie, J.; Liu, X.; Wang, Y.; Gu, J.; Yuan, Y.; Bian, J.; et al. Salidroside ameliorates Cd-induced calcium overload and gap junction dysfunction in BRL 3A rat liver cells. Biol. Trace Elem. Res. 2015, 164, 90–98. [Google Scholar] [CrossRef]
- Guo, H.; Ouyang, Y.; Yin, H.; Cui, H.; Deng, H.; Liu, H.; Jian, Z.; Fang, J.; Zuo, Z.; Wang, X.; et al. Induction of autophagy via the ROS-dependent AMPK-mTOR pathway protects copper-induced spermatogenesis disorder. Redox Biol. 2022, 49, 102227. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zuo, Z.; Zeng, Y.; Ouyang, Y.; Cui, H.; Deng, H.; Zhu, Y.; Deng, J.; Geng, Y.; Ouyang, P.; et al. Autophagy-mediated ferroptosis involved in nickel-induced nephrotoxicity in the mice. Ecotoxicol. Environ. Saf. 2023, 259, 115049. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ to 3′) |
|---|---|
| Beclin1 | Forward: AGGAGTTGCCGTTGTACTGTTCTG Reverse: TGCCTCCAGTGTCTTCAATCTTGC |
| BAX | Forward: CCAGGACGCATCCACCAAGAAGC Reverse: TGCCACACGGAAGAAGACCTCTCG |
| CRAT | Forward: CAAGCAGGACTTCATGGATCTACAG Reverse: GGCAGCGTCTCGTTGTCAATC |
| PRDX5 | Forward: CCACCAGGCAGAAGGCAAGG Reverse: CGATTCCCAAAGAGAGACACCAAAG |
| SIAH1 | Forward: CAAAGTGTCCACCATCCCAGAG Reverse: GGTGGCAATACATAGTCAAAGCAG |
| β-actin | Forward: CTAAGGCCAACCGTGAAAAG Reverse: AACACAGCCTGGATGGCTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, J.; Liu, H.; Yin, T.; Zhou, X.; Song, X.; Zou, Y.; Li, L.; Jia, R.; Fu, Y.; Zhao, X.; et al. Rat Hepatocytes Protect against Lead–Cadmium-Triggered Apoptosis Based on Autophagy Activation. Toxics 2024, 12, 285. https://doi.org/10.3390/toxics12040285
Xue J, Liu H, Yin T, Zhou X, Song X, Zou Y, Li L, Jia R, Fu Y, Zhao X, et al. Rat Hepatocytes Protect against Lead–Cadmium-Triggered Apoptosis Based on Autophagy Activation. Toxics. 2024; 12(4):285. https://doi.org/10.3390/toxics12040285
Chicago/Turabian StyleXue, Junshu, Huimao Liu, Tianyi Yin, Xun Zhou, Xu Song, Yuanfeng Zou, Lixia Li, Renyong Jia, Yuping Fu, Xinghong Zhao, and et al. 2024. "Rat Hepatocytes Protect against Lead–Cadmium-Triggered Apoptosis Based on Autophagy Activation" Toxics 12, no. 4: 285. https://doi.org/10.3390/toxics12040285
APA StyleXue, J., Liu, H., Yin, T., Zhou, X., Song, X., Zou, Y., Li, L., Jia, R., Fu, Y., Zhao, X., & Yin, Z. (2024). Rat Hepatocytes Protect against Lead–Cadmium-Triggered Apoptosis Based on Autophagy Activation. Toxics, 12(4), 285. https://doi.org/10.3390/toxics12040285