Cytotoxicity and Toxicity Evaluation of Xanthone Crude Extract on Hypoxic Human Hepatocellular Carcinoma and Zebrafish (Danio rerio) Embryos

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Xanthone Extract
2.2. High Performance Liquid Chromatography (HPLC) Profiling of XCE
2.3. Cytotoxicity Assay
2.4. Zebrafish Embryos
2.5. Zebrafish Embryonic Toxicity Test
3. Results and Discussion
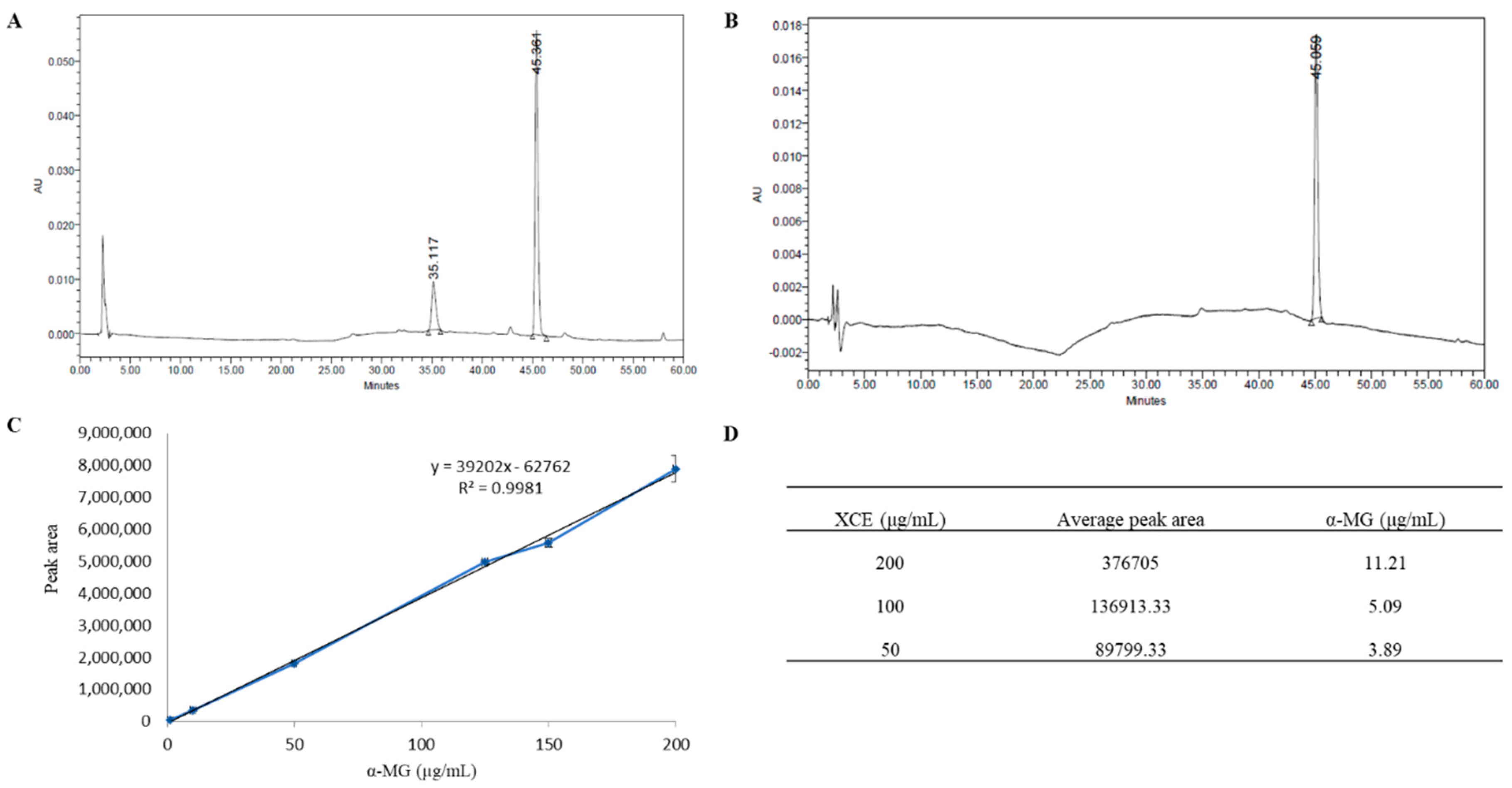
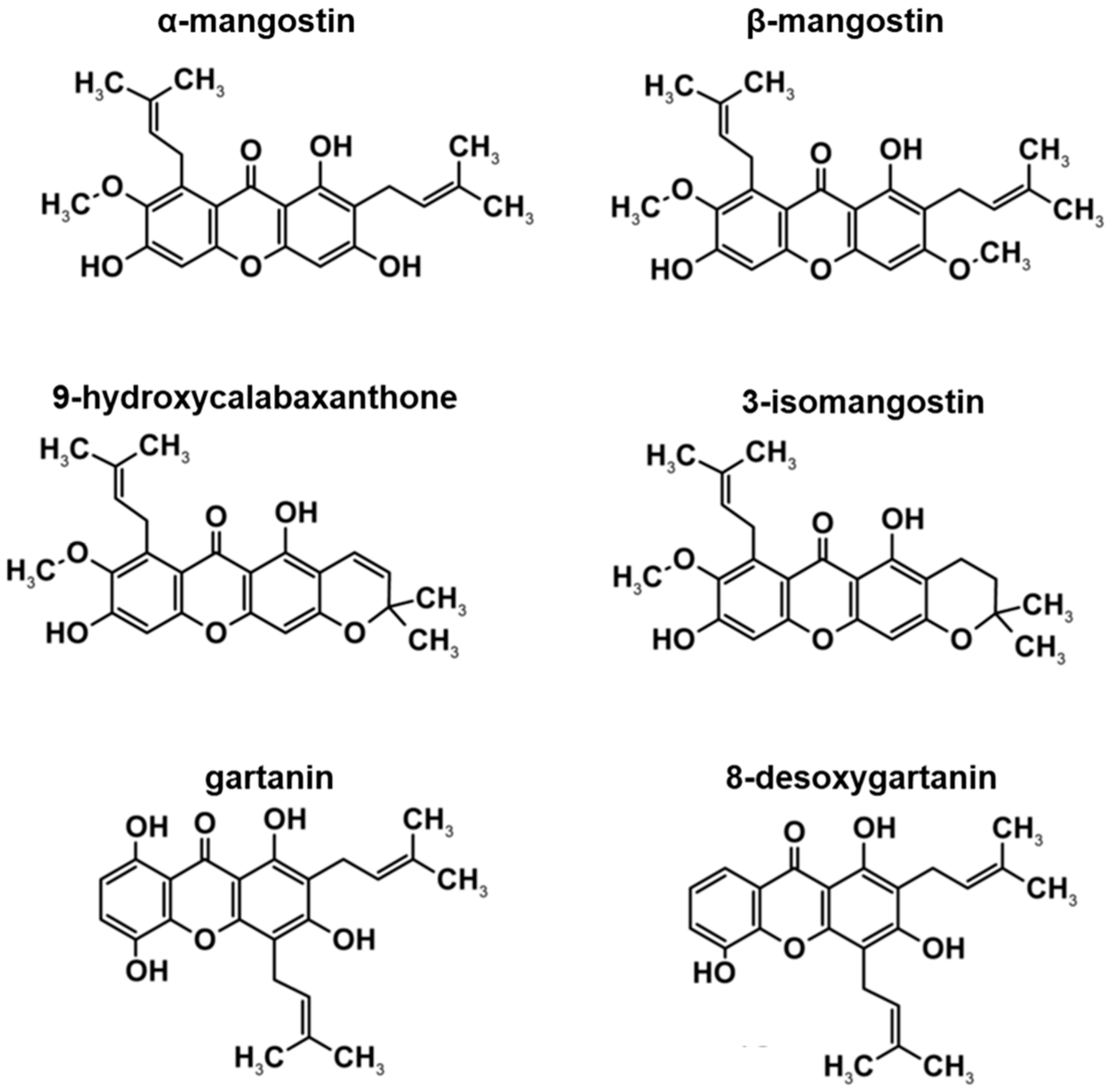
3.1. HPLC Profile of Xanthone Crude Extract
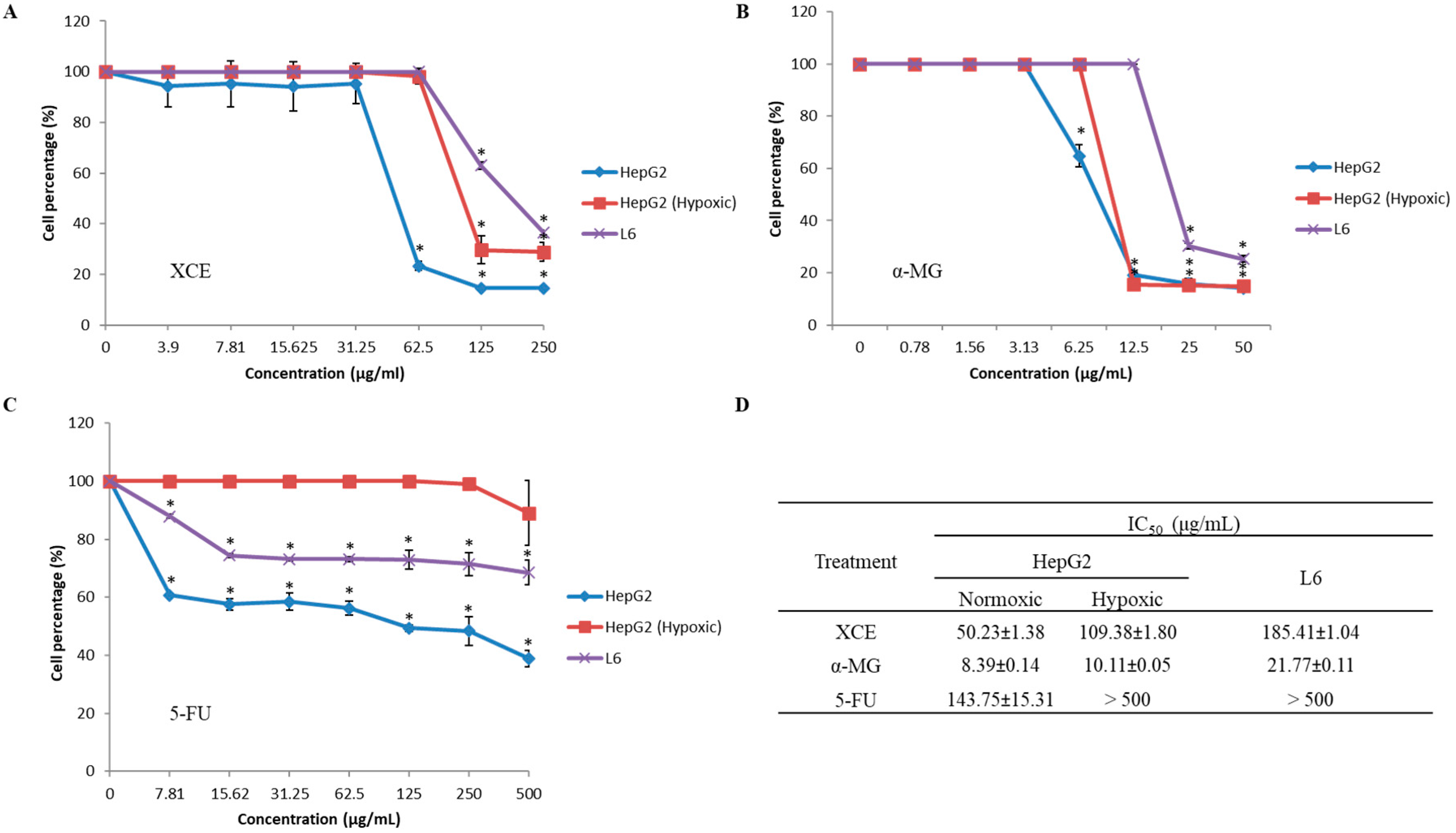
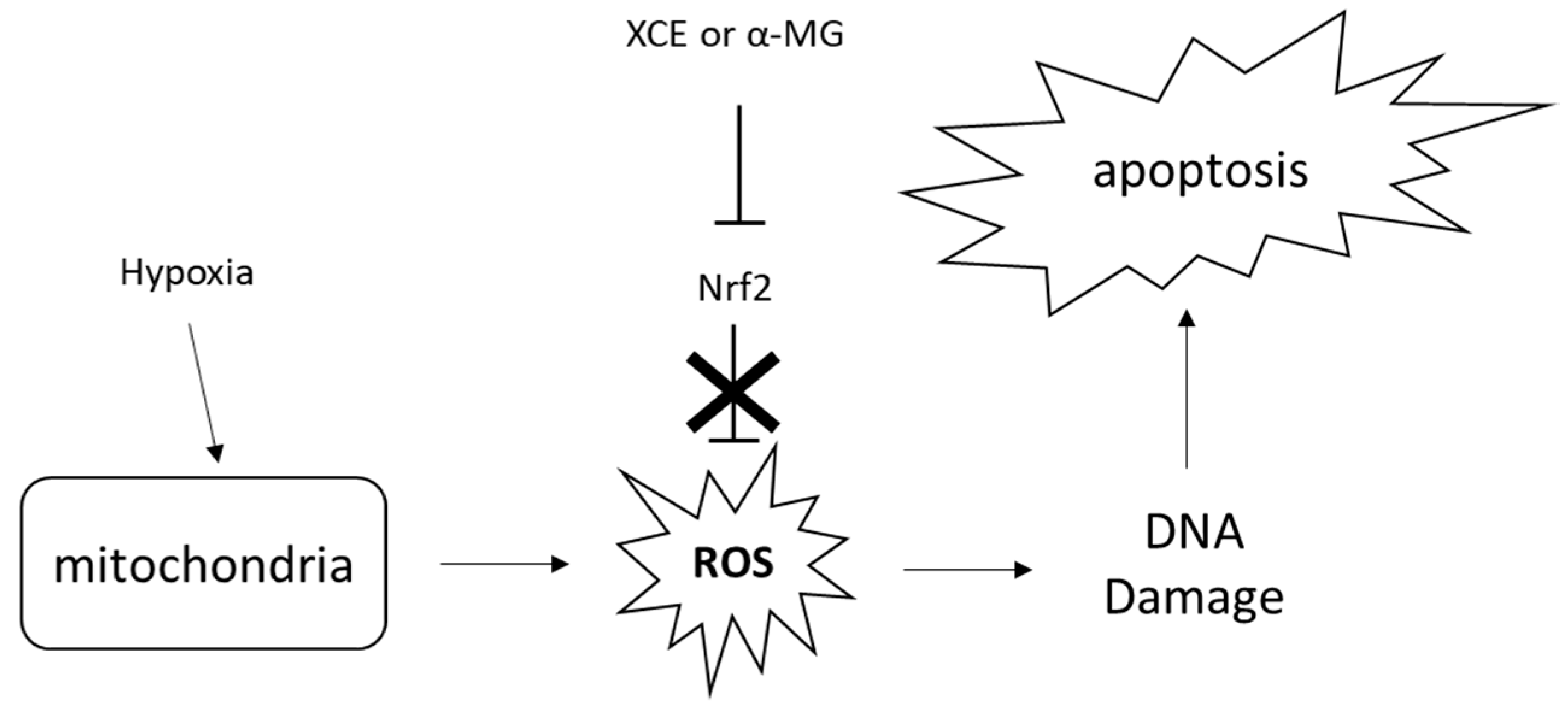
3.2. Cytotoxicity of XCE and α-MG
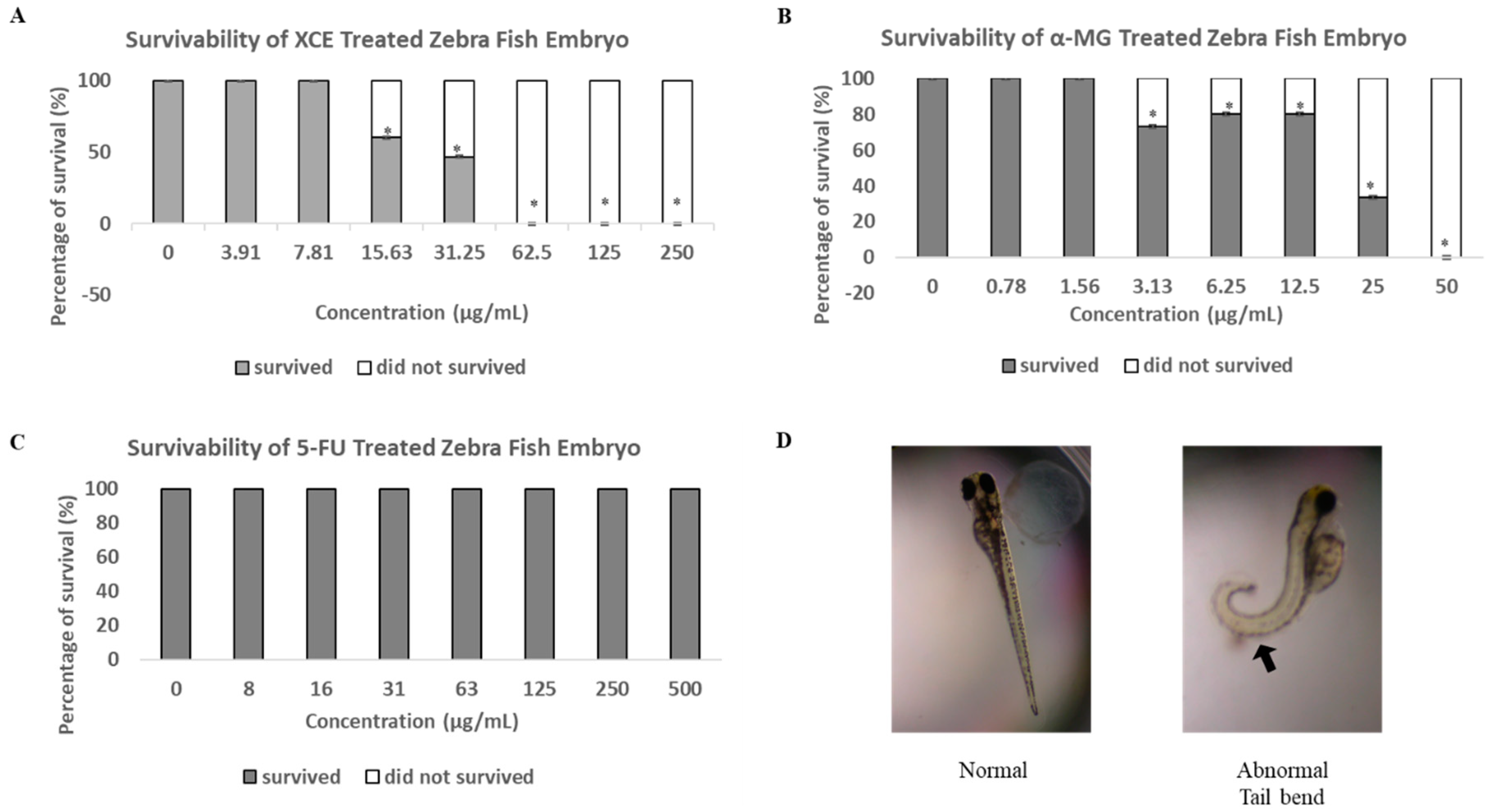
3.3. Toxicity of XCE and α-MG on Zebrafish Embryos
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sia, D.; Villanueva, A.; Friedman, S.L.; Llovet, J.M. Liver cancer cell of origin, molecular class, and effects on patient prognosis. Gastroenterology 2017, 152, 745–761. [Google Scholar] [CrossRef] [PubMed]
- Goh, K.L.; Razlan, H.; Hartono, J.L.; Qua, C.S.; Yoong, B.K.; Koh, P.S.; Abdullah, B.J.J. Liver cancer in Malaysia: Epidemiology and clinical presentation in a multiracial Asian population. J. Digest. Dis. 2015, 16, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Raihan, R.; Azzeri, A.; Shabaruddin, F.H.; Mohamed, R. Hepatocellular Carcinoma in Malaysia and Its Changing Trend. Euroasian J. Hepato-Gastroenterol. 2018, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Leung, T.W.; Tang, A.M.; Zee, B.; Yu, S.C.; Lai, P.B.; Lau, W.Y.; Johnson, P.J. Factors predicting response and survival in 149 patients with unresectable hepatocellular carcinoma treated by combination cisplatin, interferon-α, doxorubicin and 5-fluorouracil chemotherapy. Cancer 2002, 94, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Okusaka, T.; Ueno, H.; Takezako, Y.; Morizane, C. A phase II trial of continuous infusion of 5-fluorouracil, mitoxantrone, and cisplatin for metastatic hepatocellular carcinoma. Cancer 2005, 103, 756–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Ahn, S.H.; Yoon, Y.J.; Kim, J.K.; Lee, H.W.; Lee, D.Y.; Chon, C.Y.; Moon, Y.M.; Han, K.H. Repetitive short-course hepatic arterial infusion chemotherapy with high-dose 5-fluorouracil and cisplatin in patients with advanced hepatocellular carcinoma. Cancer 2007, 110, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, C.K.; Yang, Z.F.; Ho, D.W.; Ng, M.N.; Yeoh, G.C.; Poon, R.T.; Fan, S.T. An Akt/hypoxia-inducible factor-1α/platelet-derived growth factor-BB autocrine loop mediates hypoxia-induced chemoresistance in liver cancer cells and tumorigenic hepatic progenitor cells. Clin. Cancer Res. 2009, 15, 3462–3471. [Google Scholar] [CrossRef] [PubMed]
- Yoshiba, S.; Ito, D.; Nagumo, T.; Shirota, T.; Hatori, M.; Shintani, S. Hypoxia induces resistance to 5-fluorouracil in oral cancer cells via G1 phase cell cycle arrest. Oral Oncol. 2009, 45, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Onishi, H.; Morifuji, Y.; Kai, M.; Suyama, K.; Iwasaki, H.; Katano, M. Hedgehog inhibitor decreases chemosensitivity to 5-fluorouracil and gemcitabine under hypoxic conditions in pancreatic cancer. Cancer Sci. 2012, 103, 1272–1279. [Google Scholar] [CrossRef] [PubMed]
- Na, Y. Recent cancer drug development with xanthone structures. J. Pharm. Pharmacol. 2009, 61, 707–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrippina, W.R.G.; Widiyanti, P.; Yusuf, H. Synthesis and characterization of bacterial cellulose-Garcinia mangostana extract as anti breast cancer biofilm candidate. J. Biomim. Biomater. Biomed. Eng. 2017, 30, 76–85. [Google Scholar] [CrossRef]
- Nanasombat, S.; Kuncharoen, N.; Ritcharoon, B.; Sukcharoen, P. Antibacterial activity of Thai medicinal plant extracts against oral and gastrointestinal pathogenic bacteria and prebiotic effect on the growth of Lactobacillus acidophilus. Chiang Mai J. Sci. 2018, 45, 33–44. [Google Scholar]
- Karim, N.; Jeenduang, N.; Tangpong, J. Renoprotective Effects of Xanthone Derivatives from Garcinia mangostana Against High Fat Diet and Streptozotocin-Induced Type II Diabetes in Mice. Walailak J. Sci. Technol. 2016, 15, 107–116. [Google Scholar]
- Chatatikun, M.; Chiabchalard, A. Thai plants with high antioxidant levels, free radical scavenging activity, anti-tyrosinase and anti-collagenase activity. BMC Complement. Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.-S.; You, B.H.; Song, J.; Ko, H.W.; Choi, Y.H.; Chin, Y.-W. Mangosteen Extract Prevents Dextran Sulfate Sodium-Induced Colitis in Mice by Suppressing NF-κB Activation and Inflammation. J. Med. Food 2017, 20, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Boonprom, P.; Boonla, O.; Chayaburakul, K.; Welbat, J.U.; Pannangpetch, P.; Kukongviriyapan, U.; Kukongviriyapan, V.; Pakdeechote, P.; Prachaney, P. Garcinia mangostana pericarp extract protects against oxidative stress and cardiovascular remodeling via suppression of p47phox and iNOS in nitric oxide deficient rats. Ann. Anat. 2017, 212, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Karim, N.; Jeenduang, N.; Tangpong, J. Anti-Glycemic and Anti-Hepatotoxic Effects of Mangosteen Vinegar Rind from Garcinia mangostana Against HFD/STZ-Induced Type II Diabetes in Mice. Pol. J. Food Nutr. Sci. 2018, 68, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.K.; Huang, Y.L.; Chen, C.C. Garcinone E, a xanthone derivative, has potent cytotoxic effect against hepatocellular carcinoma cell lines. Planta Med. 2002, 68, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Akao, Y.; Ohguchi, K.; Ito, T.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Xanthones induce cell-cycle arrest and apoptosis in human colon cancer DLD-1 cells. Bioorg. Med. Chem. 2005, 13, 6064–6069. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.J.; Petiwala, S.M.; Syed, D.N.; Rasmussen, J.T.; Adhami, V.M.; Siddiqui, I.A.; Kohl, A.M.; Mukhtar, H. α-Mangostin, a xanthone from mangosteen fruit, promotes cell cycle arrest in prostate cancer and decreases xenograft tumor growth. Carcinogenesis 2011, 33, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Huo, X.; Duan, W.; Xu, Q.; Li, R.; Ma, J.; Li, X.; Han, L.; Li, W.; Sun, H.; et al. α-Mangostin inhibits hypoxia-driven ROS-induced PSC activation and pancreatic cancer cell invasion. Cancer Lett. 2014, 347, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delvecchio, C.; Tiefenbach, J.; Krause, H.M. The zebrafish: A powerful platform for in vivo, HTS drug discovery. Assay Drug Dev. Technol. 2011, 9, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.C.; Rouhi, P.; Jensen, L.D.; Zhang, D.; Ji, H.; Hauptmann, G.; Ingham, P.; Cao, Y. Hypoxia-induced pathological angiogenesis mediates tumor cell dissemination, invasion, and metastasis in a zebrafish tumor model. Proc. Natl. Acad. Sci. USA 2009, 106, 19485–19490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Huang, J.; Ye, J. A fresh look at zebrafish from the perspective of cancer research. J. Exp. Clin. Cancer Res. 2015, 34, 80. [Google Scholar] [CrossRef] [PubMed]
- Tulotta, C.; He, S.; Van Der Ent, W.; Chen, L.; Groenewoud, A.; Spaink, H.P.; Snaar-Jagalska, B.E. Imaging cancer angiogenesis and metastasis in a zebrafish embryo model. In Cancer and Zebrafish; Springer: Cham, Switzerland, 2016; pp. 239–263. ISBN 978-3-319-30654-4. [Google Scholar]
- Chen, L.; Groenewoud, A.; Tulotta, C.; Zoni, E.; Kruithof-de Julio, M.; van der Horst, G.; van der Pluijm, G.; Snaar-Jagalska, B.E. A zebrafish xenograft model for studying human cancer stem cells in distant metastasis and therapy response. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2017; Volume 138, pp. 471–496. ISSN 0091-679X. [Google Scholar]
- Lam, S.H.; Wu, Y.L.; Vega, V.B.; Miller, L.D.; Spitsbergen, J.; Tong, Y.; Zhan, H.; Govindarajan, K.R.; Lee, S.; Mathavan, S.; et al. Conservation of gene expression signatures between zebrafish and human liver tumors and tumor progression. Nat. Biotechnol. 2006, 24, 73. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.B. HPLC analysis of selected xanthones in mangosteen fruit. J. Sep. Sci. 2007, 30, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, M.; Ninomiya, K.; Tagashira, Y.; Maejima, K.; Yoshida, T.; Amakura, Y. Polyphenolic constituents of the pericarp of mangosteen (Garcinia mangostana L.). J. Agric. Food Chem. 2015, 63, 7670–7674. [Google Scholar] [CrossRef] [PubMed]
- Gatei, M.; Jacob, B.; Chen, P.; Kijas, A.W.; Bacherel, O.J.; Guaven, N.; Birrel, G.; Lee, J.; Paul, T.T.; Lerenthal, Y.; et al. ATM protein-dependent phosphorylation of Rad50 protein regulates DNA repair and cell cycle control. J. Biol. Chem. 2011, 286, 31542–31556. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wang, J.; Xu, S.; Xiao, L.; Chen, G.; Zhang, W.; Li, J. Radioprotective effect on HepG2 cells of low concentrations of cobalt chloride: Induction of hypoxia-inducible factor-1 α and clearance of reactive oxygen species. J. Radiat. Res. 2013, 54, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Muniandy, S.; Daud, F.; Senafi, S.; Noor, M.M.; Kumaran, M.; Alwi, A.N.A.M.; Long, A.Y.Y.J.; Zuki, R.B.M.; Isac, J.S.; Krishnan, S.; et al. Active compound, antioxidant, antiproliferative and effect on stz induced zebrafish of various crude extracts from Boletus Qriseipurpureus. Malays. Appl. Biol. 2016, 45, 69–80. [Google Scholar]
- OECD. Draft Proposal for A New Guideline—Fish Embryo Toxicity (FET) Test; Draft Guideline; Organization for Economic Cooperation and Development: Paris, France, 2006. [Google Scholar]
- OECD. Draft Proposal for A New Guideline—Fish Embryo Toxicity (FET) Test; Draft Guideline; Organization for Economic Cooperation and Development: Paris, France, 2013. [Google Scholar]
- Gutierrez-Orozco, F.; Failla, M.L. Biological activities and bioavailability of mangosteen xanthones: A critical review of the current evidence. Nutrients 2013, 5, 3163–3183. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, Y.; Wang, W.; Deng, L. Bioactivity and pharmacological Properties of α-mangostin from the mangosteen fruit: A review. Expert Opin. Ther. Pat. 2018, 28, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Syam, S.; Bustamam, A.; Abdullah, R.; Sukari, M.A.; Hashim, N.M.; Ghaderian, M.; Rahmani, M.; Mohan, S.; Abdelwahab, S.I.; Ali, H.M. β-Mangostin induces p53-dependent G2/M cell cycle arrest and apoptosis through ROS mediated mitochondrial pathway and NfkB suppression in MCF-7 cells. J. Funct. Foods 2014, 6, 290–304. [Google Scholar] [CrossRef]
- Liu, Z.; Antalek, M.; Nguyen, L.; Li, X.; Tian, X.; Le, A.; Zi, X. The effect of gartanin, a naturally occurring xanthone in mangosteen juice, on the mTOR pathway, autophagy, apoptosis, and the growth of human urinary bladder cancer cell lines. Nutr. Cancer 2013, 65 (Suppl. 1), 68–77. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Y.; Wang, S.N.; Yang, X.H.; Lan, W.J.; Chen, Z.W.; Chen, J.K.; Xie, J.H.; Han, Y.F.; Pi, R.B.; Yang, X.B. Gartanin protects neurons against glutamate-induced cell death in HT22 cells: Independence of Nrf-2 but involvement of HO-1 and AMPK. Neurochem. Res. 2016, 41, 2267–2277. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Ma, Q.; Guo, K.; Liu, J.; Li, W.; Wang, F.; Wu, E. Xanthones from mangosteen extracts as natural chemopreventive agents: Potential anticancer drugs. Curr. Mol. Med. 2011, 11, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Chitchumroonchokchai, C.; Thomas, J.L.; Gushchina, L.V.; DiSilvestro, D.; Failla, M.L.; Ziouzenkova, O. Adipocyte reporter assays: Application for identification of anti-inflammatory and antioxidant properties of mangosteen xanthones. Mol. Nutr. Food Res. 2014, 58, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Averill-Bates, D.; Grondin, M.; Ouellet, F. Activation of apoptosis signaling pathways by reactive oxygen species. Cryobiology 2018, 80, 170. [Google Scholar] [CrossRef]
- Hamanaka, R.B.; Chandel, N.S. Mitochondrial reactive oxygen species regulate hypoxic signaling. Curr. Opin. Cell Biol. 2009, 21, 894–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunbeck, T.; Lammer, E. Fish embryos toxicity assays. Ger. Fed. Environ. Agency 2006, 136, 1–298. [Google Scholar]
- Braunbeck, T.; Böttcher, M.; Hollert, H.; Kosmehl, T.; Lammer, E.; Leist, E.; Rudolf, M.; Seitz, N. Towards an alternative for the acute fish LC50 test in chemical assessment: The fish embryo toxicity test goes multi-species an update. Altex 2005, 22, 87–102. [Google Scholar] [PubMed]
- Lammer, E.; Carr, G.J.; Wendler, K.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T.H. Is the fish embryo toxicity test (FET) with the zebrafish (Danio rerio) a potential alternative for the fish acute toxicity test? Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Abreu, J.G.; Coffinier, C.; Larraın, J.; Oelgeschläger, M.; De Robertis, E.M. Chordin-like CR domains and the regulation of evolutionarily conserved extracellular signaling systems. Gene 2002, 287, 39–47. [Google Scholar] [CrossRef]
- Wilkinson, L.; Gilbert, T.; Kinna, G.; Ruta, L.A.; Pennisi, D.; Kett, M.; Little, M.H. Crim1KST264/KST264 mice implicate Crim1 in the regulation of vascular endothelial growth factor-A activity during glomerular vascular development. J. Am. Soc. Nephrol. 2007, 18, 1697–1708. [Google Scholar] [CrossRef] [PubMed]
- Kolle, G.; Jansen, A.; Yamada, T.; Little, M. In vivo electroporation of Crim1 in the developing chick spinal cord. Dev. Dyn. 2003, 226, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Kinna, G.; Kolle, G.; Carter, A.; Key, B.; Lieschke, G.J.; Perkins, A.; Little, M.H. Knockdown of zebrafish crim1 results in a bent tail phenotype with defects in somite and vascular development. Mech. Dev. 2006, 123, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Tang, L. CRIM1, the antagonist of BMPs, is a potential risk factor of cancer. Curr. Cancer Drug Targets 2014, 14, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Zhang, Y.; Yi, Q.; Wu, Y.; Wan, R.; Tang, L. CRIM1, a newfound cancer-related player, regulates the adhesion and migration of lung cancer cells. Growth Factors 2015, 33, 384–392. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazry, S.; Noordin, M.A.M.; Sanusi, S.; Noor, M.M.; Aizat, W.M.; Lazim, A.M.; Dyari, H.R.E.; Jamar, N.H.; Remali, J.; Othman, B.A.; et al. Cytotoxicity and Toxicity Evaluation of Xanthone Crude Extract on Hypoxic Human Hepatocellular Carcinoma and Zebrafish (Danio rerio) Embryos. Toxics 2018, 6, 60. https://doi.org/10.3390/toxics6040060
Fazry S, Noordin MAM, Sanusi S, Noor MM, Aizat WM, Lazim AM, Dyari HRE, Jamar NH, Remali J, Othman BA, et al. Cytotoxicity and Toxicity Evaluation of Xanthone Crude Extract on Hypoxic Human Hepatocellular Carcinoma and Zebrafish (Danio rerio) Embryos. Toxics. 2018; 6(4):60. https://doi.org/10.3390/toxics6040060
Chicago/Turabian StyleFazry, Shazrul, Muhammad Akram Mohd Noordin, Salahuddin Sanusi, Mahanem Mat Noor, Wan Mohd Aizat, Azwan Mat Lazim, Herryawan Ryadi Eziwar Dyari, Nur Hidayah Jamar, Juwairiah Remali, Babul Airianah Othman, and et al. 2018. "Cytotoxicity and Toxicity Evaluation of Xanthone Crude Extract on Hypoxic Human Hepatocellular Carcinoma and Zebrafish (Danio rerio) Embryos" Toxics 6, no. 4: 60. https://doi.org/10.3390/toxics6040060
APA StyleFazry, S., Noordin, M. A. M., Sanusi, S., Noor, M. M., Aizat, W. M., Lazim, A. M., Dyari, H. R. E., Jamar, N. H., Remali, J., Othman, B. A., Law, D., Sidik, N. M., Cheah, Y. H., & Lim, Y. C. (2018). Cytotoxicity and Toxicity Evaluation of Xanthone Crude Extract on Hypoxic Human Hepatocellular Carcinoma and Zebrafish (Danio rerio) Embryos. Toxics, 6(4), 60. https://doi.org/10.3390/toxics6040060