Health Impact Assessment of Sulfolane on Embryonic Development of Zebrafish (Danio rerio)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry and Embryo Collection
2.2. Sulfolane Treatment
2.3. Morphometric Assessment
2.4. Behavioural Assessment
2.5. Transcript Abundance
2.6. Statistics
3. Results
3.1. Morphometric Assessment
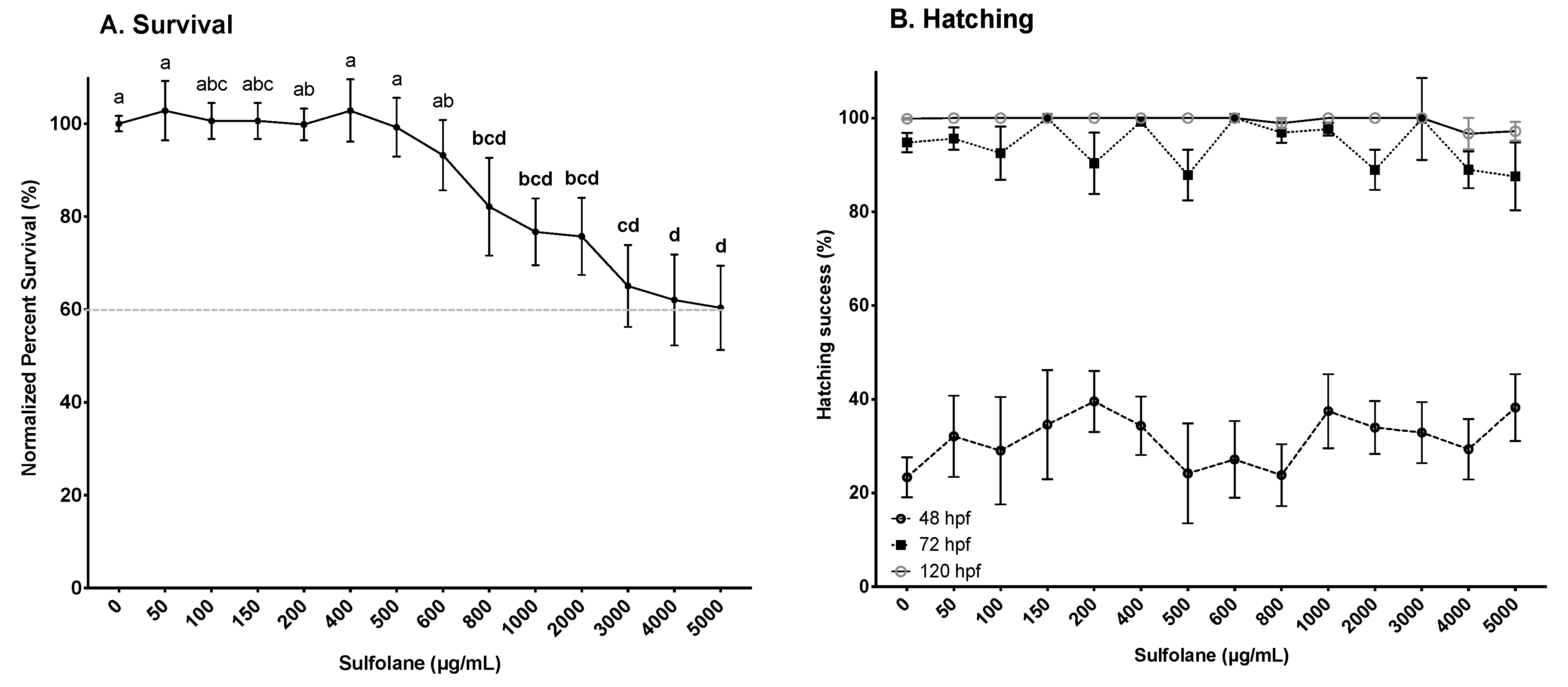
3.1.1. Survival and Hatching
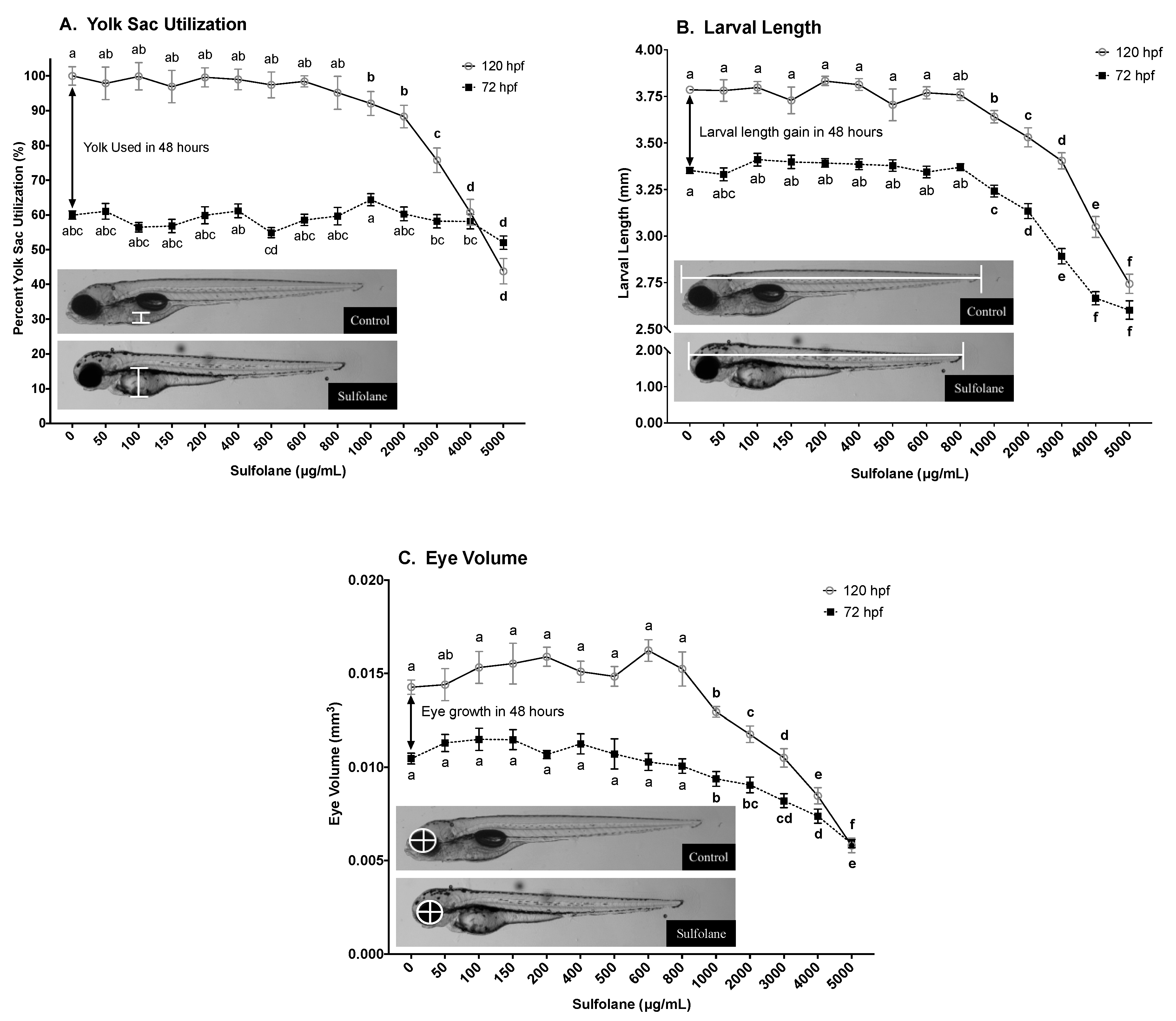
3.1.2. Yolk Sac Utilisation
3.1.3. Larval Length
3.1.4. Larval Eye Volume
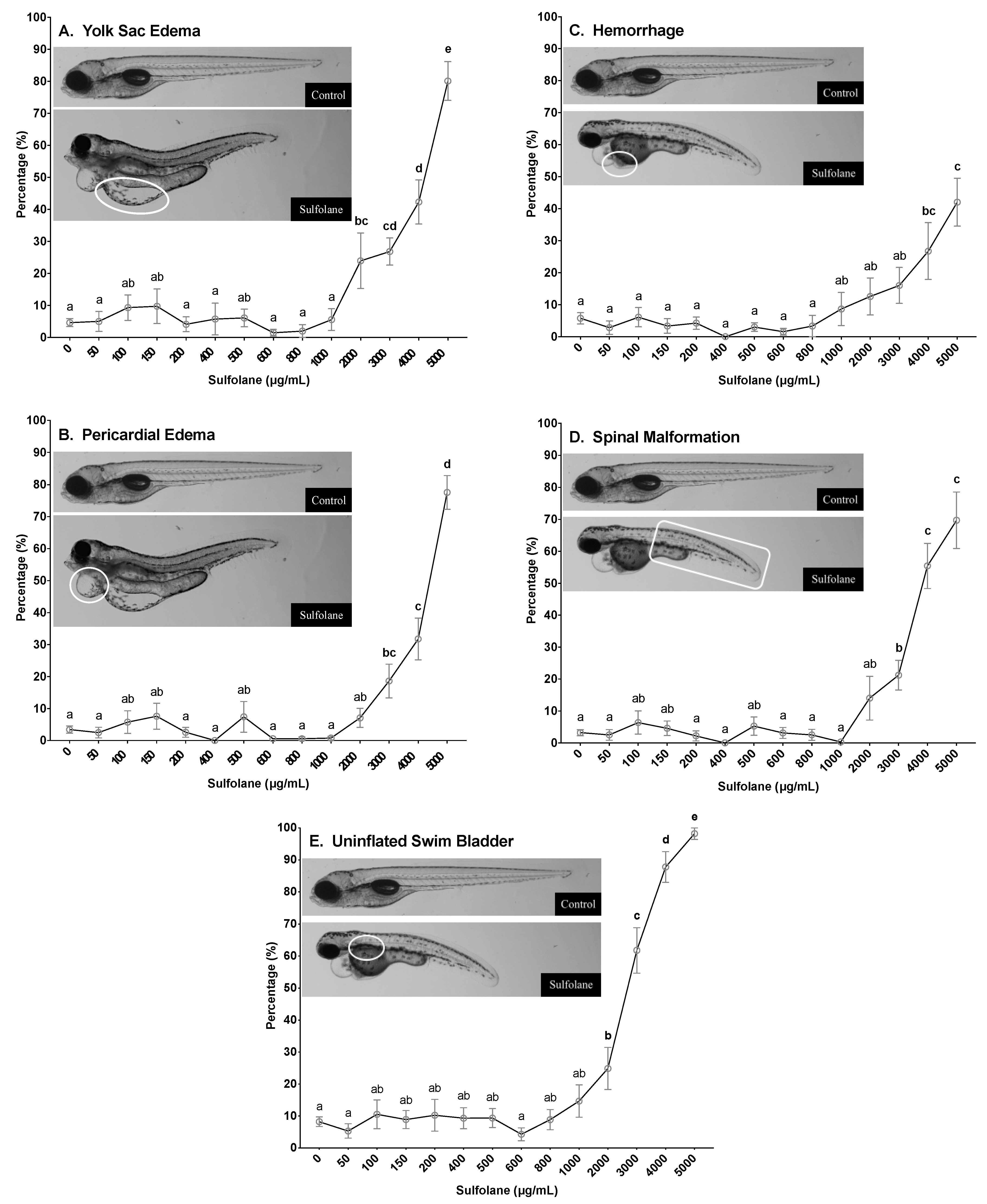
3.1.5. Yolk Sac and Pericardial Oedemas
3.1.6. Haemorrhaging
3.1.7. Spinal Malformations
3.1.8. Swim Bladder Malformations
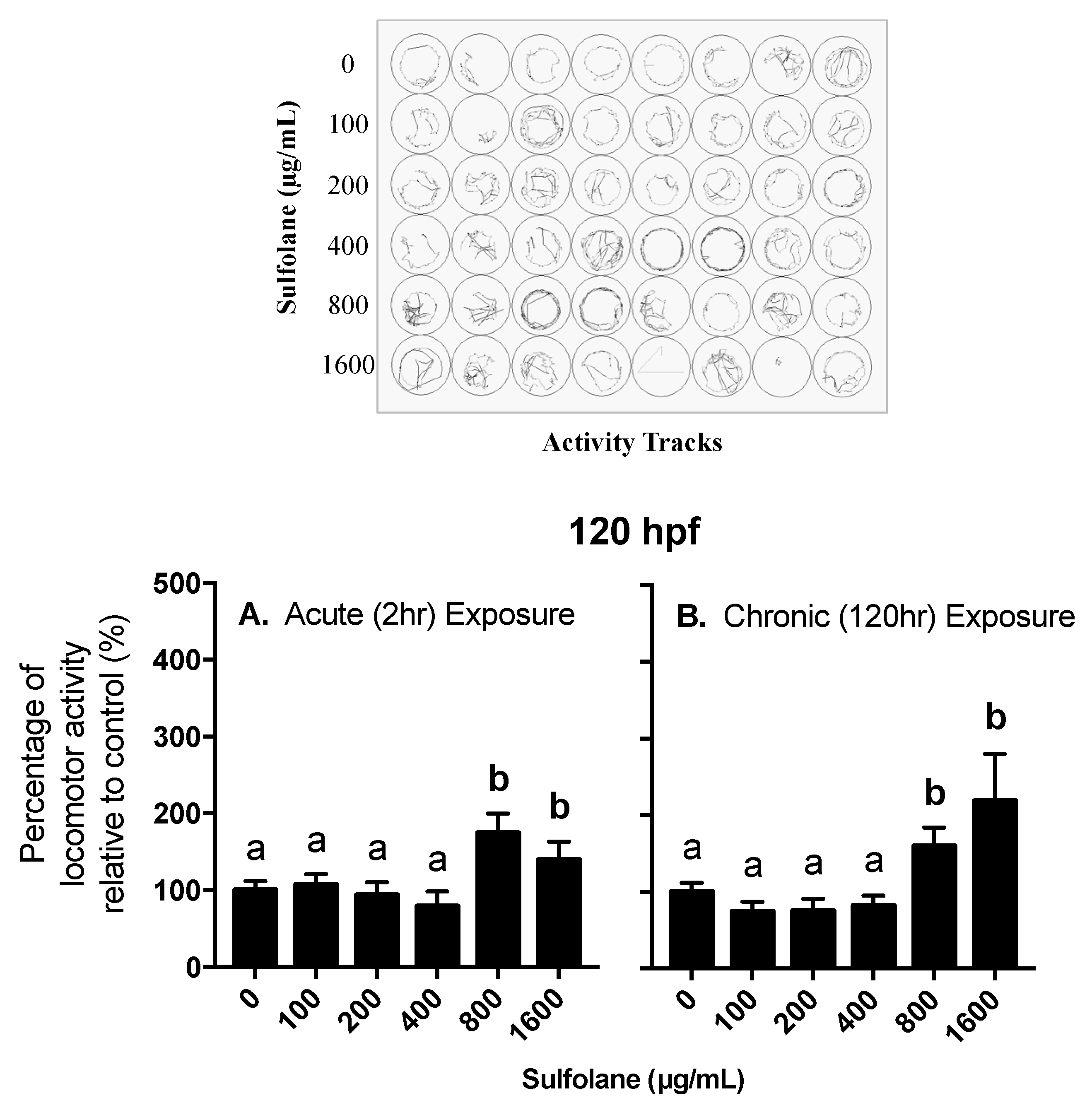
3.2. Behavioural Assessment
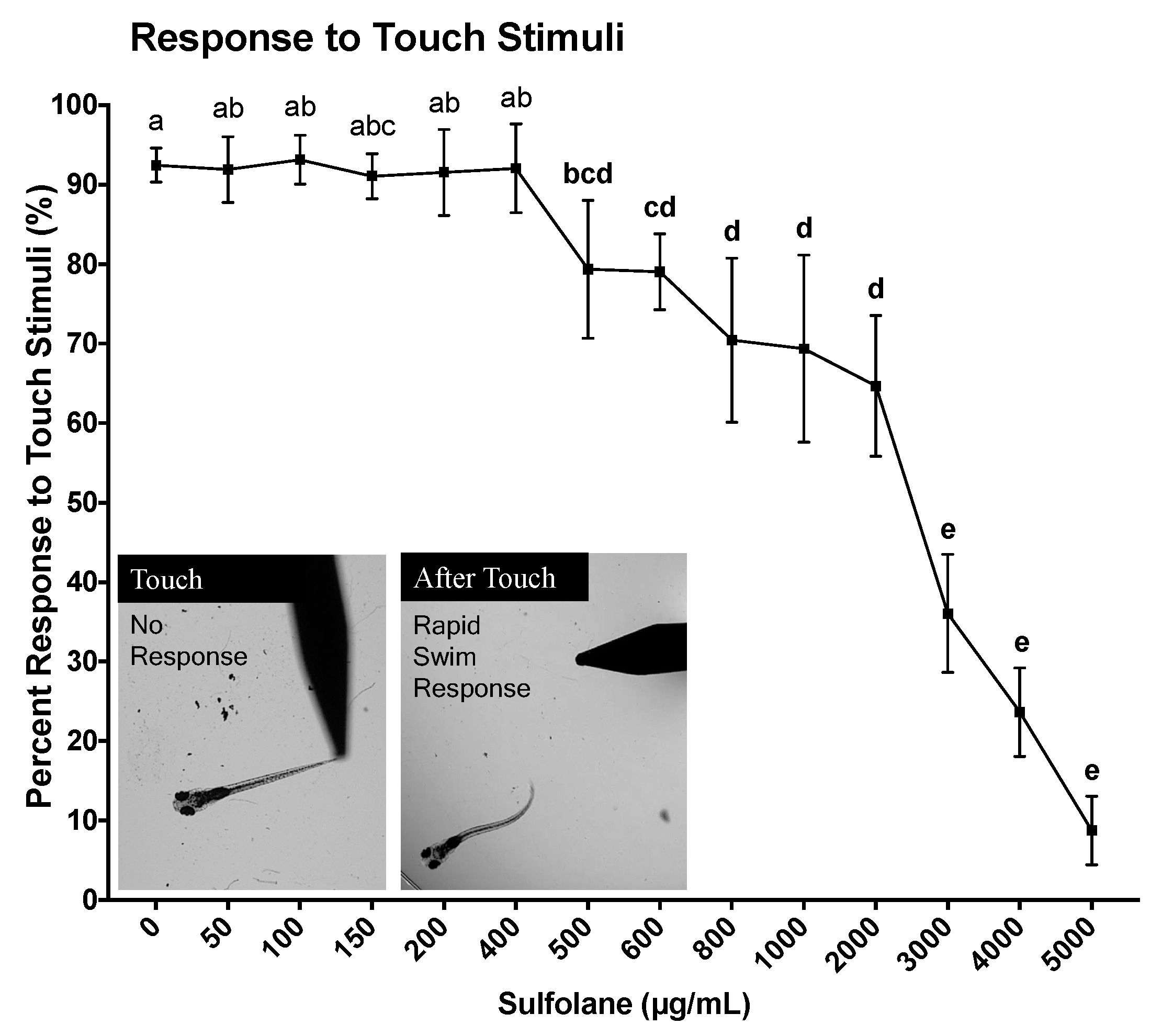
3.2.1. Response to Touch Stimuli
3.2.2. Locomotor Activity
3.3. Transcript Abundance
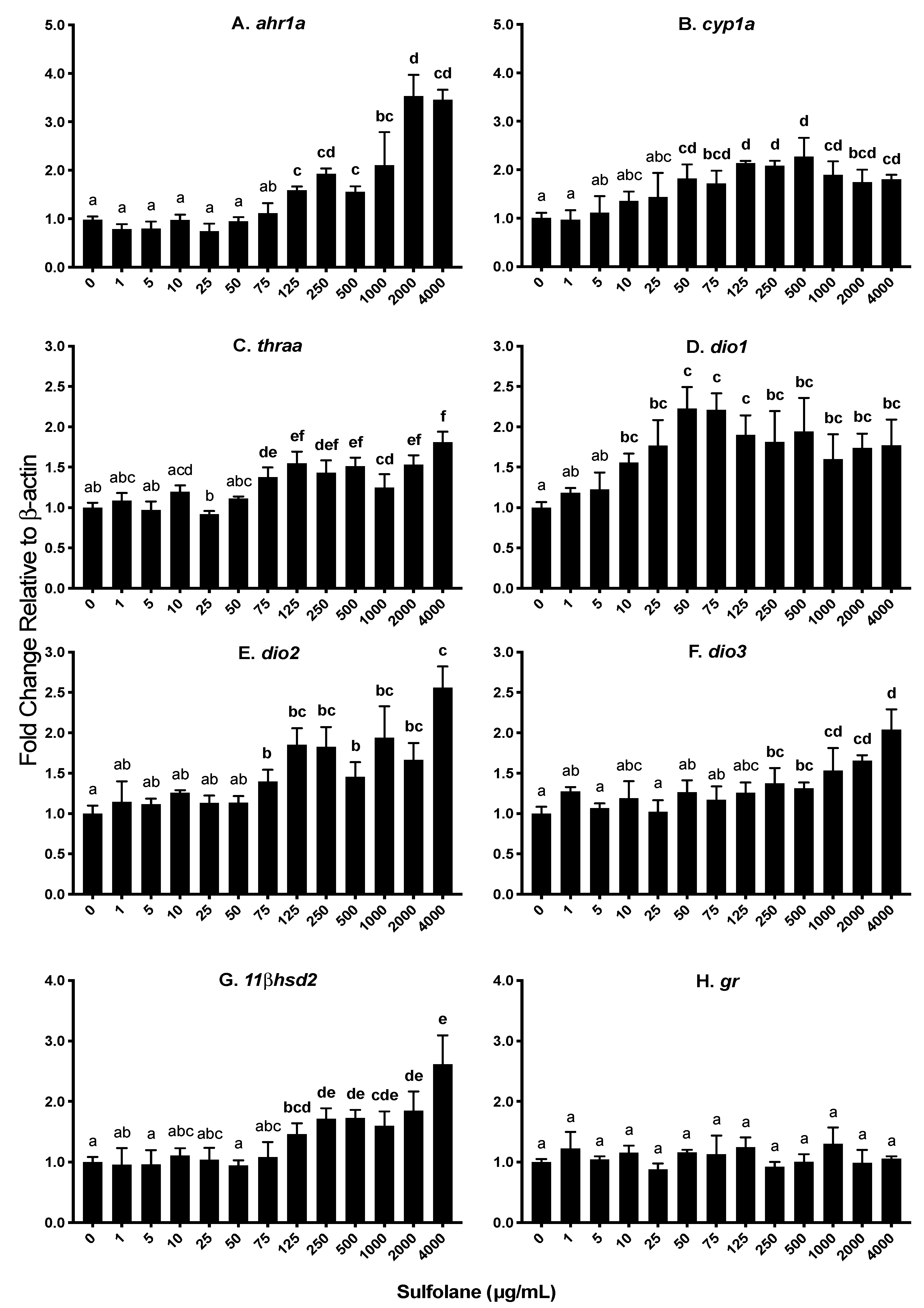
3.3.1. Aryl hydrocarbon receptor (ahr1a), Cytochrome p450 (cyp1a) and Heat shock protein (hsp70)
3.3.2. Thyroid hormone receptor (thraa) and Deiodinases 1, 2, and 3 (dio1, dio2, dio3)
3.3.3. 11 β Dehydrogenase (11βhsd2) and Glucocorticoid receptor (gr)
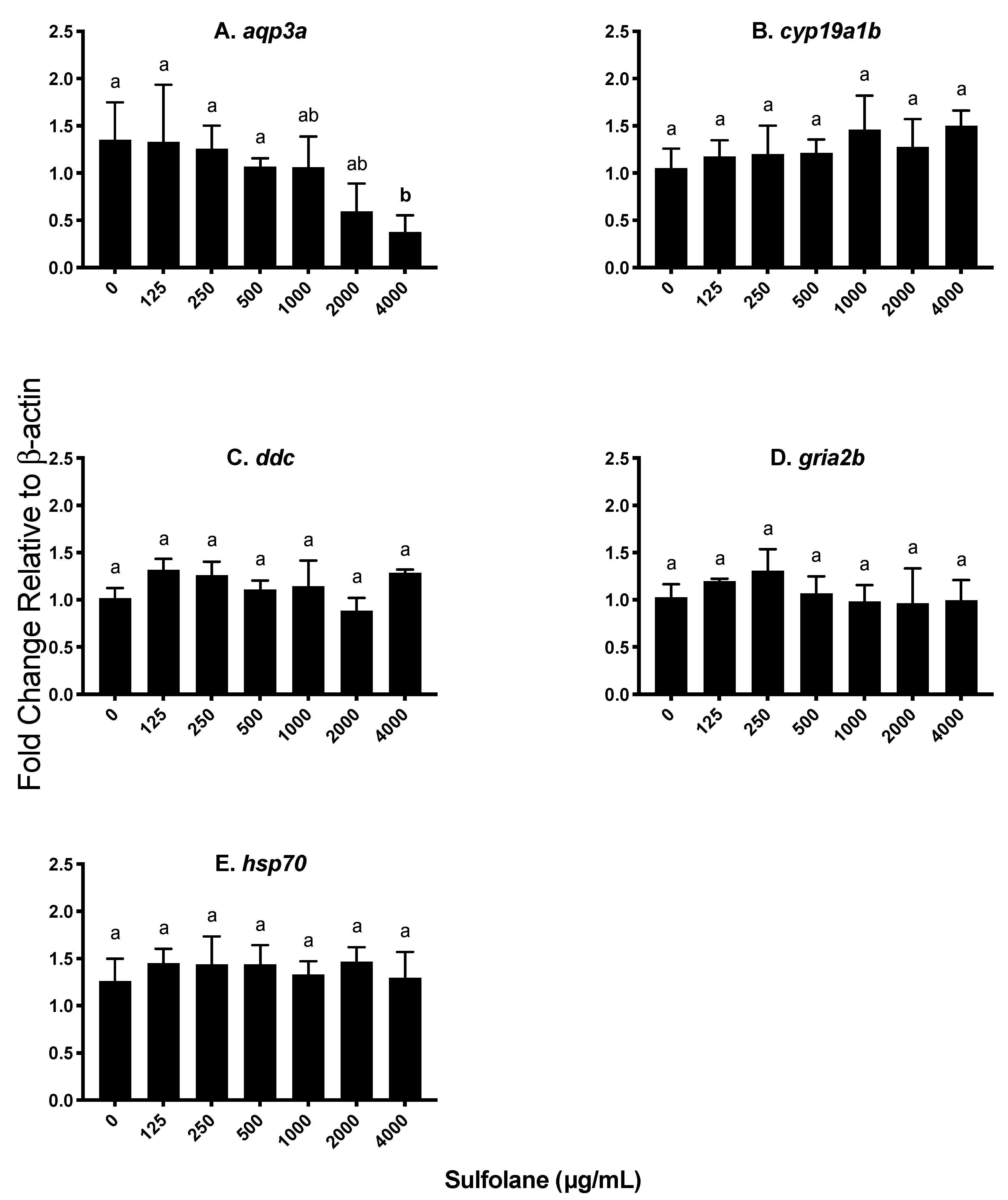
3.3.4. Aquaporin (aqp3a)
3.3.5. Other Endocrine Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CCME (Canadian Council of Ministers of the Environment). Canadian Water Quality Guidelines for the Protection of Aquatic Life. 2005. Available online: http://ceqg-rcqe.ccme.ca/download/en/212 (accessed on 12 July 2016).
- Pierotti, G.J.; Dunn, C.L. Solvent Extraction Process Patent. U.S. Patent #US2360861A, 24 October 1944. [Google Scholar]
- Deal, C.; Evans, H.; Oliver, E.; Papadopoulus, M. Extraction of Aromatics with Sulfolane. In Proceedings of the 5th World Petroleum Congress, New York, NY, USA, 30 May–5 June 1959. [Google Scholar]
- Tilstam, U. Sulfolane: A versatile dipolar aprotic solvent. Org. Process Res. Dev. 2012, 16, 1273–1278. [Google Scholar] [CrossRef]
- Blystone, C. National Toxicology Program-Sulfolane. 2011. Available online: https://ntp.niehs.nih.gov/ntp/about_ntp/bsc/2011/december/presentations/5_blystonesulfolane.pdf (accessed on 6 September 2016).
- Thompson, C.M.; Gaylor, D.W.; Tachovsky, J.A.; Perry, C.; Carakostas, M.C.; Haws, L.C. Development of a chronic noncancer oral reference dose and drinking water screening level for sulfolane using benchmark dose modeling. J. Appl. Toxicol. 2013, 33, 1395–1406. [Google Scholar] [CrossRef]
- Clark, E. Sulfolane and sulfones. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons: Hoboken, NJ, USA, 2000. [Google Scholar]
- Greene, E.A.; Gieg, L.M.; Coy, D.L.; Fedorak, P.M. Sulfolane biodegradation potential in aquifer sediments at sour natural gas plant sites. Water Res. 1998, 32, 3680–3688. [Google Scholar] [CrossRef]
- Gieg, L.M.; Greene, E.A.; Coy, D.L.; Fedorak, P.M. Diisopropanolamine biodegradation potential at sour gas plants. Groundw. Monit. Remediat. 1998, 18, 158–173. [Google Scholar] [CrossRef]
- Advisian. Waterton Complex Groundwater Monitoring Annual Report; File No. 407074-00722; Shell Canada Energy: Calgary, AB, Canada, 2016. [Google Scholar]
- LDEQ (Louisiana Department of Environmental Quality). Environmental Indicator Events CA 725 & CA 750; Unites States Environmental Protection Agency: Washington, DC, USA, 2002. [Google Scholar]
- Kim, C.C.; Clarke, W.P.; Lockington, D. Anaerobic Biodegradation of Sulfolane in Soil Environment. Environ. Eng. Res. 1999, 4, 185–193. [Google Scholar]
- Agatonovic, V.; Vaisman, E. Sulfolane Impacted Soil and Groundwater Treatibility Study. 2005. Available online: http://www.esaa.org/wp-content/uploads/2015/06/05-Agatonovic.pdf (accessed on 5 January 2017).
- TTC Labs Inc. Sulfolane Technical Assistance and Evaluation; OASIS Environmental: Anchorage, AK, USA, April 2010. [Google Scholar]
- OASIS (Oasis Environmental Inc.). Sulfolane Technical Assistance and Evaluation Report. 2010. Available online: dec.alaska.gov/spar/csp/sites/north-pole-refinery/docs/sulfolanereportfinal.pdf (accessed on 14 June 2018).
- Chou, C.C.; Swatloski, R.A. Biodegradation of sulfolane in refinery wastewater. In Proceedings of the 37th Industrial Waste Conference, West Lafayette, IN, USA, 11–13 May 1982; Purdue University: West Lafayette, IN, USA, 1983; pp. 559–566. [Google Scholar]
- Andersen, M.E.; Jones, R.A.; Kurlansik, L.; Mehl, R.G.; Jenkins, L.J. Sulfolane-induced convulsions in rodents. Res. Commun. Chem. Pathol. Pharmacol. 1976, 15, 571–580. [Google Scholar]
- Zhu, Z.H.; Sun, M.L.; Li, Z.S.; Yang, Z.C.; Zhang, T.B.; Heng, Z.C.; Xiao, B.L.; Li, Q.Y.; Peng, Q.Y.; Dong, Y.H. An investigation of maximum allowable concentration of sulfolane in surface water. J. West China Univ. Med. Soc. 1987, 18, 376–380. [Google Scholar]
- ERAC (Environmental Research Advisory Council). Toxicity Assessment of Sulfolane and Diisopropanolamine; HydroQual Laboratories and Golder Associates: Calgary, AB, Canada, 1998. [Google Scholar]
- CAPP (Canadian Association of Petroleum Producers). Soil and Water Quality Guidelines for Sulfolane and Di-Isopropanolamine (DIPA): Environmental and Human Health; File No. C50560000; Komex International Ltd.: Calgary, AB, Canada, 2001. [Google Scholar]
- Bridié, A.L.; Wolff, C.J.M.; Winter, M. The acute toxicity of some petrochemicals to goldfish. Water Res. 1979, 13, 623–626. [Google Scholar] [CrossRef]
- Shell Oil Company. FYI-OTS-0484-0304S, Supplement. Sequence E. Technical Bulletin IC:71-20-Sulfolane; Office of Toxic Substances. U.S. Environmental Protection Agency: Washington, DC, USA, 1984. [Google Scholar]
- Girling, A.E. Validation of a Method for Determining the Acute Toxicity of Test Substances to the Marine Copepod Acartia Tonsa and Its Application to the Testing of Oil-Based Products; Group Research Report. SBGR 87082; Shell Research Limited, Sittingborne Research Centre: London, UK, 1987. [Google Scholar]
- CCME (Canadian Council of Ministers of the Environment). Canadian Environmental Quality Guidelines for Sulfolane: Water and Soil Scientific Supporting Document. 2006. Available online: https://www.ccme.ca/files/Resources/supporting_scientific_documents/sulfolane_ssd_soil_water_1.1_e.pdf (accessed on 12 July 2016).
- ATSDR (Agency for Toxic Substances and Disease Registry). Health Consultation: Sulfolane. 2011. Available online: https://ntp.niehs.nih.gov/ntp/noms/support_docs/atsdrsulfolanehc_2011_508.pdf (accessed on 23 July 2019).
- CCME (Canadian Council of Ministers of the Environment). CCME Summary Table: Sulfolane Factsheet. Available online: http://st-ts.ccme.ca/en/index.html?lang=en&factsheet=203 (accessed on 15 July 2019).
- Goverment of Alberta. Guidance for Selecting Toxicity Reference Values for Alberta Tier 1 and Tier 2 Soil and Groundwater Remediation Guidelines. 2017. Available online: https://open.alberta.ca/dataset/5d56d9c3-5642-40f7-ab61-73feabf09241/resource/2cb0d119-1fc1-4e60-b285-630f6a8ee097/download/guidancetoxicityvaluestier1and2-2017.pdf (accessed on 23 July 2019).
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD Publishing: Paris, France, 2013; pp. 1–22. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef]
- Kunz, Y.W. Developmental Biology of Teleost Fishes; Springer: Dordrecht, The Netherlands, 2004; pp. 457–504. [Google Scholar]
- Rubinstein, A.L. Zebrafish: From disease modeling to drug discovery. Curr. Opin. Drug Discov. Dev. 2003, 6, 218–223. [Google Scholar]
- Fraysse, B.; Mons, R.; Garric, J. Development of a zebrafish 4-day embryo-larval bioassay to assess toxicity of chemicals. Ecotoxicol. Environ. Saf. 2006, 63, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.N.; Lindsey, B.W.; Dumbarton, T.C.; Croll, R.P.; Smith, F.M. The contribution of the swimbladder to buoyancy in the adult zebrafish (Danio rerio): A morphometric analysis. J. Morphol. 2008, 269, 666–673. [Google Scholar] [CrossRef]
- Vargas, R.A.; Vásquez, I.C. Morphometric parameters to evaluate zebrafish heart: A model to study cardiovascular function. FASEB J. 2016, 30 (Suppl. 1), 556. [Google Scholar]
- Chhetri, J.; Jacobson, G.; Gueven, N. Zebrafish—On the move towards ophthalmological research. Eye 2014, 28, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Padilla, S.; MacPhail, R. Using zebrafish to assess developmental neurotoxicity. In Reproductive and Developmental Toxicology; Academic Press: Cambridge, MA, USA, 2011; pp. 179–191. [Google Scholar]
- Selderslaghs, I.W.T.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Kinch, C.D.; Ibhazehiebo, K.; Jeong, J.-H.; Habibi, H.R.; Kurrasch, D.M. Low-dose exposure to bisphenol A and replacement bisphenol S induces precocious hypothalamic neurogenesis in embryonic zebrafish. Proc. Natl. Acad. Sci. USA 2015, 112, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Best, C.; Kurrasch, D.M.; Vijayan, M.M. Maternal cortisol stimulates neurogenesis and affects larval behaviour in zebrafish. Sci. Rep. 2017, 7, 40905. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.F.; Yu, L.; Achari, G.; Tay, J.H. Degradation of sulfolane in aqueous media by integrating activated sludge and advanced oxidation process. Chemosphere 2019, 222, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Topic Popovic, N.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Barisic, J.; Jadan, M.; Persin Berakovic, A.; Sauerborn Klobucar, R. Tricaine methane-sulfonate (MS-222) application in fish anaesthesia. J. Appl. Ichthyol. 2012, 28, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Rombough, P.J. Ontogenetic changes in the toxicity and efficacy of the anaesthetic MS222 (tricaine methanesulfonate) in zebrafish (Danio rerio) larvae. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 148, 463–469. [Google Scholar] [CrossRef]
- Collymore, C.; Tolwani, A.; Lieggi, C.; Rasmussen, S. Efficacy and safety of 5 anesthetics in adult zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 198–203. [Google Scholar] [PubMed]
- Peterson, S.M.; Freeman, J.L. RNA isolation from embryonic zebrafish and cDNA synthesis for gene expression analysis. J. Vis. Exp. 2009, 30, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Pohlert, T. Calculate Pairwise Multiple Comparisons of Mean Rank Sums (Version 31). 2018. Available online: https://cran.r-project.org/web/packages/PMCMRplus/PMCMRplus.pdf (accessed on 13 April 2018).
- Wilson, C. Aspects of Larval Rearing. ILAR J. 2012, 53, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, G.N.; McGee, C.A.S.; Dumbarton, T.C.; Croll, R.P.; Smith, F.M. Development of the swimbladder and its innervation in the zebrafish, Danio rerio. J. Morphol. 2007, 268, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Colwill, R.M.; Creton, R. Locomotor behaviors in zebrafish (Danio rerio) larvae. Behav. Process. 2011, 86, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Budick, S.A.; O’Malley, D.M. Locomotor repertoire of the larval zebrafish: Swimming, turning and prey capture. J. Exp. Biol. 2000, 203, 2565–2579. [Google Scholar] [PubMed]
- Goolish, E.M.; Okutake, K. Lack of gas bladder inflation by the larvae of zebrafish in the absence of an air-water interface. J. Fish Biol. 1999, 55, 1054–1063. [Google Scholar]
- Hill, A.J.; Bello, S.M.; Prasch, A.L.; Peterson, R.E.; Heideman, W. Water permeability and TCDD-induced edema in Zebrafish early-life stages. Toxicol. Sci. 2004, 78, 78–87. [Google Scholar] [CrossRef]
- Hanke, N.; Staggs, L.; Schroder, P.; Litteral, J.; Fleig, S.; Kaufeld, J.; Pauli, C.; Haller, H.; Schiffer, M. ‘Zebrafishing’ for novel genes relevant to the glomerular filtration barrier. Biomed Res. Int. 2013, 2013, 658270. [Google Scholar] [CrossRef]
- Kwong, R.W.M.; Kumai, Y.; Perry, S.F. The Role of Aquaporin and Tight Junction Proteins in the Regulation of Water Movement in Larval Zebrafish (Danio rerio). PLoS ONE 2013, 8, e70764. [Google Scholar] [CrossRef]
- Sant, K.E.; Timme-Laragy, A.R. Zebrafish as a Model for Toxicological Perturbation of Yolk and Nutrition in the Early Embryo. Curr. Environ. Heal. Rep. 2018, 5, 125–133. [Google Scholar] [CrossRef]
- Fernandez-Salguero, P.M.; Hilbert, D.M.; Rudikoff, S.; Ward, J.M.; Gonzalez, F.J. Aryl-hydrocarbon receptor-deficient mice are resistant to 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced toxicity. Toxicol. Appl. Pharmacol. 1996, 140, 173–179. [Google Scholar] [CrossRef]
- Colavecchia, M.V.; Hodson, P.V.; Parrott, J.L. The relationships among CYP1A induction, toxicity, and eye pathology in early life stages of fish exposed to oil sands. J. Toxicol. Environ. Health. Part A 2007, 70, 1542–1555. [Google Scholar] [CrossRef] [PubMed]
- Beyers, D.W.; Rice, J.A.; Clements, W.H.; Henry, C.J. Estimating physiological cost of chemical exposure: Integrating energetics and stress to quantify toxic effects in fish. Can. J. Fish. Aquat. Sci. 1999, 56, 814–822. [Google Scholar] [CrossRef]
- Smolders, R.; De Boeck, G.; Blust, R. Changes in cellular energy budget as a measure of whole effluent toxicity in zebrafish (Danio rerio). Environ. Toxicol. Chem. 2003, 22, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Calow, P. Physiological costs of combating chemical toxicants: Ecological implications. Comp. Biochem. Physiol. Part C Comp. Pharmacol. Toxicol. 1991, 100, 3–6. [Google Scholar] [CrossRef]
- Witeska, M.; Sarnowski, P.; Ługowska, K.; Kowal, E. The effects of cadmium and copper on embryonic and larval development of ide Leuciscus idus L. Fish Physiol. Biochem. 2014, 40, 151–163. [Google Scholar] [CrossRef]
- Smolders, R.; Bervoets, L.; de Boeck, G.; Blust, R. Integrated condition indices as a measure of whole effluent toxicity in zebrafish (Danio rerio). Environ. Toxicol. Chem. 2002, 21, 87–93. [Google Scholar] [CrossRef]
- Stouthart, X.J.H.X.; Haans, J.L.M.; Lock, R.A.C.; Bonga, S.E.W. Effects of water pH on copper toxicity to early life stages of the common carp (Cyprinus carpio). Environ. Toxicol. Chem. 1996, 15, 376–383. [Google Scholar] [CrossRef]
- Sarnowski, P.; Witeska, M. The effects of Copper and Cadmium in single exposure or Co-exposure on growth of common carp (Cyprinus Carpio L.) Larvae. Polish J. Environ. Stud. 2008, 17, 791–796. [Google Scholar]
- Johnson, A.; Carew, E.; Sloman, K.A. The effects of copper on the morphological and functional development of zebrafish embryos. Aquat. Toxicol. 2007, 84, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Kienle, C.; Köhler, H.R.; Gerhardt, A. Behavioural and developmental toxicity of chlorpyrifos and nickel chloride to zebrafish (Danio rerio) embryos and larvae. Ecotoxicol. Environ. Saf. 2009, 72, 1740–1747. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.-J.; Jia, Y.-F.; Chen, N.; Bian, W.-P.; Li, Q.-K.; Ma, Y.-B.; Chen, Y.-L.; Pei, D.-S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Raible, D.W.; Kruse, G.J. Organization of the lateral line system in embryonic zebrafish. J. Comp. Neurol. 2000, 421, 189–198. [Google Scholar] [CrossRef]
- Froehlicher, M.; Liedtke, A.; Groh, K.J.; Neuhauss, S.C.F.; Segner, H.; Eggen, R.I.L. Zebrafish (Danio rerio) neuromast: Promising biological endpoint linking developmental and toxicological studies. Aquat. Toxicol. 2009, 95, 307–319. [Google Scholar] [CrossRef]
- Padilla, S.; Hunter, D.L.; Padnos, B.; Frady, S.; MacPhail, R.C. Assessing locomotor activity in larval zebrafish: Influence of extrinsic and intrinsic variables. Neurotoxicol. Teratol. 2011, 33, 624–630. [Google Scholar] [CrossRef]
- De Esch, C.; Van Der Linde, H.; Slieker, R.; Willemsen, R.; Wolterbeek, A.; Woutersen, R.; De Groot, D. Locomotor activity assay in zebrafish larvae: Influence of age, strain and ethanol. Neurotoxicol. Teratol. 2012, 34, 425–433. [Google Scholar] [CrossRef]
- MacPhail, R.C.; Brooks, J.; Hunter, D.L.; Padnos, B.; Irons, T.D.; Padilla, S. Locomotion in larval zebrafish: Influence of time of day, lighting and ethanol. Neurotoxicology 2009, 30, 52–58. [Google Scholar] [CrossRef]
- Weis, J.S.; Smith, G.; Zhou, T.; Santiago-Bass, C.; Weis, P. Effects of Contaminants on Behavior: Biochemical Mechanisms and Ecological Consequences. Bioscience 2001, 51, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Eales, J.G.; Brown, S.B. Measurement and regulation of thyroidal status in teleost fish. Rev. Fish Biol. Fish. 1993, 3, 299–347. [Google Scholar] [CrossRef]
- Shi, X.; Liu, C.; Wu, G.; Zhou, B. Waterborne exposure to PFOS causes disruption of the hypothalamus-pituitary-thyroid axis in zebrafish larvae. Chemosphere 2009, 77, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Picard-Aitken, M.; Fournier, H.; Pariseau, R.; Marcogliese, D.J.; Cyr, D.G. Thyroid disruption in walleye (Sander vitreus) exposed to environmental contaminants: Cloning and use of iodothyronine deiodinases as molecular biomarkers. Aquat. Toxicol. 2007, 83, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Long, M.D.; Fehlner, K.S.; Dyer, R.S. Sulfolane-induced hypothermia enhances survivability in mice. Environ. Res. 1986, 40, 92–97. [Google Scholar] [CrossRef]
- Best, C.; Vijayan, M.M. Cortisol elevation post-hatch affects behavioural performance in zebrafish larvae. Gen. Comp. Endocrinol. 2018, 257, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Faught, E.; Best, C.; Vijayan, M.M. Maternal stress-associated cortisol stimulation may protect embryos from cortisol excess in zebrafish. R. Soc. 2016, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schotland, J.; Shupliakov, O.; Wikström, M.; Brodin, L.; Srinivasan, M.; You, Z.B.; Herrera-Marschitz, M.; Zhang, W.; Hökfelt, T.; Grillner, S.; et al. Control of lamprey locomotor neurons by colocalized monoamine transmitters. Nature 1995, 374, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-H.; Wu, C.-H.; Chen, Y.-C.; Chow, W.-Y. Embryonic expression of zebrafish AMPA receptor genes: Zygotic gria2alpha expression initiates at the midblastula transition. Brain Res. 2006, 1110, 46–54. [Google Scholar] [CrossRef]
- Spence, R.; Gerlach, G.; Lawrence, C.; Smith, C. The behaviour and ecology of the zebrafish, Danio rerio. Biol. Rev. 2008, 83, 13–34. [Google Scholar] [CrossRef]
- Bambino, K.; Chu, J. Zebrafish in Toxicology and Environmental Health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar]
- Schneider, A.J.; Branam, A.M.; Peterson, R.E. Intersection of AHR and Wnt signaling in development, health, and disease. Int. J. Mol. Sci. 2014, 15, 17852–17885. [Google Scholar] [CrossRef]
- Teraoka, H.; Dong, W.; Tsujimoto, Y.; Iwasa, H.; Endoh, D.; Ueno, N.; Stegeman, J.J.; Peterson, R.E.; Hiraga, T. Induction of cytochrome P450 1A is required for circulation failure and edema by 2,3,7,8-tetrachlorodibenzo-p-dioxin in zebrafish. Biochem. Biophys. Res. Commun. 2003, 304, 223–228. [Google Scholar] [CrossRef]
- Liu, H.; Nie, F.H.; Lin, H.Y.; Ma, Y.; Ju, X.H.; Chen, J.J. Gooneratne RDevelopmental toxicity, EROD, and CYP1A mRNA expression in zebrafish embryos exposed to dioxin-like PCB126. Environ. Toxicol. 2016, 31, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, I.; Nazir, A.; Saxena, D.K.; Kar Chowdhuri, D. Heat shock response: hsp70 in environmental monitoring. J. Biochem. Mol. Toxicol. 2003, 17, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Beere, H.M.; Green, D.R. Stress management—Heat shock protein-70 and the regulation of apoptosis. Trends Cell Biol. 2001, 11, 6–10. [Google Scholar] [CrossRef]
- Krone, P.H.; Sass, J.B.; Lele, Z. Heat shock protein gene expression during embryonic development of the zebrafish. Cell. Mol. Life Sci. 1997, 53, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yabu, T.; Ojima, N. Stress Protein HSP70 in Fish. Aqua-BioSci. Monogr. 2010, 3, 111–141. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Koziol, C.; Dapper, J.; Kurelec, B.; Batel, R.; Rinkevich, B. Combinatory effects of temperature stress and nonionic organic pollutants on stress protein (hsp70) gene expression in the freshwater sponge Ephydatia fluviatilis. Environ. Toxicol. Chem. 1995, 14, 1203–1208. [Google Scholar]
- Hagedorn, M.; Kleinhans, F.W.; Artemov, D.; Pilatus, U. Characterization of a major permeability barrier in the zebrafish embryo. Biol. Reprod. 1998, 59, 1240–1250. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Horng, J.L.; Chao, P.L.; Chen, P.Y.; Shih, T.H.; Lin, L.Y. Aquaporin 1 is involved in acid secretion by ionocytes of zebrafish embryos through facilitating CO2 transport. PLoS ONE 2015, 10, 0136440. [Google Scholar] [CrossRef]
- Giffard-Mena, I.; Boulo, V.; Aujoulat, F.; Fowden, H.; Castille, R.; Charmantier, G.; Cramb, G. Aquaporin molecular characterization in the sea-bass (Dicentrarchus labrax): The effect of salinity on AQP1 and AQP3 expression. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Little, E.E.; Archeski, R.D.; Flerov, B.A.; Kozlovskaya, V.I. Behavioral indicators of sublethal toxicity in rainbow trout. Arch. Environ. Contam. Toxicol. 1990, 19, 380–385. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nominal Concentration (µg/mL) | Measured Concentration at 120 hpf (µg/mL) |
|---|---|
| 0 | 0 ± 0 |
| 50 | 49.90 ± 1.21 |
| 500 | 503.39 ± 4.13 |
| 5000 | 4950.36 ± 25.98 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, S.M.; Wahba, M.; Yu, L.; Achari, G.; Habibi, H.R. Health Impact Assessment of Sulfolane on Embryonic Development of Zebrafish (Danio rerio). Toxics 2019, 7, 42. https://doi.org/10.3390/toxics7030042
Shah SM, Wahba M, Yu L, Achari G, Habibi HR. Health Impact Assessment of Sulfolane on Embryonic Development of Zebrafish (Danio rerio). Toxics. 2019; 7(3):42. https://doi.org/10.3390/toxics7030042
Chicago/Turabian StyleShah, Soham M., Michael Wahba, Linlong Yu, Gopal Achari, and Hamid R. Habibi. 2019. "Health Impact Assessment of Sulfolane on Embryonic Development of Zebrafish (Danio rerio)" Toxics 7, no. 3: 42. https://doi.org/10.3390/toxics7030042
APA StyleShah, S. M., Wahba, M., Yu, L., Achari, G., & Habibi, H. R. (2019). Health Impact Assessment of Sulfolane on Embryonic Development of Zebrafish (Danio rerio). Toxics, 7(3), 42. https://doi.org/10.3390/toxics7030042