
Chronic Dietary Exposure of Roosters to a Glyphosate-Based Herbicide Increases Seminal Plasma Glyphosate and AMPA Concentrations, Alters Sperm Parameters, and Induces Metabolic Disorders in the Progeny

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Issues
2.2. Animals
2.3. Treatment of Groups and Preparation of the Feed
2.4. Biological Samples
2.5. Testosterone, Oestradiol and Chemerin Assays
2.6. Evaluation of Sperm Motility Using IVOS (Integrated Visual Optical System)
2.7. Determination of Adenosine Triphosphate (ATP) and Calcium Concentration in Spermatozoa
2.8. Immunofluorescence
2.9. Histological Examination of the Testes
2.10. Fertility Parameters
2.11. Glyphosate and AMPA Assays in Seminal Liquid and Plasma
2.12. Western Blot
2.13. Determination of Mortality, Food Consumption, Body and Different Organ Weights in Offspring
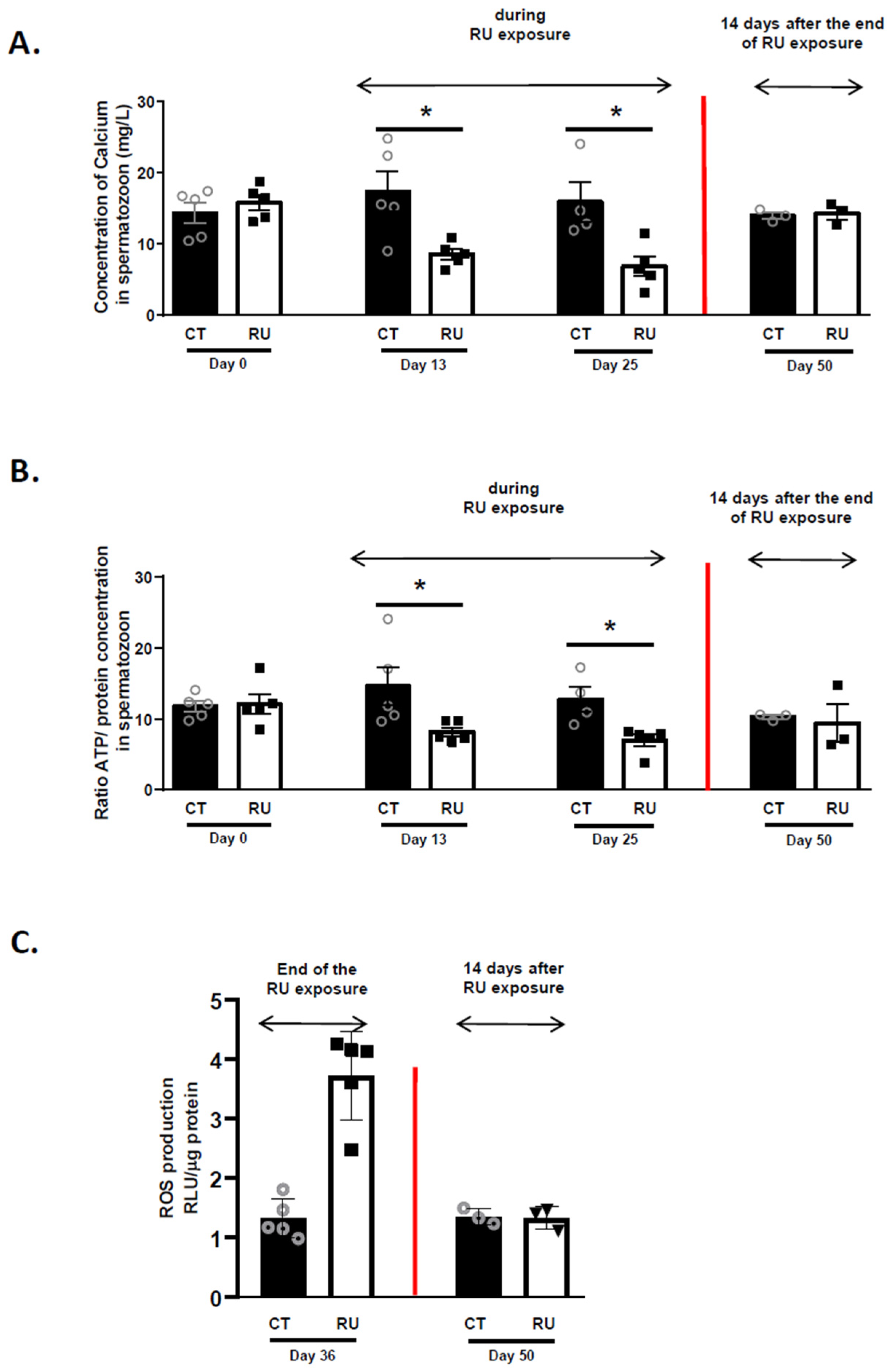
2.14. Oxidative Stress in Spermatozoa
2.15. Analysis of Genomic DNA Methylation of the CT and RU Spermatozoa
2.16. Statistical Analysis
3. Results
3.1. Measurement of Glyphosate (G) and AMPA Concentrations in the Blood Plasma (BP) and the Seminal Fluid (SF) of Roosters
3.2. Evaluation of Chronic RU Dietary Exposure on Sperm Concentration and Motility in Roosters
3.3. Effect of Roundup Dietary Exposure on Plasma Steroid Concentration in Roosters
3.4. Effect of Roundup Dietary Exposure on In Vivo Fertility
3.5. Effect of Paternal Chronic Dietary Roundup Exposure on Mortality, the Food Intake, Growth and Fattening of the Progeny
3.6. Analysis of Genomic DNA Methylation of CT and RU Spermatozoa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPA | aminomethylphosphonic |
| ATP | Adenosine Triphosphate |
| 3 β-HSD | 3β-hydroxysteroid dehydrogenase |
| CASA | Computer-assisted sperm assessment |
| CT | Control |
| CYP17 | cytochrome P450 17A1 |
| CYP19 | P450 aromatase |
| DBP | di(n-butyl) phthalate |
| EFSA | European Food Safety Authority |
| E2 | estrogen |
| ER | estrogen receptor |
| eEF1α1 | cycle-specific eukaryotic translation elongation factor 1 subunit alpha 1 |
| EEM | early embryonic mortality |
| G | Glyphosate |
| GBHs | glyphosate-based-herbicides |
| IVOS | Integrated Visual Optical System |
| LEM | Late embryonic mortality |
| MRL | maximum residue level |
| NOAEL | No observable adverse effects level |
| P | progesterone |
| PBS | Phosphate Buffered Saline |
| P450scc | cytochrome P450 side-chain cleavage |
| ROS | Reactive Oxygen Species |
| RU | Roundup |
| StAR | steroidogenic acute regulatory |
| T | Testosterone |
| VAP | Average path velocity |
| VCL | curvilinear velocity |
| VSL | straight-line velocity |
References
- Agostini, L.P.; Dettogni, R.S.; dos Reis, R.S.; Stur, E.; dos Santos, E.V.W.; Ventorim, D.P.; Garcia, F.M.; Cardoso, R.C.; Graceli, J.B.; Louro, I.D. Effects of glyphosate exposure on human health: Insights from epidemiological and in vitro studies. Sci. Total Environ. 2020, 705, 135808. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, R.; Defarge, N.; de Vendômois, J.S.; Séralini, G.E. Potential toxic effects of glyphosate and its commercial formulations below regulatory limits. Food Chem. Toxicol. 2015, 84, 133–153. [Google Scholar] [CrossRef] [Green Version]
- Gasnier, C.; Dumont, C.; Benachour, N.; Clair, E.; Chagnon, M.-C.; Séralini, G.-E. Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology 2009, 262, 184–191. [Google Scholar] [CrossRef]
- Gillezeau, C.; van Gerwen, M.; Shaffer, R.M.; Rana, I.; Zhang, L.; Sheppard, L.; Taioli, E. The evidence of human exposure to glyphosate: A review. Environ. Health 2019, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrell, Z.R.; Ahammad, M.U.; Benson, A.P. Glyphosate-based herbicide formulations and reproductive toxicity in animals. Vet. Anim. Sci. 2020, 10, 100126. [Google Scholar] [CrossRef]
- Martinez, A.; Al-Ahmad, A.J. Effects of glyphosate and aminomethylphosphonic acid on an isogeneic model of the human blood-brain barrier. Toxicol. Lett. 2019, 304, 39–49. [Google Scholar] [CrossRef]
- Teleken, J.L.; Gomes, E.C.Z.; Marmentini, C.; Moi, M.B.; Ribeiro, R.A.; Balbo, S.L.; Amorim, E.M.P.; Bonfleur, M.L. Glyphosate-Based herbicide exposure during pregnancy and lactation malprograms the male reproductive morphofunction in F1 offspring. J. Dev. Orig. Health Dis. 2020, 11, 146–153. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, N.; Yin, L.; Zhang, W.; Han, F.; Liu, W.; Chen, H.; Cao, J.; Liu, J. A Commercial Roundup® formulation induced male germ cell apoptosis by promoting the expression of XAF1 in adult mice. Toxicol. Lett. 2018, 296, 163–172. [Google Scholar] [CrossRef]
- Dallegrave, E.; Mantese, F.D.; Oliveira, R.T.; Andrade, A.J.M.; Dalsenter, P.R.; Langeloh, A. Pre- and postnatal toxicity of the commercial glyphosate formulation in Wistar rats. Arch. Toxicol. 2007, 81, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.K.L.; Schwartz, C.L.; Nielsen, L.N.; Boberg, J.; Vinggaard, A.M.; Bahl, M.I.; Svingen, T. Exposure to a glyphosate-based herbicide formulation, but not glyphosate alone, has only minor effects on adult rat testis. Reprod. Toxicol. 2018, 82, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Romano, R.M.; Romano, M.A.; Bernardi, M.M.; Furtado, P.V.; Oliveira, C.A. Prepubertal exposure to commercial formulation of the herbicide glyphosate alters testosterone levels and testicular morphology. Arch. Toxicol. 2010, 84, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, J.; Yang, L.; Zhang, H.; Zhang, Y.; Gao, D.; Jiang, H.; Li, Y.; Dong, H.; Ma, T.; et al. Glyphosate exposure attenuates testosterone synthesis via NR1D1 inhibition of StAR expression in mouse Leydig cells. Sci. Total Environ. 2021, 785, 147323. [Google Scholar] [CrossRef] [PubMed]
- Hamdaoui, L.; Naifar, M.; Rahmouni, F.; Harrabi, B.; Ayadi, F.; Sahnoun, Z.; Rebai, T. Subchronic exposure to kalach 360 SL-induced endocrine disruption and ovary damage in female rats. Arch. Physiol. Biochem. 2018, 124, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Li, R.; Liu, J.; Huang, K.; Wu, S.; Li, Y.; Li, C. Effects of glyphosate on the ovarian function of pregnant mice, the secretion of hormones and the sex ratio of their fetuses. Environ. Pollut. 2018, 243, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Feng, G.; Xiaoxu, W.; Peng, T.; Shengnan, Q.; Baoming, S.; Anshan, S. Effects of glyphosate-based herbicide-contaminated diets on reproductive organ toxicity and hypothalamic-pituitary-ovarian axis hormones in weaned piglets. Environ. Pollut. 2021, 272, 115596. [Google Scholar] [CrossRef]
- Sopko, B.; Tejral, G.; Bitti, G.; Abate, M.; Medvedikova, M.; Hajduch, M.; Chloupek, J.; Fajmonova, J.; Skoric, M.; Amler, E.; et al. Glyphosate interaction with EEF1α1 Indicates altered protein synthesis: Evidence for reduced spermatogenesis and cytostatic effect. ACS Omega 2021, 6, 14848–14857. [Google Scholar] [CrossRef] [PubMed]
- Plaza, P.I.; Martínez-López, E.; Lambertucci, S.A. The perfect threat: Pesticides and vultures. Sci. Total Environ. 2019, 687, 1207–1218. [Google Scholar] [CrossRef]
- Bello, U.M.; Madekurozwa, M.-C.; Groenewald, H.B.; Aire, T.A.; Arukwe, A. The effects on steroidogenesis and histopathology of adult male Japanese quails (Coturnix coturnix japonica) testis following pre-pubertal exposure to di(n-butyl) phthalate (DBP). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 166, 24–33. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA) Evaluation of the impact of glyphosate and its residues in feed on animal health. EFSA J. 2018, 16, e05283. [CrossRef]
- Ruuskanen, S.; Rainio, M.J.; Gómez-Gallego, C.; Selenius, O.; Salminen, S.; Collado, M.C.; Saikkonen, K.; Saloniemi, I.; Helander, M. Glyphosate-based herbicides influence antioxidants, reproductive hormones and gut microbiome but not reproduction: A long-term experiment in an avian model. Environ. Pollut. 2020, 266, 115108. [Google Scholar] [CrossRef]
- Ruuskanen, S.; Rainio, M.J.; Uusitalo, M.; Saikkonen, K.; Helander, M. Effects of parental exposure to glyphosate-based herbicides on embryonic development and oxidative status: A long-term experiment in a bird model. Sci. Rep. 2020, 10, 6349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.G.; Telles, L.F.; Hess, R.A.; Mahecha, G.A.B.; Oliveira, C.A. Effects of the herbicide roundup on the epididymal region of drakes Anas platyrhynchos. Reprod. Toxicol. 2007, 23, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Szabó, R.; Szemerédy, G.; Kormos, É.; Lehel, J.; Budai, P. Studies on joint toxic effects of a glyphosate herbicide (fozát 480) and a heavy metal (cadmium) on chicken embryos. AGROFOR 2018, 2. [Google Scholar] [CrossRef]
- Fathi, M.A.; Abdelghani, E.; Shen, D.; Ren, X.; Dai, P.; Li, Z.; Tang, Q.; Li, Y.; Li, C. Effect of in Ovo glyphosate injection on embryonic development, serum biochemistry, antioxidant status and histopathological changes in newly hatched chicks. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Fathi, M.A.; Han, G.; Kang, R.; Shen, D.; Shen, J.; Li, C. Disruption of cytochrome P450 enzymes in the liver and small intestine in chicken embryos in Ovo exposed to glyphosate. Environ. Sci. Pollut. Res. 2020, 27, 16865–16875. [Google Scholar] [CrossRef]
- Bakst, M.R.; Akuffo, V.; Trefil, P.; Brillard, J.P. Morphological and histochemical characterization of the seminiferous epithelial and Leydig cells of the turkey. Anim. Reprod. Sci. 2007, 97, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Michaylova, V.; Ilkova, P. Photometric determination of micro amounts of calcium with arsenazo III. Anal. Chim. Acta 1971, 53, 194–198. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Barbe, A.; Mellouk, N.; Ramé, C.; Grandhaye, J.; Anger, K.; Chahnamian, M.; Ganier, P.; Brionne, A.; Riva, A.; Froment, P.; et al. A grape seed extract maternal dietary supplementation improves egg quality and reduces ovarian steroidogenesis without affecting fertility parameters in reproductive hens. PLoS ONE 2020, 15, e0233169. [Google Scholar] [CrossRef] [PubMed]
- Estienne, A.; Reverchon, M.; Partyka, A.; Bourdon, G.; Grandhaye, J.; Barbe, A.; Caldas-Silveira, E.; Rame, C.; Niżański, W.; Froment, P.; et al. Chemerin impairs in vitro testosterone production, sperm motility, and fertility in chicken: Possible involvement of its receptor CMKLR1. Cells 2020, 9, 1599. [Google Scholar] [CrossRef]
- Eason, T.H.; Scanlon, P.F. Effects of atrazine and glyphosate ingestion on body weight and nutritional well-being of coturnix quail. Z. Für Jagdwiss. 2002, 48, 281–285. [Google Scholar] [CrossRef]
- Riechers, D.E.; Wax, L.M.; Liebl, R.A.; Bullock, D.G. Surfactant effects on glyphosate efficacy. Weed Technol. 1995, 9, 281–285. [Google Scholar] [CrossRef]
- Moore, L.J.; Fuentes, L.; Rodgers, J.H.; Bowerman, W.W.; Yarrow, G.K.; Chao, W.Y.; Bridges, W.C. Relative toxicity of the components of the original formulation of roundup® to five north American anurans. Ecotoxicol. Environ. Saf. 2012, 78, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Tush, D.; Maksimowicz, M.M.; Meyer, M.T. Dissipation of polyoxyethylene tallow amine (POEA) and glyphosate in an agricultural field and their co-occurrence on streambed sediments. Sci. Total Environ. 2018, 636, 212–219. [Google Scholar] [CrossRef]
- Kongtip, P.; Nankongnab, N.; Phupancharoensuk, R.; Palarach, C.; Sujirarat, D.; Sangprasert, S.; Sermsuk, M.; Sawattrakool, N.; Woskie, S.R. Glyphosate and paraquat in maternal and fetal serums in thai women. J. Agromedicine 2017, 22, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Zoller, O.; Rhyn, P.; Zarn, J.A.; Dudler, V. Urine glyphosate level as a quantitative biomarker of oral exposure. Int. J. Hyg. Environ. Health 2020, 228, 113526. [Google Scholar] [CrossRef]
- Williams, G.; Kroes, R.; Munro, I. Safety evaluation and risk assessment of the herbicide Roundup and its active ingredient, glyphosate, for humans. Regul. Toxicol. Pharmacol. 2000, 31, 117–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubsad, D.; Nilsson, E.E.; King, S.E.; Sadler-Riggleman, I.; Beck, D.; Skinner, M.K. Assessment of glyphosate induced epigenetic transgenerational inheritance of pathologies and sperm epimutations: Generational toxicology. Sci. Rep. 2019, 9, 6372. [Google Scholar] [CrossRef] [Green Version]
- Abarikwu, S.O.; Akiri, O.F.; Durojaiye, M.A.; Adenike, A. Combined effects of repeated administration of Bretmont wipeout (glyphosate) and Ultrazin (atrazine) on testosterone, oxidative stress and sperm quality of Wistar Rats. Toxicol. Mech. Methods 2015, 25, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Owagboriaye, F.O.; Dedeke, G.A.; Ademolu, K.O.; Olujimi, O.O.; Ashidi, J.S.; Adeyinka, A.A. Reproductive toxicity of roundup herbicide exposure in male albino rat. Exp. Toxicol. Pathol. 2017, 69, 461–468. [Google Scholar] [CrossRef]
- Razi, M.; Shahmohamadloo, S. Histological and histochemical effects of Gly-phosate on testicular tissue and function. Iran. J. Reprod. Med. 2012, 10, 181. [Google Scholar] [PubMed]
- Mutwedu, V.B.; Nyongesa, A.W.; Azine, P.C.; Chiregereza, D.K.; Ngoumtsop, V.H.; Mugumaarhahama, Y.; Ayagirwe, R.B.B. Growth performance and reproductive function impairment of glyphosate-based herbicide in male guinea pig (Cavia porcellus). Vet. Med. Sci. 2021, 7, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Nerozzi, C.; Recuero, S.; Galeati, G.; Bucci, D.; Spinaci, M.; Yeste, M. Effects of roundup and its main component, glyphosate, upon mammalian sperm function and survival. Sci. Rep. 2020, 10, 11026. [Google Scholar] [CrossRef] [PubMed]
- Anifandis, G.; Amiridis, G.; Dafopoulos, K.; Daponte, A.; Dovolou, E.; Gavriil, E.; Gorgogietas, V.; Kachpani, E.; Mamuris, Z.; Messini, C.; et al. The in vitro impact of the herbicide roundup on human sperm motility and sperm mitochondria. Toxics 2017, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales Velásquez, M.; Velásquez Rivera, V.; Cardona Maya, W.D. The effect of glyphosate on human sperm: In vitro approximation. Rev. Urol. Colomb. Colomb. Urol. J. 2021, 30, e194–e198. [Google Scholar] [CrossRef]
- Parodi, J. Motility, viability, and calcium in the sperm cells. Syst. Biol. Reprod. Med. 2014, 60, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Moreno, J.; Blesbois, E. Functional aspects of seminal plasma in bird reproduction. Int. J. Mol. Sci. 2020, 21, 5664. [Google Scholar] [CrossRef]
- de Liz Oliveira Cavalli, V.L.; Cattani, D.; Heinz Rieg, C.E.; Pierozan, P.; Zanatta, L.; Benedetti Parisotto, E.; Wilhelm Filho, D.; Mena Barreto Silva, F.R.; Pessoa-Pureur, R.; Zamoner, A. Roundup disrupts male reproductive functions by triggering calcium-mediated cell death in rat testis and Sertoli cells. Free Radic. Biol. Med. 2013, 65, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Bongrani, A.; Elfassy, Y.; Brun, J.; Ramé, C.; Mellouk, N.; Fellahi, S.; Bastard, J.; Levy, R.; Vasseur, C.; Froment, P.; et al. Expression of adipokines in seminal fluid of men of normal weight. Asian J. Androl. 2019, 21, 528. [Google Scholar] [CrossRef]
- Dai, P.; Hu, P.; Tang, J.; Li, Y.; Li, C. Effect of glyphosate on reproductive organs in male rat. Acta Histochem. 2016, 118, 519–526. [Google Scholar] [CrossRef]
- Yousef, M.I.; Salem, M.H.; Ibrahim, H.Z.; Helmi, S.; Seehy, M.A.; Bertheussen, K. Toxic effects of carbofuran and glyphosate on semen characteristics in rabbits. J. Environ. Sci. Health Part B 1995, 30, 513–534. [Google Scholar] [CrossRef] [PubMed]
- Nardi, J.; Moras, P.B.; Koeppe, C.; Dallegrave, E.; Leal, M.B.; Rossato-Grando, L.G. Prepubertal subchronic exposure to soy milk and glyphosate leads to endocrine disruption. Food Chem. Toxicol. 2017, 100, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.A.; Romano, R.M.; Santos, L.D.; Wisniewski, P.; Campos, D.A.; de Souza, P.B.; Viau, P.; Bernardi, M.M.; Nunes, M.T.; de Oliveira, C.A. Glyphosate impairs male offspring reproductive development by disrupting gonadotropin expression. Arch. Toxicol. 2012, 86, 663–673. [Google Scholar] [CrossRef]
- Liu, J.-B.; Chen, K.; Li, Z.-F.; Wang, Z.-Y.; Wang, L. Glyphosate-induced gut microbiota dysbiosis facilitates male reproductive toxicity in rats. Sci. Total Environ. 2022, 805, 150368. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Derian, L.; Kervarrec, C.; Kernanec, P.-Y.; Jégou, B.; Smagulova, F.; Gely-Pernot, A. Perinatal exposure to glyphosate and a glyphosate-based herbicide affect spermatogenesis in mice. Toxicol. Sci. 2019, 169, 260–271. [Google Scholar] [CrossRef]
- Popoola, S.O.; Sakpa, C.L. Hormones of pituitary-gonadal axis and histology of pituitary gland following oral treatment of male Wistar rats with glyphosate. J. Biomed. Res. Clin. Pract. 2018, 1, 164–169. [Google Scholar] [CrossRef]
- Lorenz, V.; Pacini, G.; Luque, E.H.; Varayoud, J.; Milesi, M.M. Perinatal exposure to glyphosate or a glyphosate-based formulation disrupts hormonal and uterine milieu during the receptive state in rats. Food Chem. Toxicol. 2020, 143, 111560. [Google Scholar] [CrossRef]
- Milesi, M.M.; Lorenz, V.; Pacini, G.; Repetti, M.R.; Demonte, L.D.; Varayoud, J.; Luque, E.H. Perinatal exposure to a glyphosate-based herbicide impairs female reproductive outcomes and induces second-generation adverse effects in Wistar rats. Arch. Toxicol. 2018, 92, 2629–2643. [Google Scholar] [CrossRef]
- Ait-Bali, Y.; Ba-M’hamed, S.; Gambarotta, G.; Sassoè-Pognetto, M.; Giustetto, M.; Bennis, M. Pre- and postnatal exposure to glyphosate-based herbicide causes behavioral and cognitive impairments in adult mice: Evidence of cortical ad hippocampal dysfunction. Arch. Toxicol. 2020, 94, 1703–1723. [Google Scholar] [CrossRef]
- Gigante, P.; Berni, M.; Bussolati, S.; Grasselli, F.; Grolli, S.; Ramoni, R.; Basini, G. Glyphosate affects swine ovarian and adipose stromal cell functions. Anim. Reprod. Sci. 2018, 195, 185–196. [Google Scholar] [CrossRef]
- Winnick, B.; Dzialowski, E.M. The effects of glyphosaote based herbicides on chick embryo morphology during development. FASEB J. 2013, 27, 874.12. [Google Scholar] [CrossRef]
- Ganesan, S.; Keating, A.F. Ovarian mitochondrial and oxidative stress proteins are altered by glyphosate exposure in mice. Toxicol. Appl. Pharmacol. 2020, 402, 115116. [Google Scholar] [CrossRef] [PubMed]
- Maamar, M.B.; Beck, D.; Nilsson, E.E.; Kubsad, D.; Skinner, M.K. Epigenome-wide association study for glyphosate induced transgenerational sperm dna methylation and histone retention epigenetic biomarkers for disease. Epigenetics 2020, 19, 1150–1167. [Google Scholar] [CrossRef]
- Smith, C.M.; Vera, M.K.M.; Bhandari, R.K. Developmental and epigenetic effects of roundup and glyphosate exposure on Japanese medaka (Oryzias latipes). Aquat. Toxicol. 2019, 210, 215–226. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | % |
|---|---|
| Corn | 39.64 |
| Wheat | 23.86 |
| Soybean meal | 29.3 |
| Soy oil | 2.2 |
| Sunflower oil | 1 |
| Sodium bicarbonate | 0.18 |
| Calcium carbonate | 0.86 |
| Phosphate | 2.06 |
| Salt | 0.3 |
| Methionine DL | 0.1 |
| Mineral premix | 0.5 |
| Sperm Parameters | Days | CT | SEM | RU | SEM |
|---|---|---|---|---|---|
| VAP (μm/s) | Day 0 | 60.3 | ±3.2 | 60.7 | ±2.3 |
| Day 5 | 74.4 | ±1.0 | 66.5 * | ±3.3 | |
| Day 13 | 61.2 | ±2.3 | 56.8 | ±2.2 | |
| Day 25 | 71.8 | ±4.1 | 61.6 * | ±2.2 | |
| Day 50 | 65.9 | ±3.6 | 64.6 | ±1.0 | |
| VSL (μm/s) | Day 0 | 39.4 | ±3.5 | 40.5 | ±2.2 |
| Day 5 | 55.1 | ±1.3 | 46.6 * | ±3.4 | |
| Day 13 | 40.5 | ±1.6 | 34.1 * | ±2.4 | |
| Day 25 | 51.7 | ±3.3 | 39.7 *** | ±2.1 | |
| Day 50 | 44.6 | ±4.1 | 43.0 | ±1.1 | |
| VCL (μm/s) | Day 0 | 133.5 | ±3.0 | 136.8 | ±4.0 |
| Day 5 | 140.9 | ±1.8 | 136.7 | ±3.9 | |
| Day 13 | 133.0 | ±3.1 | 125.6 | ±2.5 | |
| Day 25 | 142.4 | ±3.4 | 131.8 * | ±2.9 | |
| Day 50 | 129.5 | ±5.7 | 129.3 | ±0.9 | |
| Progressive motility (%) | Day 0 | 15.6 | ±3.4 | 11.8 | ±2.4 |
| Day 5 | 35.3 | ±2.3 | 15.4 ** | ±4.5 | |
| Day 13 | 12.2 | ±1.8 | 3.9 ** | ±0.8 | |
| Day 25 | 24.9 | ±3.5 | 9.6 * | ±3.4 | |
| Day 50 | 14.3 | ±3.5 | 12.1 | ±3.0 | |
| Speed (μm/s) | Day 0 | 43.1 | ±6.2 | 31.3 | ±5.0 |
| Day 5 | 63.7 | ±2.6 | 33.9 *** | ±6.6 | |
| Day 13 | 31.0 | ±2.9 | 15.3 *** | ±2.1 | |
| Day 25 | 52.4 | ±4.2 | 26.9 * | ±7.2 | |
| Day 50 | 32.1 | ±4.0 | 30.3 | ±4.5 |
| Parameters | Sperm Pool from 5 CT Roosters | Sperm Pool from 5 RU Roosters | p-Value |
|---|---|---|---|
| Unfertilised | 6.75 ± 0.89 | 6.75 ± 0.75 | 0.90 |
| EEM | 2.51 ± 1.04 | 1.51 ± 0.92 | 0.44 |
| LEM | 0.80 ± 0.80 | 0.83 ± 0.83 | 0.99 |
| Hatchability of fertile eggs | 90.87 ± 3.40 | 91.11 ± 2.37 | 0.99 |
| Fertility | 93.98 ± 3.42 | 93.28 ± 2.10 | 0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, L.; Estienne, A.; Bourdon, G.; Ramé, C.; Chevaleyre, C.; Didier, P.; Chahnamian, M.; El Balkhi, S.; Froment, P.; Dupont, J. Chronic Dietary Exposure of Roosters to a Glyphosate-Based Herbicide Increases Seminal Plasma Glyphosate and AMPA Concentrations, Alters Sperm Parameters, and Induces Metabolic Disorders in the Progeny. Toxics 2021, 9, 318. https://doi.org/10.3390/toxics9120318
Serra L, Estienne A, Bourdon G, Ramé C, Chevaleyre C, Didier P, Chahnamian M, El Balkhi S, Froment P, Dupont J. Chronic Dietary Exposure of Roosters to a Glyphosate-Based Herbicide Increases Seminal Plasma Glyphosate and AMPA Concentrations, Alters Sperm Parameters, and Induces Metabolic Disorders in the Progeny. Toxics. 2021; 9(12):318. https://doi.org/10.3390/toxics9120318
Chicago/Turabian StyleSerra, Loïse, Anthony Estienne, Guillaume Bourdon, Christelle Ramé, Claire Chevaleyre, Philippe Didier, Marine Chahnamian, Souleiman El Balkhi, Pascal Froment, and Joëlle Dupont. 2021. "Chronic Dietary Exposure of Roosters to a Glyphosate-Based Herbicide Increases Seminal Plasma Glyphosate and AMPA Concentrations, Alters Sperm Parameters, and Induces Metabolic Disorders in the Progeny" Toxics 9, no. 12: 318. https://doi.org/10.3390/toxics9120318
APA StyleSerra, L., Estienne, A., Bourdon, G., Ramé, C., Chevaleyre, C., Didier, P., Chahnamian, M., El Balkhi, S., Froment, P., & Dupont, J. (2021). Chronic Dietary Exposure of Roosters to a Glyphosate-Based Herbicide Increases Seminal Plasma Glyphosate and AMPA Concentrations, Alters Sperm Parameters, and Induces Metabolic Disorders in the Progeny. Toxics, 9(12), 318. https://doi.org/10.3390/toxics9120318