Toxicity of Aqueous L-Selenomethionine and Tert-Butyl Hydroperoxide Exposure to Zebrafish (Danio rerio) Embryos Following Tert-Butyl Hydroquinone Treatment



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Zebrafish Embryos
2.3. Aqueous Selenomethionine Dose-Response Embryo Exposures
2.4. Aqueous Selenomethionine, Pro- and Antioxidant Embryo Exposures
2.5. Deformities Analysis
2.6. Water and Tissue Concentrations
2.7. Real-Time Quantitative PCR (RT-qPCR)
2.8. Statistics
3. Results
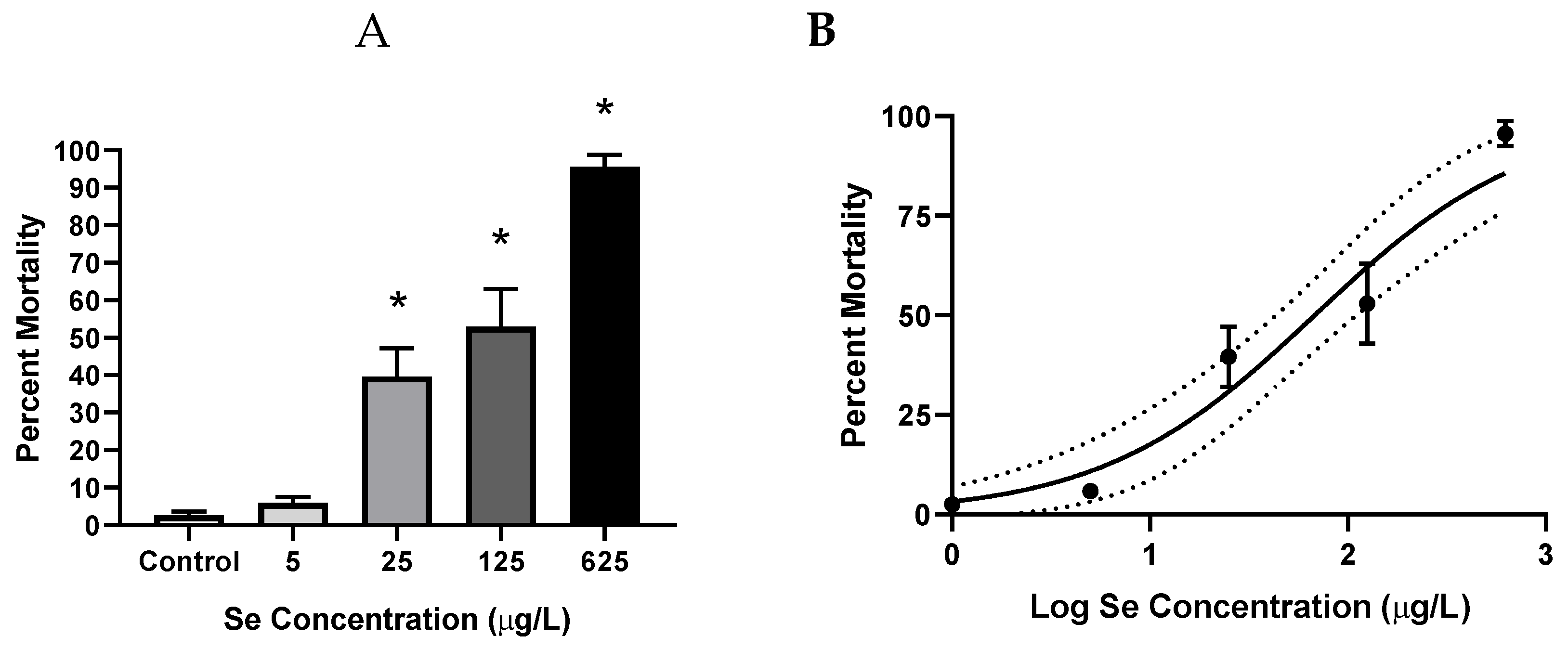
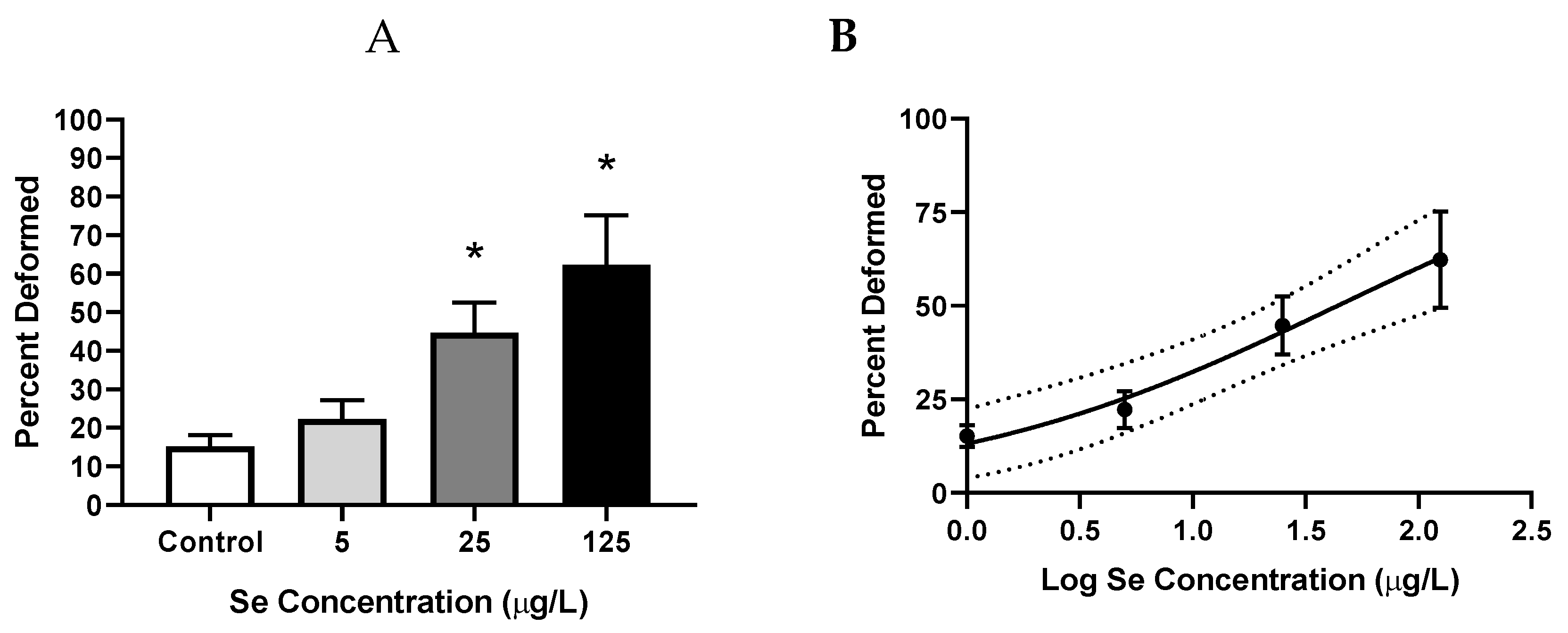
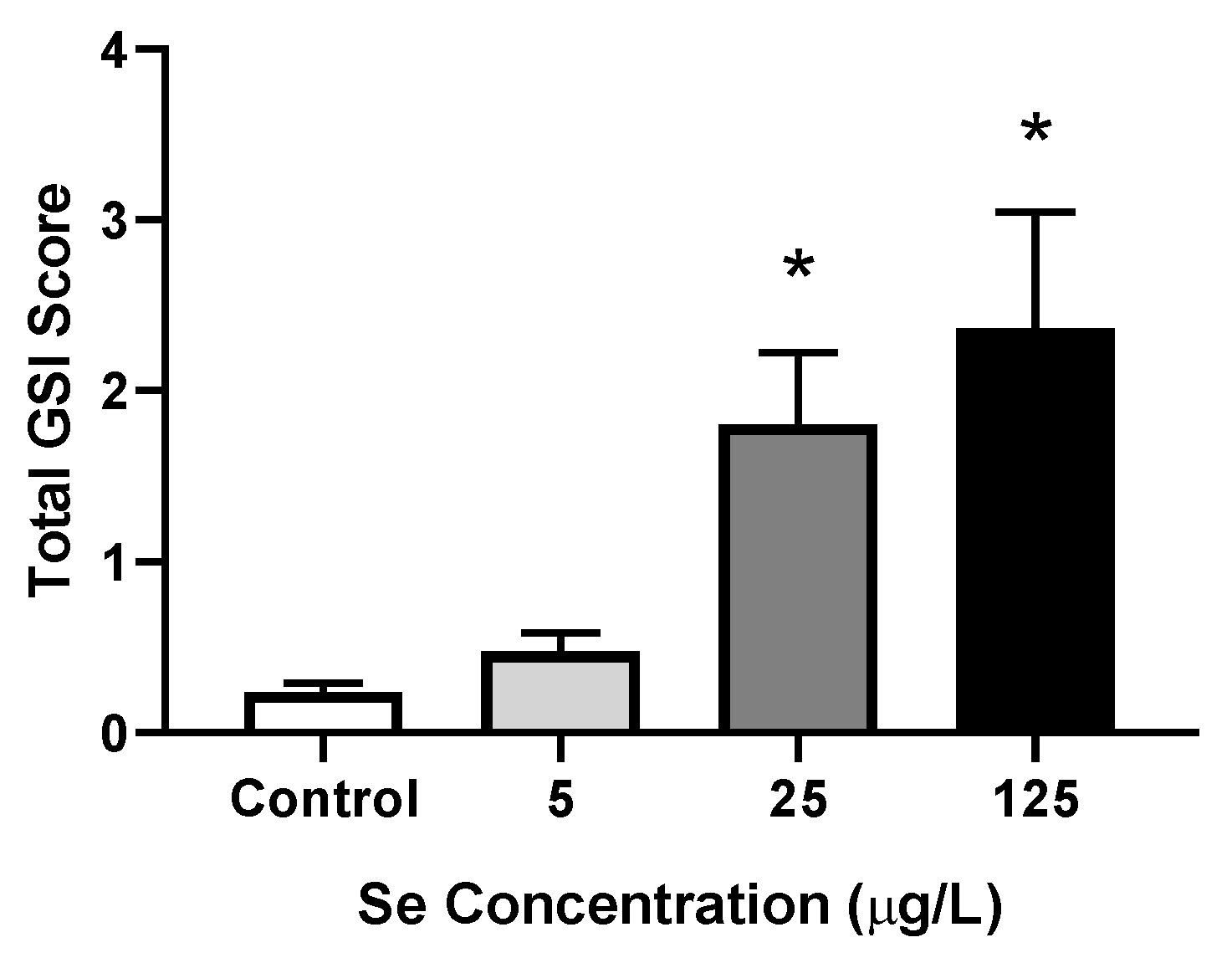
3.1. Aqueous Selenomethionine Dose-Response Embryo Exposures
3.1.1. Tissue Concentrations
3.1.2. Mortality
3.1.3. Frequency of Deformities
3.1.4. Severity of Deformities
3.1.5. Hatchability and Time to Hatch
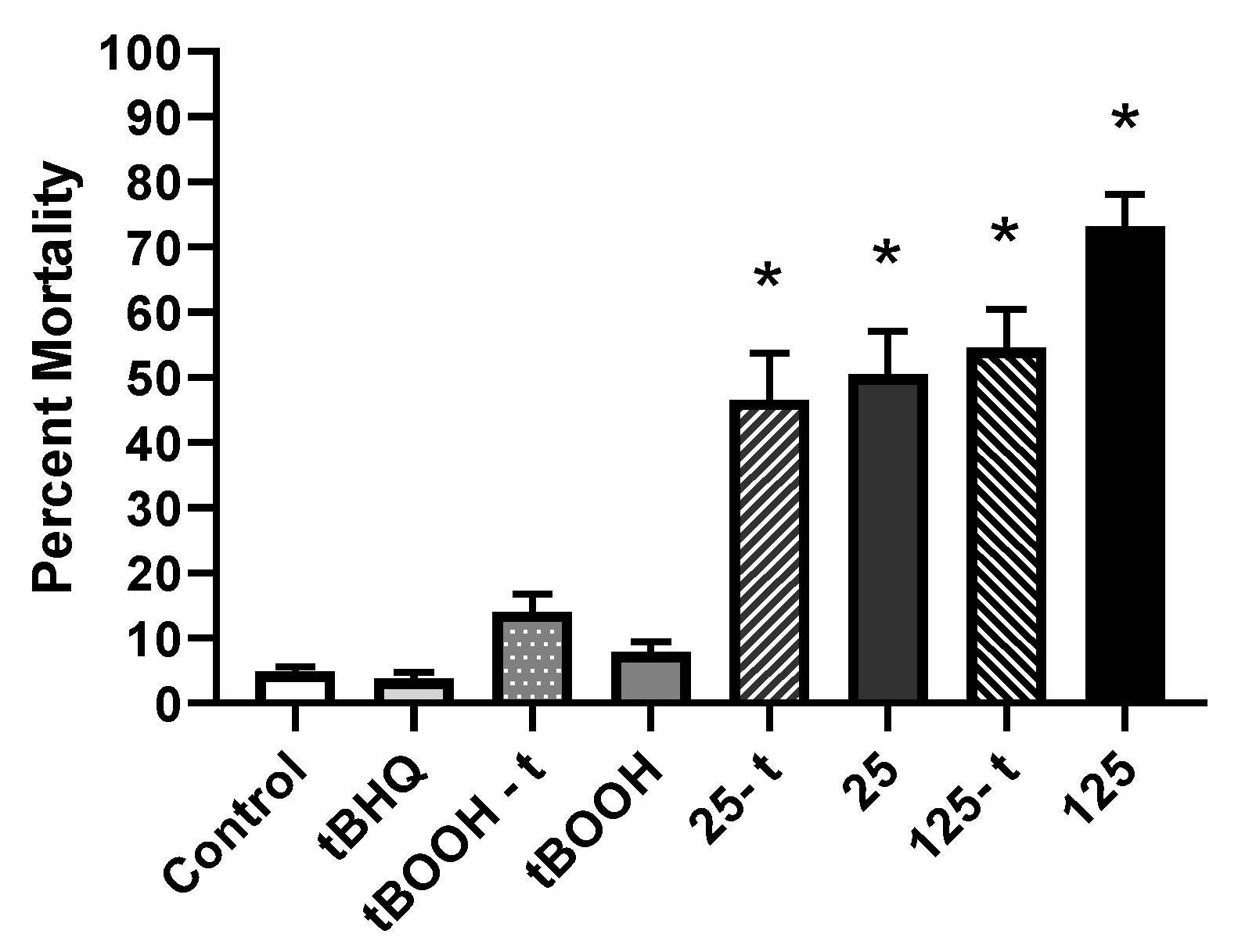
3.2. Aqueous Selenomethionine, Pro- and Antioxidant Embryo Exposures
3.2.1. Mortality
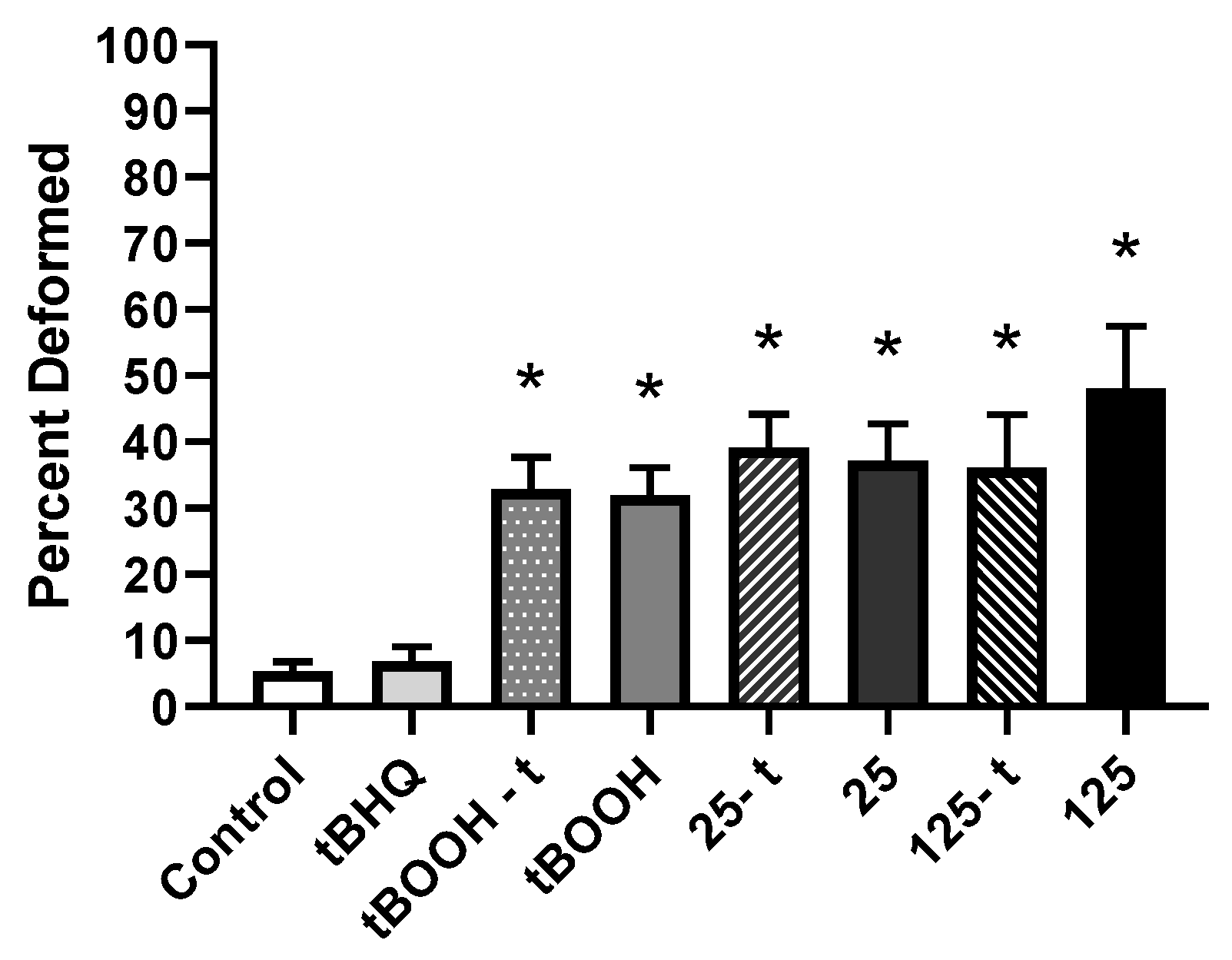
3.2.2. Frequency of Deformities
3.2.3. Severity of Deformities
3.2.4. Hatchability and Time to Hatch
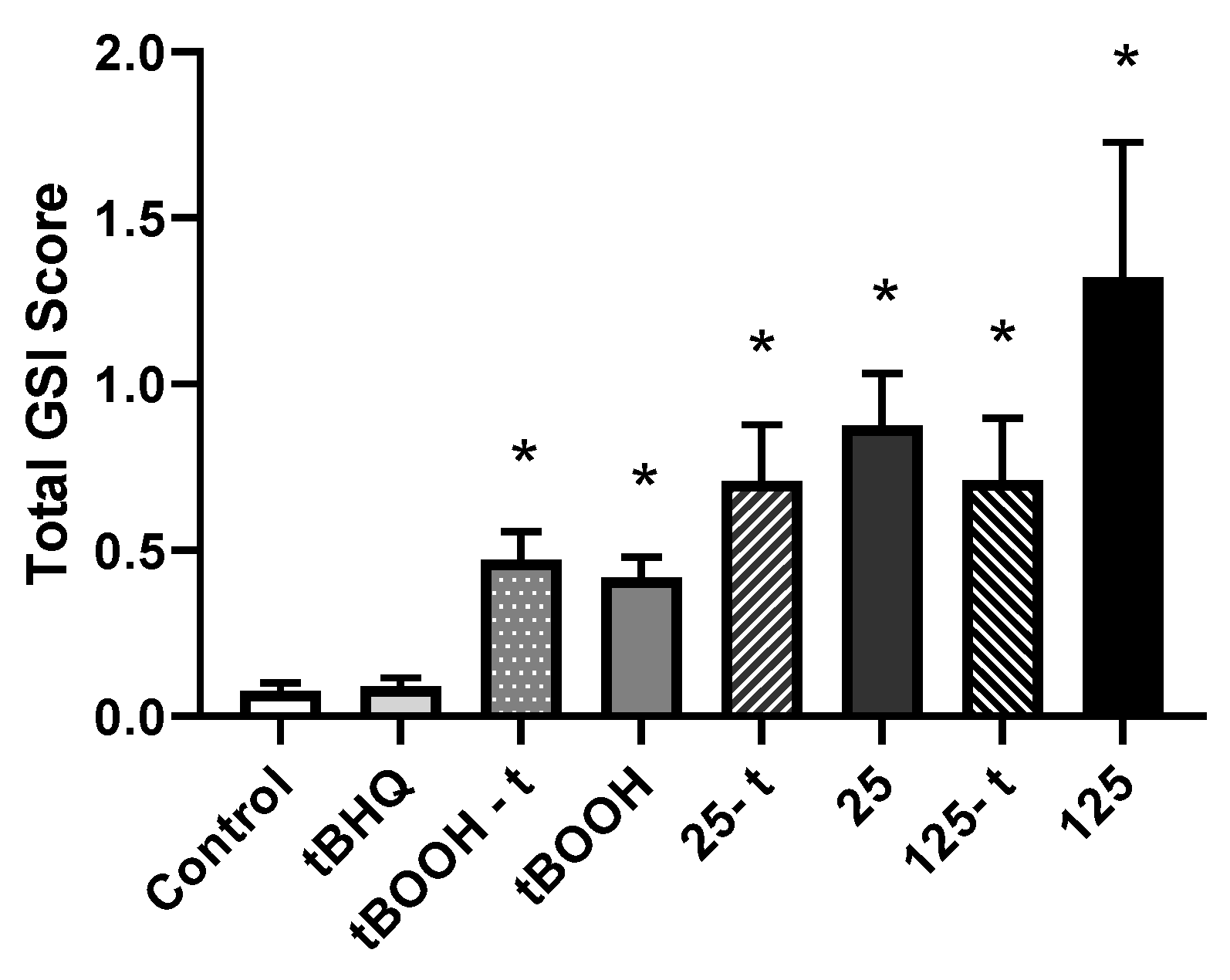
3.2.5. Gene Expression
4. Discussion
4.1. Aqueous Selenomethionine Dose-Response Embryo Exposures
4.1.1. Total Selenium in Stock Solutions and Tissue
4.1.2. Mortality
4.1.3. Frequency and Severity of Deformities
4.1.4. Hatchability and Time to Hatch
4.2. Aqueous Selenomethionine, Pro- and Antioxidant Embryo Exposures
4.2.1. Mortality
4.2.2. Frequency and Severity of Deformities
4.2.3. Hatchability and Time to Hatch
4.2.4. Gene Expression
5. Conclusions
- tBHQ is not ideally suited for use as an antioxidant. As noted by Kobayashi et al. [16], tBHQ is metabolized to an electrophilic quinone in cells, suggesting it could cause oxidative stress itself. Furthermore, it has been referenced to as a weak pro-oxidant in previous work studying oxidative stress [30].
- The concentration of tBHQ (100 µg/L) selected was low enough that no significant differences in mortality or deformities were observed relative to controls, but perhaps not high enough to induce the activation of the antioxidant defense system.
- The observed results were influenced by the duration and developmental stage of exposure. In the present study, embryos were pre-treated for 4h with 100 µg/L tBHQ immediately following collection. Previous work by Timme-Laragy et al. [17] treated embryos (at 48 or 72 hpf) with tBHQ for 4h. Another study conducted a 6h tBHQ treatment in 7-day-old zebrafish larvae [16], while Arnold et al. [12] pretreated zebrafish embryos (within 2 hpf) for 24h with the antioxidant N-acetylcysteine (NAC). Different exposure regimes will likely result in different responses.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ankley, G.T.; Johnson, R.D. Small Fish Models for Identifying and Assessing the Effects of Endocrine-disrupting Chemicals. ILAR J. 2004, 45, 469–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, T.F.; Finley, K.; Adams, W.J.; Besser, J.; Hopkins, W.D.; Jolley, D.; McNaughton, E.; Presser, T.S.; Shaw, D.P.; Unrine, J. What You Need to Know About Selenium. In Ecological Assessment of Selenium in the Aquatic Environment; Chapman, P.M., Adams, W.J., Brooks, M.L., Delos, C.G., Luoma, S.N., Maher, W.A., Ohlendorf, H.M., Presser, T.S., Shaw, D.P., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 7–45. [Google Scholar]
- Janz, D.M. Selenium. In Fish Physiology: Homeostasis and Toxicology of Essential Metals; Wood, C.M., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: San Diego, CA, USA, 2012; Volume 31, pp. 327–373. [Google Scholar]
- Lemly, D.A. A teratogenic deformity index for evaluating impacts of selenium on fish populations. Ecotoxicol. Environ. Saf. 1997, 37, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Lemly, D.A. Symptoms and implications of selenium toxicity in fish: The Belews Lake case example. Aquat. Toxicol. 2002, 57, 39–49. [Google Scholar] [CrossRef]
- Janz, D.M.; DeForest, D.K.; Brooks, M.L.; Chapman, P.M.; Gilron, G.; Hoff, D.; Hopkins, W.D.; McIntyre, D.O.; Mebane, C.A.; Palace, V.P.; et al. Selenium Toxicity to Aquatic Organisms. In Ecological Assessment of Selenium in the Aquatic Environment; Chapman, P.M., Adams, W.J., Brooks, M.L., Delos, C.G., Luoma, S.N., Maher, W.A., Ohlendorf, H.M., Presser, T.S., Shaw, D.P., Eds.; CRC Press: Pensacola, FL, USA, 2010; pp. 141–231. [Google Scholar]
- Spallholz, J.E. On the Nature of Selenium Toxicity and Carcinostatic Activity. Free Radic. Biol. Med. 1994, 17, 45–64. [Google Scholar] [CrossRef]
- Spallholz, J.E.; Shriver, B.J.; Reid, T.W. Dimethyldiselenide and Methylseleninic Acid Generate Superoxide in an In Vitro Chemiluminescence Assay in the Presence of Glutathione: Implications for the Anticarcinogenic Activity of L-Selenomethionine and L-Se Methylselenocysteine. Nutr. Cancer 2001, 40, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Palace, V.P.; Spallholz, J.E.; Holm, J.; Wautier, K.; Evans, R.E.; Baron, C.L. Metabolism of selenomethionine by rainbow trout (Oncorhynchus mykiss) embryos can generate oxidative stress. Ecotoxicol. Environ. Saf. 2004, 58, 17–21. [Google Scholar] [CrossRef]
- Thomas, J.K.; Janz, D.M. Embryo Microinjection of Selenomethionine Reduces Hatchability and Modifies Oxidant Responsive Gene Expression in Zebrafish. Sci. Rep. 2016, 6, 26520. [Google Scholar] [CrossRef]
- Lavado, R.; Shi, D.; Schlenk, D. Effects of salinity on the toxicity and biotransformation of l-selenomethionine in Japanese medaka (Oryzias latipes) embryos: Mechanisms of oxidative stress. Aquat. Toxicol. 2012, 108, 18–22. [Google Scholar] [CrossRef]
- Arnold, M.C.; Forte, J.E.; Osterberg, J.S.; Di Giulio, R.T. Antioxidant Rescue of Selenomethionine-Induced Teratogenesis in Zebrafish Embryos. Arch. Environ. Contam. Toxicol. 2016, 70, 311–320. [Google Scholar] [CrossRef]
- Kupsco, A.; Schlenk, D. Molecular mechanisms of selenium-Induced spinal deformities in fish. Aquat. Toxicol. 2016, 179, 143–150. [Google Scholar] [CrossRef]
- Kupsco, A.; Schlenk, D. Stage susceptibility of Japanese medaka (Oryzias latipes) to selenomethionine and hypersaline developmental toxicity. Environ. Toxicol. Chem. 2016, 35, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Gerhart, A.K.; Hecker, M.; Janz, D.M. Toxicity of Aqueous l-Selenomethionine Exposure to Early Life-Stages of the Fathead Minnow (Pimephales promelas). Bull. Environ. Contam. Toxicol. 2019, 102, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Itoh, K.; Suzuki, T.; Osanai, H.; Nishikawa, K.; Katoh, Y.; Takagi, Y.; Yamamoto, M. Identification of the interactive interface and phylogenic conservation of the Nrf2-Keap1 system. Genes Cells 2002, 7, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Timme-Laragy, A.R.; Karchner, S.I.; Franks, D.G.; Jenny, M.J.; Harbeitner, R.C.; Goldstone, J.V.; McArthur, A.G.; Hahn, M.E. Nrf2b, Novel Zebrafish Paralog of Oxidant-responsive Transcription Factor NF-E2-related Factor 2 (NRF2). J. Biol. Chem. 2012, 287, 4609–4627. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Holm, J.; Palace, V.P.; Wautier, K.; Evans, R.E.; Baron, C.L.; Podemski, C.; Siwik, P.; Sterling, G. An assessment of the development and survival of wild rainbow trout (Oncorhynchus mykiss) and brook trout (Salvelinus fontinalis) exposed to elevated selenium in an area of active coal mining. In The Big Fish Bang, Proceedings of the 26th Annual Larval Fish Conference, Os, Norway, 22–26 July 2002; Browman, H.I., Skiftesvik, A.B., Eds.; Institute of Marine Research: Bergen, Norway, 2003; pp. 257–273. [Google Scholar]
- Liu, C.; Xu, H.; Lam, S.H.; Gong, Z. Selection of Reliable Biomarkers from PCR Array Analyses Using Relative Distance Computational Model: Methodology and Proof-of-Concept Study. PLoS ONE 2013, 8, e83954. [Google Scholar] [CrossRef] [PubMed]
- Tiedke, J.; Cubuk, C.; Burmester, T. Environmental acidification triggers oxidative stress and enhances globin expression in zebrafish gills. Biochem. Biophys. Res. Commun. 2013, 441, 624–629. [Google Scholar] [CrossRef] [PubMed]
- cms_042380—Guide to Performing Relative Quantitation of Gene Expression Using RT qPCR. ThermoFisher Scientific. Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https%3A%2F%2Fassets.thermofisher.com%2FTFS-Assets%2FLSG%2Fmanuals%2Fcms_042380.pdf&title=R3VpZGUgdG8gUGVyZm9ybWluZyBSZWxhdGl2ZSBRdWFudGl0YXRpb24gb2YgR2VuZSBFeHByZXNzaW9uIFVzaW5nIFJlYWwtVGltZSBRdWFudGl0YXRpdmUgUENS (accessed on 5 June 2019).
- Muscatello, J.R.; Bennett, P.M.; Himbeault, K.T.; Belknap, A.M.; Janz, D.M. Larval Deformities Associated with Selenium Accumulation in Northern Pike (Esox lucius) Exposed to Metal Mining Effluent. Environ. Sci. Technol. 2006, 40, 6506–6512. [Google Scholar] [CrossRef]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The antioxidant role of selenium and seleno-compounds. Biomed. Pharmacother. 2003, 57, 134–144. [Google Scholar] [CrossRef]
- Oakley, A. Glutathione transferases: A structural perspective. Drug Metab. Rev. 2011, 43, 138–151. [Google Scholar] [CrossRef]
- Michiels, C.; Raes, M.; Toussaint, O.; Remacle, J. Importance of SE-glutathione peroxidase, catalase, and CU/ZN-SOD for cell survival against oxidative stress. Free Radic. Biol. Med. 1994, 17, 235–248. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Dismutases. J. Biol. Chem. 1989, 264, 7761–7764. [Google Scholar] [PubMed]
- Kawajiri, K.; Fujii-Kuriyama, Y. The aryl hydrocarbon receptor: A multifunctional chemical sensor for host defense and homeostatic maintenance. Exp. Anim. 2017, 66, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.E.; McArthur, A.G.; Karchner, S.I.; Franks, D.G.; Jenny, M.J.; Timme-Laragy, A.R.; Stegeman, J.J.; Woodin, B.R.; Cipriano, M.J.; Linney, E. The Transcriptional Response to Oxidative Stress during Vertebrate Development: Effects of tert- Butylhydroquinone and 2,3,7,8-Tetrachlorodibenzo-p-Dioxin. PLoS ONE 2014, 9, e113158. [Google Scholar] [CrossRef] [PubMed]
- Sant, K.E.; Hansen, J.M.; Williams, L.M.; Tran, N.L.; Goldstone, J.V.; Stegeman, J.J.; Hahn, M.E.; Timme-Laragy, A. The role of Nrf1 and Nrf2 in the regulation of glutathione and redox dynamics in the developing zebrafish embryo. Redox Biol. 2017, 13, 207–218. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nominal Concentration (µg/L) | Measured Concentration (µg/L) | Tissue Concentration (µg/g dry mass) |
|---|---|---|
| Control | 0.351 ± 0.026 | 2.98 ± 0.073 |
| 5 | 5.13 ± 0.30 | 34.2 ± 3.5 |
| 25 | 24.8 ± 0.41 | 144 ± 26 |
| 125 | 124 ± 0.81 | 264 ± 38 |
| 625 | 615 ± 6.4 | 291 ± 152 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerhart, A.K.; Janz, D.M. Toxicity of Aqueous L-Selenomethionine and Tert-Butyl Hydroperoxide Exposure to Zebrafish (Danio rerio) Embryos Following Tert-Butyl Hydroquinone Treatment. Toxics 2019, 7, 44. https://doi.org/10.3390/toxics7030044
Gerhart AK, Janz DM. Toxicity of Aqueous L-Selenomethionine and Tert-Butyl Hydroperoxide Exposure to Zebrafish (Danio rerio) Embryos Following Tert-Butyl Hydroquinone Treatment. Toxics. 2019; 7(3):44. https://doi.org/10.3390/toxics7030044
Chicago/Turabian StyleGerhart, Allyson K., and David M. Janz. 2019. "Toxicity of Aqueous L-Selenomethionine and Tert-Butyl Hydroperoxide Exposure to Zebrafish (Danio rerio) Embryos Following Tert-Butyl Hydroquinone Treatment" Toxics 7, no. 3: 44. https://doi.org/10.3390/toxics7030044
APA StyleGerhart, A. K., & Janz, D. M. (2019). Toxicity of Aqueous L-Selenomethionine and Tert-Butyl Hydroperoxide Exposure to Zebrafish (Danio rerio) Embryos Following Tert-Butyl Hydroquinone Treatment. Toxics, 7(3), 44. https://doi.org/10.3390/toxics7030044