Combining Different In Vitro Bioassays to Evaluate Genotoxicity of Water-Accommodated Fractions from Petroleum Products

 , ,
, , 
Abstract
:1. Introduction
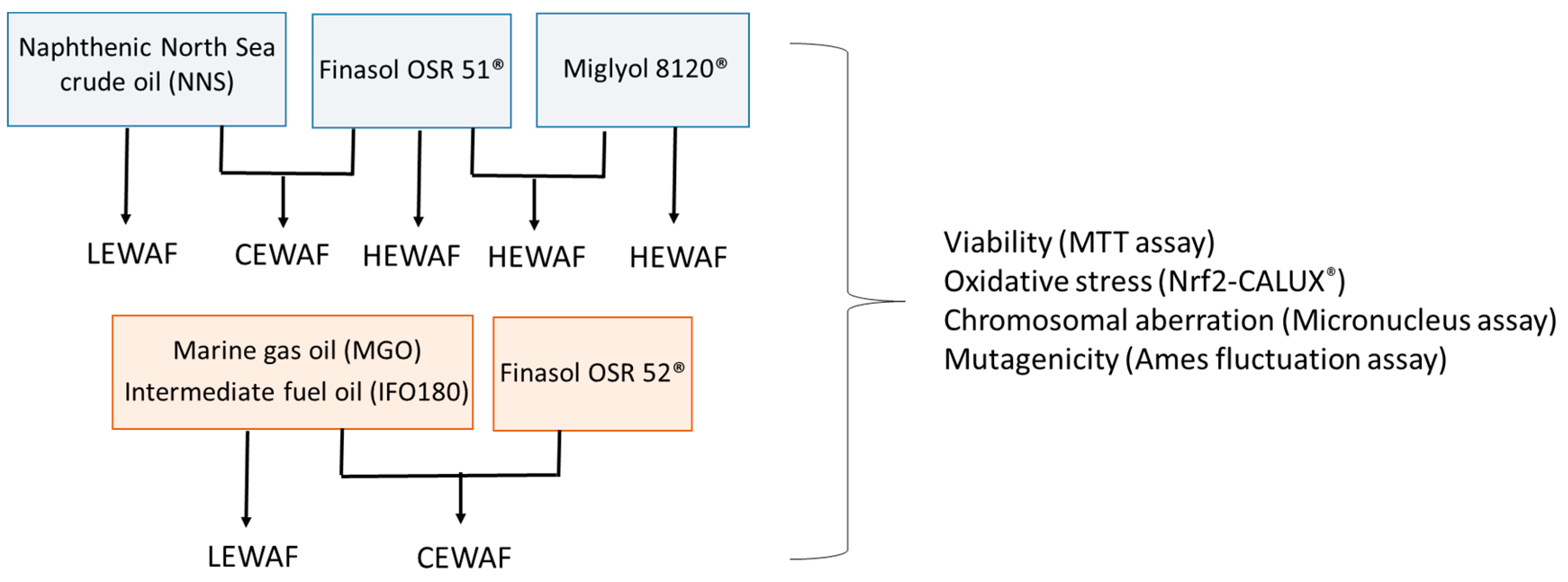
2. Materials and Methods
2.1. Sample Information
2.2. Preparation of Water-Accommodated Fractions (WAFs) for Cell Exposure
2.3. Chemical Analysis of Target PAHs in WAF Solutions
2.4. MTT Assay on Cell Viability
2.5. Nrf2-CALUX® Assay for Oxidative Stress
2.5.1. Human Osteosarcoma U2-OS Cell Culture
2.5.2. Assay Procedure
2.6. Micronucleus Assay on Chromosomal Aberration
2.6.1. Zebrafish Liver (ZF-L) Cell Culture
2.6.2. Assay Procedure
2.7. Ames Fluctuation Assay on Mutagenicity
2.7.1. Salmonella Typhimurium Bacterial Strain
2.7.2. Assay Procedure
3. Results
3.1. Target PAHs in LEWAF Stock Solutions
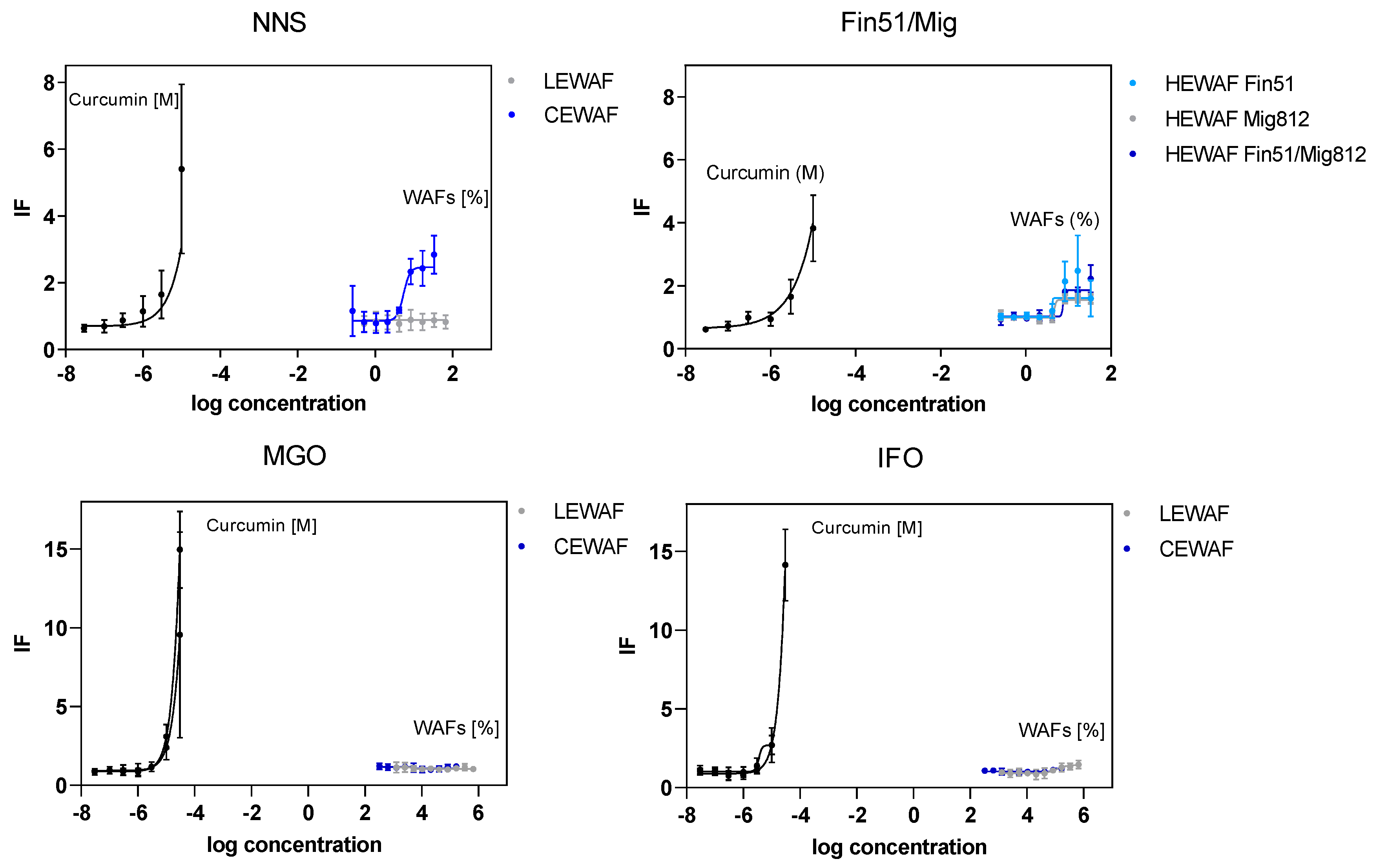
3.2. Oxidative Stress Response in U2-OS Cells Using the Nrf2-CALUX® Bioassay
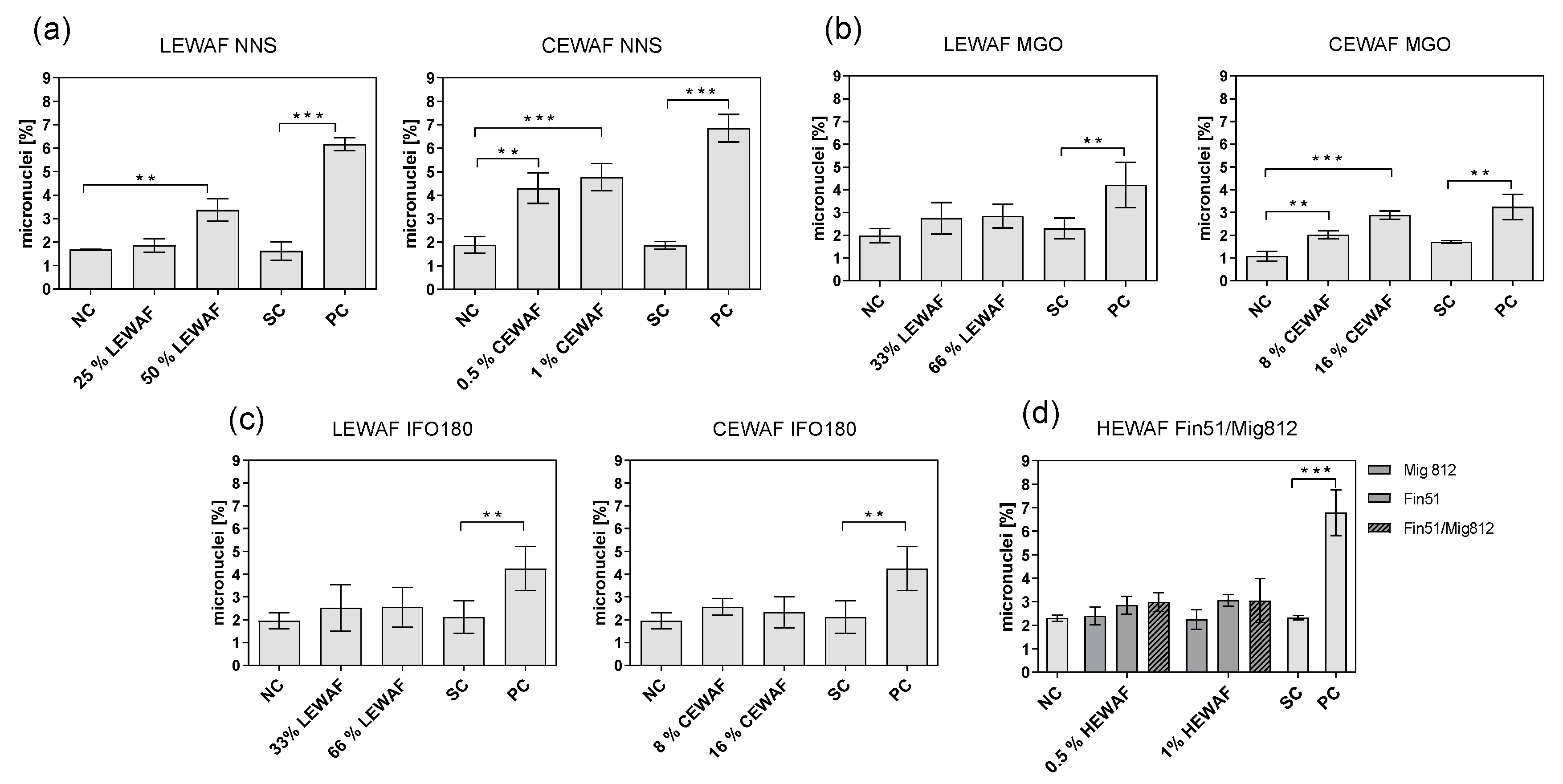
3.3. Chromosomal Aberrations in ZF-L Cells Using the Micronucleus Assay
3.4. Mutagenicity Using the Ames Fluctuation Assay
4. Discussion
4.1. Oxidative Stress
4.2. Genotoxicity
4.3. Comparison of Different Oil Types and the Influence of Chemical Dispersant Application
4.4. One Assay Is Not Enough: Combining In Vitro-Based Methods for Genotoxicity Assessment of Petroleum Products
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Babich, H.; Borenfreund, E. Cytotoxicity and genotoxicity assays with cultured fish cells: A review. Toxicol. In Vitro 1991, 5, 91–100. [Google Scholar] [CrossRef]
- Kirkland, D.; Aardema, M.; Henderson, L.; Müller, L. Evaluation of the ability of a battery of three in vitro genotoxicity tests to discriminate rodent carcinogens and non-carcinogens: I. Sensitivity, specificity and relative predictivity. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 584, 1–256. [Google Scholar] [CrossRef] [PubMed]
- Łuczyński, M.K.; Góra, M.; Brzuzan, P.; Wilamowski, J.; Kozik, B. Oxidative metabolism, mutagenic and carcinogenic properties of some polycyclic aromatic hydrocarbons. Environ. Biotechnol. 2005, 1, 16–28. [Google Scholar]
- Shimada, T. Xenobiotic-metabolizing enzymes involved in activation and detoxification of carcinogenic polycyclic aromatic hydrocarbons. Drug Metab. Pharmacokinet. 2006, 21, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Nigro, M.; Frenzilli, G.; Scarcelli, V.; Gorbi, S.; Regoli, F. Induction of DNA strand breakage and apoptosis in the eel Anguilla anguilla. Mar. Environ. Res. 2002, 54, 517–520. [Google Scholar] [CrossRef]
- Żelazna, K.; Rudnicka, K.; Tejs, S. In vitro micronucleus test assessment of polycyclic aromatic hydrocarbons. Environ. Biotechnol. 2011, 7, 70–80. [Google Scholar]
- Abdel-Massih, R.M.; Melki, P.N.; Afif, C.; Daoud, Z. Detection of genotoxicity in hospital wastewater of a developing country using SOS chromotest and Ames fluctuation test. J. Environ. Eng. Ecol. Sci. 2013, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Bickham, J.W.; Sandhu, S.; Hebert, P.D.; Chikhi, L.; Athwal, R. Effects of chemical contaminants on genetic diversity in natural populations: Implications for biomonitoring and ecotoxicology. Mutat. Res. Rev. Mutat. Res. 2000, 463, 33–51. [Google Scholar] [CrossRef]
- Regoli, F.; Gorbi, S.; Frenzilli, G.; Nigro, M.; Corsi, I.; Focardi, S.; Winston, G.W. Oxidative stress in ecotoxicology: From the analysis of individual antioxidants to a more integrated approach. Mar. Environ. Res. 2002, 54, 419–423. [Google Scholar] [CrossRef]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Bolognesi, C.; Perrone, E.; Roggieri, P.; Pampanin, D.M.; Sciutto, A. Assessment of micronuclei induction in peripheral erythrocytes of fish exposed to xenobiotics under controlled conditions. Aquat. Toxicol. 2006, 78, S93–S98. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cadahía, B.; Laffon, B.; Pásaro, E.; Méndez, J. Evaluation of PAH bioaccumulation and DNA damage in mussels (Mytilus galloprovincialis) exposed to spilled Prestige crude oil. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 138, 453–460. [Google Scholar] [CrossRef]
- Pilcher, W.; Miles, S.; Tang, S.; Mayer, G.; Whitehead, A. Genomic and genotoxic responses to controlled weathered-oil exposures confirm and extend field studies on impacts of the deepwater horizon oil spill on native killifish. PLoS ONE 2014, 9, e106351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolognesi, C.; Perrone, E.; Roggieri, P.; Sciutto, A. Bioindicators in monitoring long term genotoxic impact of oil spill: Haven case study. Mar. Environ. Res. 2006, 62, S287–S291. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tajes, J.; Rábade, T.; Laffon, B.; Méndez, J. Monitoring follow up of two areas affected by the Prestige oil four years after the spillage. J. Toxicol. Environ. Health Part A 2011, 74, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Pietrapiana, D.; Modena, M.; Guidetti, P.; Falugi, C.; Vacchi, M. Evaluating the genotoxic damage and hepatic tissue alterations in demersal fish species: A case study in the Ligurian Sea (NW-Mediterranean). Mar. Pollut. Bull. 2002, 44, 238–243. [Google Scholar] [CrossRef]
- Boettcher, M.; Grund, S.; Keiter, S.; Kosmehl, T.; Reifferscheid, G.; Seitz, N.; Rocha, P.S.; Hollert, H.; Braunbeck, T. Comparison of in vitro and in situ genotoxicity in the Danube River by means of the comet assay and the micronucleus test. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 700, 11–17. [Google Scholar] [CrossRef]
- Bolognesi, C.; Cirillo, S. Genotoxicity biomarkers in aquatic bioindicators. Curr. Zool. 2014, 60, 273–284. [Google Scholar] [CrossRef]
- Pellacani, C.; Buschini, A.; Furlini, M.; Poli, P.; Rossi, C. A battery of in vivo and in vitro tests useful for genotoxic pollutant detection in surface waters. Aquat. Toxicol. 2006, 77, 1–10. [Google Scholar] [CrossRef]
- Jennings, P. The future of in vitro toxicology. Toxicol. In Vitro 2015, 29, 1217–1221. [Google Scholar] [CrossRef]
- Araldi, R.P.; de Melo, T.C.; Mendes, T.B.; de Sá Júnior, P.L.; Nozima, B.H.N.; Ito, E.T.; de Carvalho, R.F.; de Souza, E.B.; de Cassia Stocco, R. Using the comet and micronucleus assays for genotoxicity studies: A review. Biomed. Pharmacother. 2015, 72, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Barfknecht, T.; Naismith, R. Methodology for evaluating the genotoxicity of hazardous environmental samples. Hazard. Waste 1984, 1, 93–109. [Google Scholar] [CrossRef]
- Brack, W.; Aissa, S.A.; Backhaus, T.; Dulio, V.; Escher, B.I.; Faust, M.; Hilscherova, K.; Hollender, J.; Hollert, H.; Müller, C. Effect-based methods are key. The European Collaborative Project SOLUTIONS recommends integrating effect-based methods for diagnosis and monitoring of water quality. Environ. Sci. Eur. 2019, 31, 1–6. [Google Scholar] [CrossRef]
- Brack, W.; Dulio, V.; Ågerstrand, M.; Allan, I.; Altenburger, R.; Brinkmann, M.; Bunke, D.; Burgess, R.M.; Cousins, I.; Escher, B.I. Towards the review of the European Union Water Framework Directive: Recommendations for more efficient assessment and management of chemical contamination in European surface water resources. Sci. Total Environ. 2017, 576, 720–737. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Chan, K.M. Differential effects of metal ions on TCDD-induced cytotoxicity and cytochrome P4501A1 gene expression in a zebrafish liver (ZFL) cell-line. Metallomics 2016, 8, 236–251. [Google Scholar] [CrossRef] [PubMed]
- Collodi, P.; Miranda, C.L.; Zhao, X.; Buhler, D.R.; Barnes, D.W. Induction of zebrafish (Brachydanio rerio) P450in vivoand in cell culture. Xenobiotica 1994, 24, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.E.; Lavado, R. Applicability of in vitro methods in evaluating the biotransformation of polycyclic aromatic hydrocarbons (PAHs) in fish: Advances and challenges. Sci. Total Environ. 2019, 671, 685–695. [Google Scholar] [CrossRef]
- Ames, B.N.; Lee, F.D.; Durston, W.E. An Improved Bacterial Test System for the Detection and Classification of Mutagens and Carcinogens. Proc. Natl. Acad. Sci. USA 1973, 70, 782–786. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- van der Linden, S.C.; von Bergh, A.R.M.; van Vught-Lussenburg, B.M.A.; Jonker, L.R.A.; Teunis, M.; Krul, C.A.M.; van der Burg, B. Development of a panel of high-throughput reporter-gene assays to detect genotoxicity and oxidative stress. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 760, 23–32. [Google Scholar] [CrossRef]
- Wakabayashi, N.; Slocum, S.L.; Skoko, J.J.; Shin, S.; Kensler, T.W. When NRF2 talks, who’s listening? Antioxid. Redox Signal. 2010, 13, 1649–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SDS sheet Finasol OSR 52. Safety Data Sheet According to Regulation (EC) No 1970/2006. SDS # 30034 2013. Available online: http://www.clearcoastllc.com/documents/Finasol%20OS%2052%20MSDS%20Total%20Spec.pdf (accessed on 25 June 2020).
- SDS sheet Finasol OSR 51. Material Safety Data Sheet According to Regulation (EC) No 1970/2006. SDS # 30033 2012. Available online: https://sdsinventory.com/substances/accessSDS/SDS-46002-58244a71c59605.23905294 (accessed on 25 June 2020).
- Jørgensen, K.S.; Kreutzer, A.; Lehtonen, K.K.; Kankaanpää, H.; Rytkönen, J.; Wegeberg, S.; Gustavson, K.; Fritt-Rasmussen, J.; Truu, J.; Kõuts, T. The EU Horizon 2020 project GRACE: Integrated oil spill response actions and environmental effects. Environ. Sci. Eur. 2019, 31, 44. [Google Scholar] [CrossRef]
- Johann, S.; Esser, M.; Nüßer, L.; Altin, D.; Hollert, H.; Seiler, T.-B. Receptor-mediated estrogenicity of native and chemically dispersed crude oil determined using adapted microscale reporter gene assays. Environ. Int. 2020, 134, 105320. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.M.; Aurand, D.; Bragin, G.E.; Clark, J.R.; Coelho, G.M.; Sowby, M.L.; Tjeerdema, R.S. Standardization of the Preparation and Quantitation of Water-accommodated Fractions of Petroleum for Toxicity Testing. Mar. Pollut. Bull. 2000, 40, 1007–1016. [Google Scholar] [CrossRef]
- Potter, D.W.; Pawliszyn, J. Rapid determination of polyaromatic hydrocarbons and polychlorinated biphenyls in water using solid-phase microextraction and GC/MS. Environ. Sci. Technol. 1994, 28, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.T.; Achten, C. Time to Say Goodbye to the 16 EPA PAHs? Toward an Up-to-Date Use of PACs for Environmental Purposes. Polycycl. Aromat. Compd. 2015, 35, 330–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Legler, J.; van den Brink, C.E.; Brouwer, A.; Murk, A.J.; van der Saag, P.T.; Vethaak, A.D.; van der Burg, B. Development of a stably transfected estrogen receptor-mediated luciferase reporter gene assay in the human T47D breast cancer cell line. Toxicol. Sci. Off. J. Soc. Toxicol. 1999, 48, 55–66. [Google Scholar] [CrossRef]
- Kuckelkorn, J.; Redelstein, R.; Heide, T.; Kunze, J.; Maletz, S.; Waldmann, P.; Grummt, T.; Seiler, T.B.; Hollert, H. A hierarchical testing strategy for micropollutants in drinking water regarding their potential endocrine-disrupting effects—towards health-related indicator values. Environ. Sci. Pollut. Res. 2017, 25, 4051–4065. [Google Scholar] [CrossRef]
- Steinberg, P.; Behnisch, P.A.; Besselink, H.; Brouwer, A.A. Screening of molecular cell targets for carcinogenic heterocyclic aromatic amines by using CALUX® reporter gene assays. Cell Biol. Toxicol. 2016, 33, 283–293. [Google Scholar] [CrossRef] [PubMed]
- ISO. ISO/CD 19040-3 Water Quality—Determination of the Estrogenic Potential of Water and Waste Water—Part 3: In Vitro Human Cell-Based Reporter Gene Assay (ISO/TC 147/SC 5N 864); ISO: Geneva, Switzerland, 2014. [Google Scholar]
- Ghosh, C.; Zhou, Y.L.; Collodi, P. Derivation and characterization of a zebrafish liver cell line. Cell Biol. Toxicol. 1994, 10, 167–176. [Google Scholar] [CrossRef] [PubMed]
- ISO. ISO/DIS 21427-2 Water Quality—Evaluation of the Genotoxicity by Measurements of the Induction of Micronuclei; ISO: Geneva, Switzerland, 2004. [Google Scholar]
- Bluhm, K.; Heger, S.; Redelstein, R.; Brendt, J.; Anders, N.; Mayer, P.; Schaeffer, A.; Hollert, H. Genotoxicity of three biofuel candidates compared to reference fuels. Environ. Toxicol. Pharmacol. 2018, 64, 131–138. [Google Scholar] [CrossRef] [PubMed]
- ISO. Water Quality—Determination of Genotoxicity of Water and Waste Water using the Salmonella/Microsome Fluctuation Test. Ames Fluctuation Test; ISO: Geneva, Switzerland, 2012. [Google Scholar]
- Reifferscheid, G.; Maes, H.M.; Allner, B.; Badurova, J.; Belkin, S.; Bluhm, K.; Brauer, F.; Bressling, J.; Domeneghetti, S.; Elad, T.; et al. International round-robin study on the Ames fluctuation test. Environ. Mol. Mutagen. 2012, 53, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Escher, B.I.; Aїt-Aїssa, S.; Behnisch, P.A.; Brack, W.; Brion, F.; Brouwer, A.; Buchinger, S.; Crawford, S.E.; Du Pasquier, D.; Hamers, T.; et al. Effect-based trigger values for in vitro and in vivo bioassays performed on surface water extracts supporting the environmental quality standards (EQS) of the European Water Framework Directive. Sci. Total Environ. 2018, 628–629, 748–765. [Google Scholar] [CrossRef]
- van der Oost, R.; Sileno, G.; Suárez-Muñoz, M.; Nguyen, M.T.; Besselink, H.; Brouwer, A. SIMONI (Smart Integrated Monitoring) as a novel bioanalytical strategy for water quality assessment: Part I-model design and effect-based trigger values. Environ. Toxicol. Chem. 2017, 36, 2385–2399. [Google Scholar] [CrossRef]
- Pieterse, B.; Rijk, I.J.C.; Simon, E.; van Vugt-Lussenburg, B.M.A.; Fokke, B.F.H.; van der Wijk, M.; Besselink, H.; Weber, R.; van der Burg, B. Effect-based assessment of persistent organic pollutant and pesticide dumpsite using mammalian CALUX reporter cell lines. Environ. Sci. Pollut. Res. 2015, 22, 14442–14454. [Google Scholar] [CrossRef]
- Shi, H.; Sui, Y.; Wang, X.; Luo, Y.; Ji, L. Hydroxyl radical production and oxidative damage induced by cadmium and naphthalene in liver of Carassius auratus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 140, 115–121. [Google Scholar] [CrossRef]
- Sun, Y.; Yu, H.; Zhang, J.; Yin, Y.; Shi, H.; Wang, X. Bioaccumulation, depuration and oxidative stress in fish Carassius auratus under phenanthrene exposure. Chemosphere 2006, 63, 1319–1327. [Google Scholar] [CrossRef]
- Farmen, E.; Harman, C.; Hylland, K.; Tollefsen, K.-E. Produced water extracts from North Sea oil production platforms result in cellular oxidative stress in a rainbow trout in vitro bioassay. Mar. Pollut. Bull. 2010, 60, 1092–1098. [Google Scholar] [CrossRef]
- Yilmaz, B.; Ssempebwa, J.; Mackerer, C.R.; Arcaro, K.F.; Carpenter, D.O. Effects of Polycyclic Aromatic Hydrocarbon-Containing Oil Mixtures on Generation of Reactive Oxygen Species and Cell Viability in MCF-7 Breast Cancer Cells. J. Toxicol. Environ. Health Part A 2007, 70, 1108–1115. [Google Scholar] [CrossRef]
- Achuba, F.I.; Osakwe, S.A. Petroleum – induced free radical toxicity in African catfish (Clarias gariepinus). Fish Physiol. Biochem. 2003, 29, 97–103. [Google Scholar] [CrossRef]
- Silva, C.A.; Ribeiro, C.O.; Katsumiti, A.; Araújo, M.L.P.; Zandoná, E.M.; Silva, G.C.; Maschio, J.; Roche, H.; de Assis, H.S. Evaluation of waterborne exposure to oil spill 5 years after an accident in Southern Brazil. Ecotoxicol. Environ. Saf. 2009, 72, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Wang, X.R.; Guo, H.Y.; Wu, J.C.; Xue, Y.Q. Effects of water-soluble fractions of diesel oil on the antioxidant defenses of the goldfish, Carassius auratus. Ecotoxicol. Environ. Saf. 2004, 58, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Mollergues, J.; van Vugt-Lussenburg, B.; Kirchnawy, C.; Bandi, R.A.; van der Lee, R.B.; Marin-Kuan, M.; Schilter, B.; Fussell, K.C. Incorporation of a metabolizing system in biodetection assays for endocrine active substances. ALTEX Altern. Anim. Exp. 2017, 34, 389–398. [Google Scholar]
- Lachner, D.; Oliveira, L.F.; Martinez, C.B.R. Effects of the water soluble fraction of gasoline on ZFL cell line: Cytotoxicity, genotoxicity and oxidative stress. Toxicol. In Vitro 2015, 30, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Kammann, U.; Biselli, S.; Hühnerfuss, H.; Reineke, N.; Theobald, N.; Vobach, M.; Wosniok, W. Genotoxic and teratogenic potential of marine sediment extracts investigated with comet assay and zebrafish test. Environ. Pollut. 2004, 132, 279–287. [Google Scholar] [CrossRef]
- Grant, G.M.; Jackman, S.M.; Kolanko, C.J.; Stenger, D.A. JP-8 jet fuel-induced DNA damage in H4IIE rat hepatoma cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2001, 490, 67–75. [Google Scholar] [CrossRef]
- Leme, D.M.; Grummt, T.; de Oliveira, D.P.; Sehr, A.; Renz, S.; Reinel, S.A.; Ferraz, E.R.; de Marchi, M.R.R.; Machado, M.C.; Zocolo, G.J.; et al. Genotoxicity assessment of water soluble fractions of biodiesel and its diesel blends using the Salmonella assay and the in vitro MicroFlow® kit (Litron) assay. Chemosphere 2012, 86, 512–520. [Google Scholar] [CrossRef]
- Lockard, J.M.; Prater, J.W.; Viau, C.J.; Enoch, H.G.; Sabharwal, P.S. Comparative study of the genotoxic properties of Eastern and Western U.S. shale oils, crude petroleum, and coal-derived oil. Mutat. Res. Genet. Toxicol. 1982, 102, 221–235. [Google Scholar] [CrossRef]
- Petrilli, F.L.; Renzi, G.P.D.; Flora, S.D. Interaction between polycyclic aromatic hydrocarbons, crude oil and oil dispersants in the Salmonella mutagenesis assay. Carcinogenesis 1980, 1, 51–56. [Google Scholar] [CrossRef]
- Gatehouse, D. Mutagenicity of 1, 2 ring-fused acenaphthenes against S. typhimurium TA1537 and TA1538: Structure—Activity relationships. Mutat. Res. Genet. Toxicol. 1980, 78, 121–135. [Google Scholar] [CrossRef]
- McCann, J.; Spingarn, N.E.; Kobori, J.; Ames, B.N. Detection of carcinogens as mutagens: Bacterial tester strains with R factor plasmids. Proc. Natl. Acad. Sci. USA 1975, 72, 979–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, M.; Yoshida, D.; Mizusaki, S. Mutagenicity of polycyclic aromatic hydrocarbons and quinones on Salmonella typhimurium TA97. Mutat. Res. Genet. Toxicol. 1985, 156, 61–67. [Google Scholar] [CrossRef]
- Schreiner, C. Genetic Toxicity of Naphthalene: A Review. J. Toxicol. Environ. Health Part B 2003, 6, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Muthusamy, S.; Xia, Q.; Lal, V.; Denison, M.S.; Ng, J.C. Micronucleus formation by single and mixed heavy metals/loids and PAH compounds in HepG2 cells. Mutagenesis 2015, 30, 593–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthusamy, S.; Peng, C.; Ng, J.C. Genotoxicity evaluation of multi-component mixtures of polyaromatic hydrocarbons (PAHs), arsenic, cadmium, and lead using flow cytometry based micronucleus test in HepG2 cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 827, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Brusick, D.; Small, M.S.; Cavalieri, E.L.; Chakravarti, D.; Ding, X.; Longfellow, D.G.; Nakamura, J.; Rogan, E.C.; Swenberg, J.A. Possible genotoxic modes of action for naphthalene. Regul. Toxicol. Pharmacol. 2008, 51, 43–50. [Google Scholar] [CrossRef]
- Rudnicka, K.; Łuczyński, M.K.; Góra, M. Genotoxicity of cyclopentha [c] phenanthrene and its two derivatives based on an in vitro micronucleus test. Environ. Biotechnol. 2012, 8, 63–66. [Google Scholar]
- Redman, A.D.; Parkerton, T.F. Guidance for improving comparability and relevance of oil toxicity tests. Mar. Pollut. Bull. 2015, 98, 156–170. [Google Scholar] [CrossRef]
- Redman, A.D.; Butler, J.D.; Letinski, D.J.; Di Toro, D.M.; Paumen, M.L.; Parkerton, T.F. Technical basis for using passive sampling as a biomimetic extraction procedure to assess bioavailability and predict toxicity of petroleum substances. Chemosphere 2018, 199, 585–594. [Google Scholar] [CrossRef]
- Couillard, C.M.; Lee, K.; Légaré, B.; King, T.L. Effect of dispersant on the composition of the water-accommodated fraction of crude oil and its toxicity to larval marine fish. Environ. Toxicol. Chem. 2005, 24, 1496. [Google Scholar] [CrossRef] [PubMed]
- Dussauze, M.; Pichavant-Rafini, K.; Le Floch, S.; Lemaire, P.; Theron, M. Acute toxicity of chemically and mechanically dispersed crude oil to juvenile sea bass (Dicentrarchus labrax): Absence of synergistic effects between oil and dispersants. Environ. Toxicol. Chem. 2015, 34, 1543–1551. [Google Scholar] [CrossRef]
- Ramachandran, S.D.; Hodson, P.V.; Khan, C.W.; Lee, K. Oil dispersant increases PAH uptake by fish exposed to crude oil. Ecotoxicol. Environ. Saf. 2004, 59, 300–308. [Google Scholar] [CrossRef]
- Clark, J.; Becker, K.; Venosa, A.; Lewis, A. Assessing dispersant effectiveness for heavy fuel oils using small-scale laboratory tests. Int. Oil Spill Conf. Proc. 2005, 2005, 59–63. [Google Scholar] [CrossRef]
- Li, Z.; Lee, K.; King, T.; Boufadel, M.C.; Venosa, A.D. Effects of temperature and wave conditions on chemical dispersion efficacy of heavy fuel oil in an experimental flow-through wave tank. Mar. Pollut. Bull. 2010, 60, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, D.; Reeve, L.; Gatehouse, D.; Vanparys, P. A core in vitro genotoxicity battery comprising the Ames test plus the in vitro micronucleus test is sufficient to detect rodent carcinogens and in vivo genotoxins. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 721, 27–73. [Google Scholar] [CrossRef] [PubMed]
- Claxton Larry, D.; de A. Umbuzeiro, G.; DeMarini David, M. The Salmonella Mutagenicity Assay: The Stethoscope of Genetic Toxicology for the 21st Century. Environ. Health Perspect. 2010, 118, 1515–1522. [Google Scholar] [CrossRef] [Green Version]
- Gee, P.; Maron, D.M.; Ames, B.N. Detection and classification of mutagens: A set of base-specific Salmonella tester strains. Proc. Natl. Acad. Sci. USA 1994, 91, 11606–11610. [Google Scholar] [CrossRef] [Green Version]
- Kamber, M.; Fluckiger-Isler, S.; Engelhardt, G.; Jaeckh, R.; Zeiger, E. Comparison of the Ames II and traditional Ames test responses with respect to mutagenicity, strain specificities, need for metabolism and correlation with rodent carcinogenicity. Mutagenesis 2009, 24, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Fowler, P.; Smith, K.; Young, J.; Jeffrey, L.; Kirkland, D.; Pfuhler, S.; Carmichael, P. Reduction of misleading (“false”) positive results in mammalian cell genotoxicity assays. I. Choice of cell type. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2012, 742, 11–25. [Google Scholar] [CrossRef]
- Pfuhler, S.; Fellows, M.; Van Benthem, J.; Corvi, R.; Curren, R.; Dearfield, K.; Fowler, P.; Frötschl, R.; Elhajouji, A.; Le Hégarat, L. In vitro genotoxicity test approaches with better predictivity: Summary of an IWGT workshop. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 723, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.R.; Velasques, R.R.; Hoff, M.L.M.; Souza, M.M.; Sandrini, J.Z. Characterization of different DNA repair pathways in hepatic cells of Zebrafish (Danio rerio). DNA Repair 2019, 83, 102695. [Google Scholar] [CrossRef] [PubMed]
- Fowler, P.; Smith, R.; Smith, K.; Young, J.; Jeffrey, L.; Kirkland, D.; Pfuhler, S.; Carmichael, P. Reduction of misleading (“false”) positive results in mammalian cell genotoxicity assays. II. Importance of accurate toxicity measurement. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2012, 747, 104–117. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Compound | MGO (µg L−1) | IFO180 [µg L−1] |
|---|---|---|
| Naphthalene | 229.52 | 219.80 |
| Fluorene | 6.10 | 4.64 |
| Phenanthrene | 7.33 | 8.77 |
| Anthracene | n.d. | 0.83 |
| Fluoranthene | 0.12 | 0.12 |
| Pyrene | 0.25 | 0.39 |
| 11h-benzo[a]fluorene | 0.34 | 0.34 |
| 11h-benzo[b]fluorene | 0.21 | 0.24 |
| Benzo[a]anthracene | 0.10 | 0.13 |
| Chrysene | 0.18 | 0.34 |
| Benzo[b]fluoranthene | 0.08 | 0.09 |
| Benzo[k]fluoranthene | 0.11 | 0.10 |
| Benzo[a]pyrene | 0.08 | 0.11 |
| Benzo[e]pyrene | 0.10 | 0.09 |
| Indeno[1,2,3 cd]pyrene | n.d. | n.d. |
| Dibenz[a,h]anthracene | n.d. | n.d. |
| Benzo[ghi]perylene | n.d. | n.d. |
| Dibenzo[a,e]pyrene | n.d. | n.d. |
| Σ PAHs | 244.50 | 235.99 |
| Treatment | Mean Specific Activity (ng Curc. µL−1 Sample) | SD | LOQ (ng Curc. µL−1 Sample) | SD |
|---|---|---|---|---|
| LEWAF NNS | <LOQ | 2.0 | 0.7 | |
| CEWAF NNS | 19.2 | 10.6 | ||
| LEWAF MGO | <LOQ | 2.7 | 1.1 | |
| CEWAF MGO | <LOQ | 8.2 | 1.5 | |
| LEWAF IFO 180 | 9.0 | 7.0 | ||
| CEWAF IFO 180 | <LOQ | 9.1 | 0.5 | |
| HEWAF Fin 51 | 21.0 | 8.4 | ||
| HEWAF Mig 812 | 20.0 | 13.2 | ||
| HEWAF Fin51/Mig812 | 32.1 | 33.7 |
| Treatment | LEWAF (% of Stock) | IF Mean | SD | CEWAF (% of Stock) | IF Mean | SD |
|---|---|---|---|---|---|---|
| NNS | 50 | 2.01 | 0.29 | 1 | 2.61 | 0.73 |
| 25 | 1.11 | 0.16 | 0.5 | 2.37 | 0.79 | |
| MGO | 66 | 1.35 | 0.22 | 16 | 2.74 | 0.54 |
| 33 | 1.42 | 0.42 | 8 | 1.90 | 0.24 | |
| IFO 180 | 66 | 1.32 | 0.37 | 16 | 1.25 | 0.51 |
| 33 | 1.30 | 0.43 | 8 | 1.35 | 0.29 | |
| Fin51 | 1 | 1.33 | 0.09 | |||
| 0.5 | 1.24 | 0.12 | ||||
| Mig 812 | 1 | 0.97 | 0.14 | |||
| 0.5 | 1.04 | 0.14 | ||||
| Fin51/Mig812 | 1 | 1.32 | 0.38 | |||
| 0.5 | 1.30 | 0.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johann, S.; Goßen, M.; Behnisch, P.A.; Hollert, H.; Seiler, T.-B. Combining Different In Vitro Bioassays to Evaluate Genotoxicity of Water-Accommodated Fractions from Petroleum Products. Toxics 2020, 8, 45. https://doi.org/10.3390/toxics8020045
Johann S, Goßen M, Behnisch PA, Hollert H, Seiler T-B. Combining Different In Vitro Bioassays to Evaluate Genotoxicity of Water-Accommodated Fractions from Petroleum Products. Toxics. 2020; 8(2):45. https://doi.org/10.3390/toxics8020045
Chicago/Turabian StyleJohann, Sarah, Mira Goßen, Peter A. Behnisch, Henner Hollert, and Thomas-Benjamin Seiler. 2020. "Combining Different In Vitro Bioassays to Evaluate Genotoxicity of Water-Accommodated Fractions from Petroleum Products" Toxics 8, no. 2: 45. https://doi.org/10.3390/toxics8020045
APA StyleJohann, S., Goßen, M., Behnisch, P. A., Hollert, H., & Seiler, T. -B. (2020). Combining Different In Vitro Bioassays to Evaluate Genotoxicity of Water-Accommodated Fractions from Petroleum Products. Toxics, 8(2), 45. https://doi.org/10.3390/toxics8020045