
Whole-Body Acute Contact Toxicity of Formulated Insecticide Mixtures to Blue Orchard Bees (Osmia lignaria)


Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Design
2.1.1. Bees
2.1.2. Experimental Cage Design
2.1.3. Spray Tower
2.1.4. Post Treatment Observations
2.2. Experiment 1: Male O. lignaria Acute Whole-Body Contact Toxicity to Premix Insecticides
2.3. Experiment 2: Female O. lignaria Acute Whole-Body Contact Toxicity to Premix Insecticides
2.4. Experiment 3: Male and Female O. lignaria Acute Whole-Body Contact Toxicity to Individual Active Ingredient Insecticides and 1:1 Binary Combinations
2.5. Statistical Analysis
3. Results
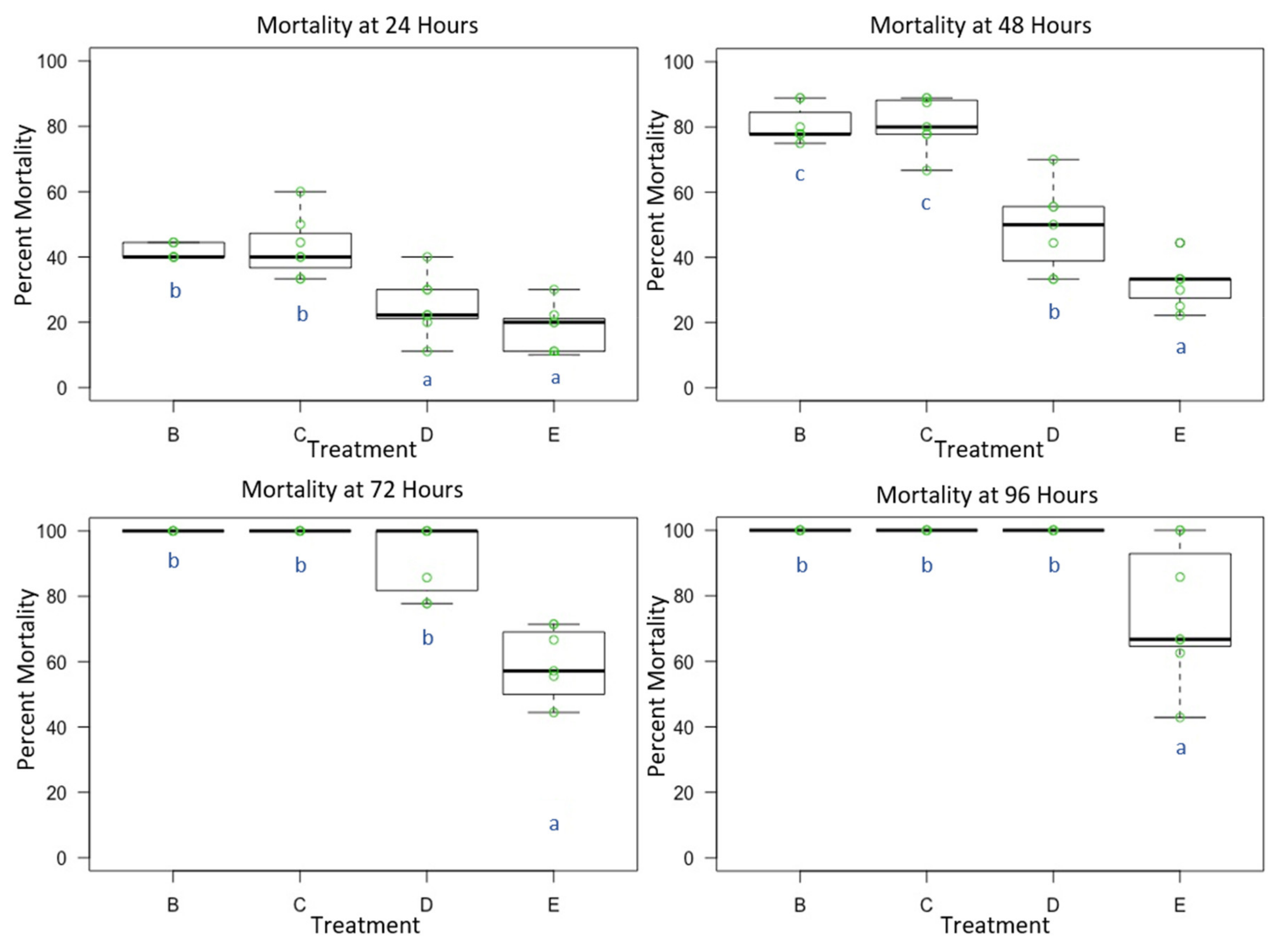
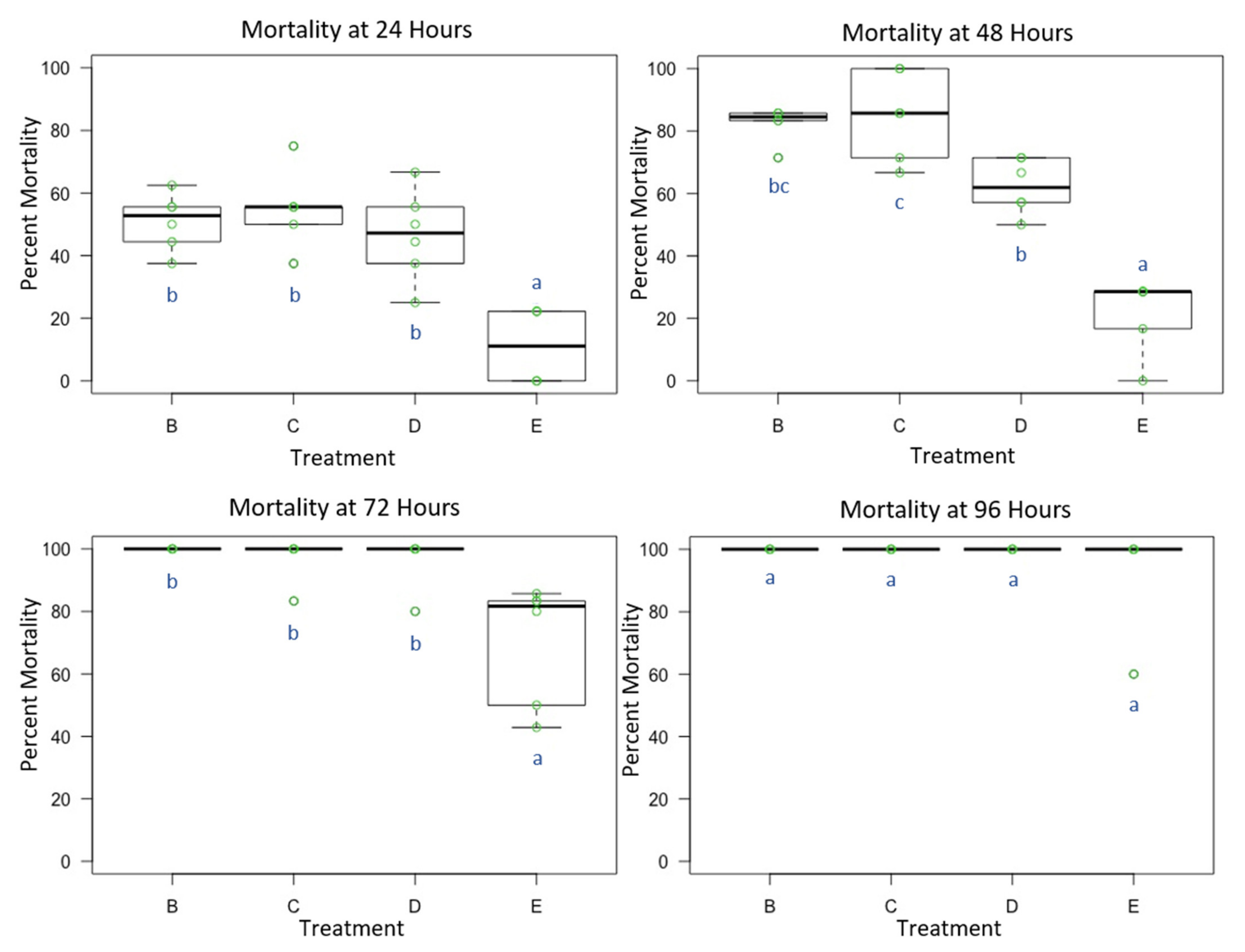
3.1. Experiment 1: Male O. lignaria Acute Whole-Body Contact Toxicity Following Whole Bodily Contact Exposure to Premix Insecticide Sprays
3.2. Experiment 2: Female O. lignaria Acute Contact Toxicity Following Whole-Body Contact Exposure to Premix Insecticides
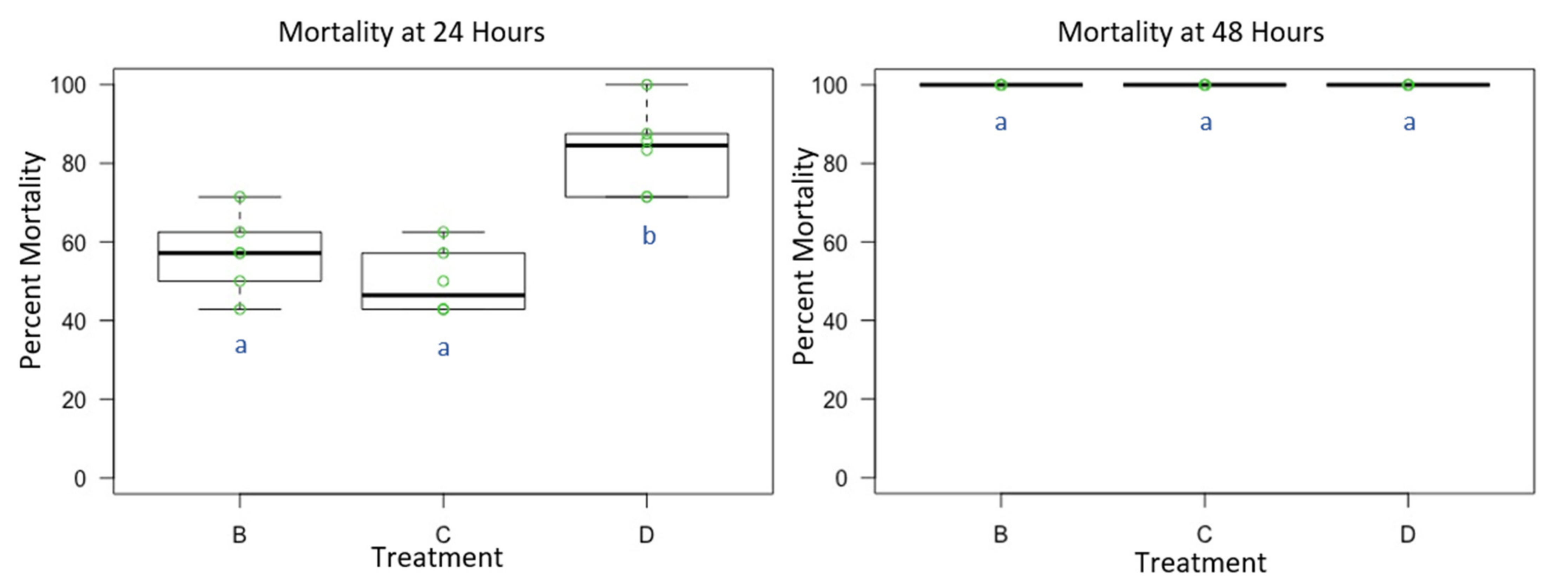
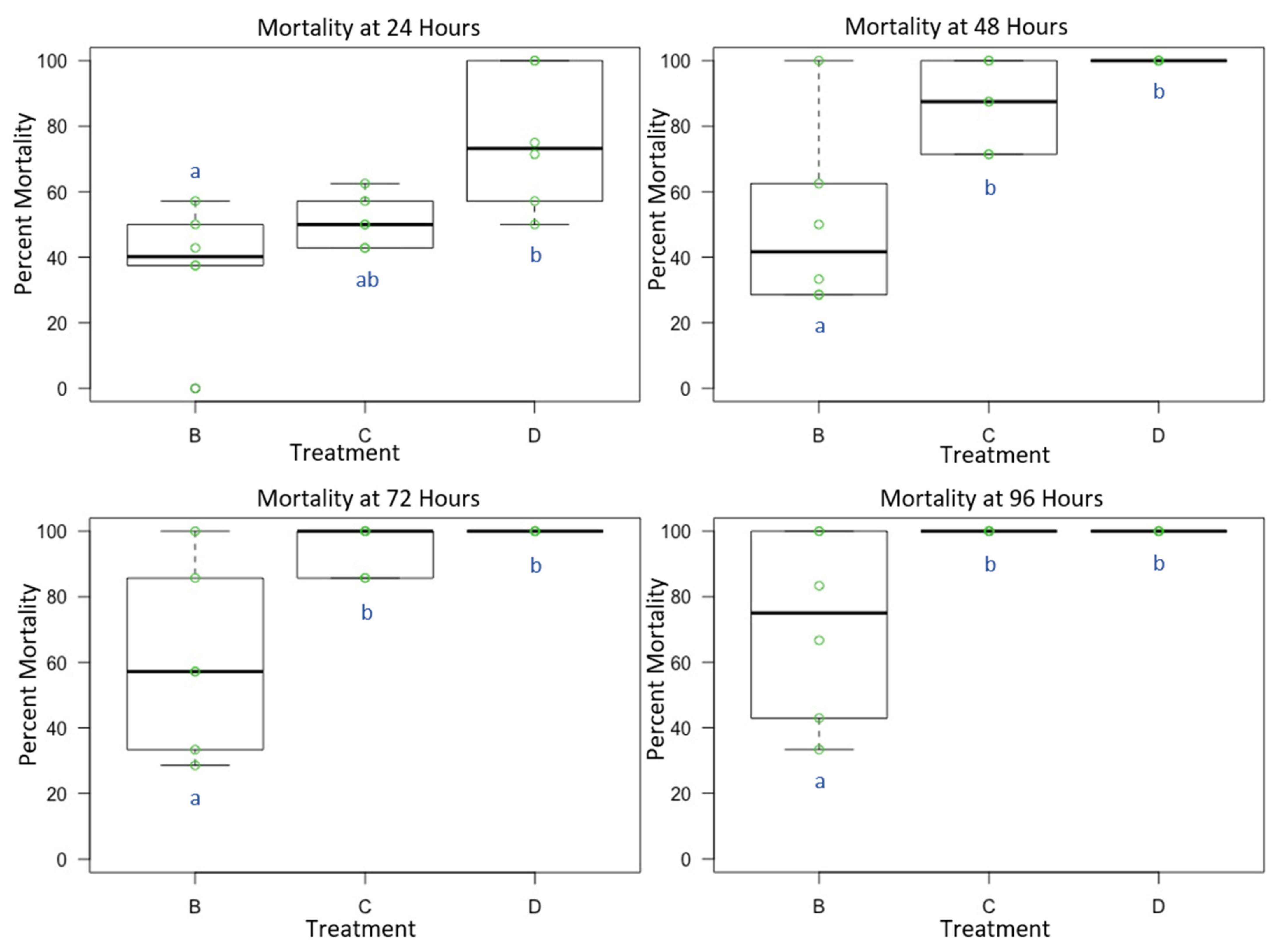
3.3. Experiment 3: Male and Female O. lignaria Acute Contact Toxicity Following Whole-Body Contact Exposure to Individual Active Ingredient Insecticides and 1:1 Binary Combinations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosch, J.; Kemp, W.P. How to Manage the Blue Orchard Bee: As an Orchard Pollinator; Sustainable Agriculture Network: Beltsville, MD, USA, 2001; p. 88. [Google Scholar]
- Bosch, J.; Kemp, W.P.; Trostle, G.E. Bee population returns and cherry yields in an orchard pollinated with Osmia lignaria (Hymenoptera: Megachilidae). J. Econ. Entomol. 2006, 99, 408–413. [Google Scholar] [CrossRef]
- Brittain, C.; Williams, N.; Kremen, C.; Klein, A.-M. Synergistic effects of non-Apis bees and honey bees for pollination services. Proc. R. Soc. B Biol. Sci. 2013, 280, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitts-Singer, T.L.; Artz, D.R.; Peterson, S.S.; Boyle, N.K.; Wardell, G.I. Examination of a managed pollinator strategy for almond production using Apis mellifera (Hymenoptera: Apidae) and Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2018, 47, 364–377. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N.K. Impact of botic and abiotic stressors on managed and feral bees. Insects 2019, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Heller, S.; Joshi, N.K.; Chen, J.; Rajotte, E.G.; Mullin, C.; Biddinger, D. Pollinator exposure to systemic insecticides and fungicides applied in the previous fall and pre-bloom period in apple orchards. Environ. Pollut. 2020, 265, 114589. [Google Scholar] [CrossRef]
- Abbott, V.A.; Nadeau, J.L.; Higo, H.A.; Winston, M.L. Lethal and sublethal effects of imidacloprid on Osmia lignaria and clothianidin on Megachile rotundata (Hymenoptera: Megachilidae). J. Econ. Entomol. 2008, 101, 13. [Google Scholar] [CrossRef]
- Ladurner, E.; Bosch, J.; Kemp, W.P.; Maini, S. Assessing delayed and acute toxicity of five formulated fungicides to Osmia lignaria Say and Apis mellifera. Apidologie 2005, 36, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Artz, D.R.; Pitts-Singer, T.L. Effects of fungicide and adjuvant sprays on nesting behavior in two managed solitary bees, Osmia lignaria and Megachile rotundata. PLoS ONE 2015, 10, e0135688. [Google Scholar] [CrossRef] [PubMed]
- Ilias, A.; Lagnel, J.; Kapantaidaki, D.E.; Roditakis, E.; Tsigenopoulos, C.S.; Vontas, J.; Tsagkarakou, A. Transcription analysis of neonicotinoid resistance in Mediterranean (MED) populations of B. tabaci reveal novel cytochrome P450s, but no nAChR mutations associated with the phenotype. BMC Genom. 2015, 16, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Højland, D.H.; Nauen, R.; Foster, S.P.; Williamson, M.S.; Kristensen, M. Incidence, spread and mechanisms of pyrethroid resistance in European populations of the cabbage stem flea beetle, Psylliodes chrysocephala L. (Coleoptera: Chrysomelidae). PLoS ONE 2015, 10, e0146045. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Adamczyk, J.; Rinderer, T.; Yao, J.; Danka, R.; Luttrell, R.; Gore, J. Spray toxicity and risk potential of 42 commonly used formulations of row crop pesticides to adult honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 2015, 108, 2640–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.C.; Yao, J.; Adamczyk, J.; Luttrell, R. Synergistic toxicity and physiological impact of imidacloprid alone and binary mixtures with seven representative pesticides on honey bee (Apis mellifera). PLoS ONE 2017, 12, e0176837. [Google Scholar] [CrossRef] [PubMed]
- Carnesecchi, E.; Toma, C.; Roncaglioni, A.; Kramer, N.; Benfenati, E.; Dorne, C.M.J.-L. Integrating QSAR models predicting acute contact toxicity and mode of action profiling in honey bees (A. mellifera): Data curation using open source databases, performance testing and validation. Sci. Total Environ. 2020, 735, 1–20. [Google Scholar] [CrossRef]
- Bibbs, C.S.; Fulcher, A.; Xue, R.-D. Allethrin-based mosquito control device causing knockdown, morbidity, and mortality in four species of field-caught mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.; Zoller, H.; Roepke, R.; Zschiesche, E.; Heckeroth, A.R. Fluralaner activity against life stages of ticks using Rhipicephalus sanguineus and Ornithodoros moubata IN in vitro contact and feeding assays. Parasit. Vectors 2015, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Glazer, I.; Navon, A. Activity and persistence of entomoparasitic nematodes tested against Heliothis armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 1990, 83, 1795–1800. [Google Scholar] [CrossRef]
- Wise, J.C.; Hulbert, D.; Vandervoort, C. Rainfall influences performance of insecticides on the codling moth (Lepidoptera: Tortricidae) in apples. Can. Entomol. 2017, 149, 118–128. [Google Scholar] [CrossRef]
- Phan, N.T.; Joshi, N.K.; Rajotte, E.G.; Lopez-Uribe, M.M.; Zhu, F.; Biddinger, D.J. A new ingestion bioassay protocol for assessing pesticide toxicity to the adult Japanese orchard bee (Osmia cornifrons). Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vijverberg, H.P.M.; van der Zalm, J.M.; van den Bercken, J. Similar mode of action of pyrethroids and DDT on sodium channel gating in myelinated nerves. Nature 1982, 295, 601–603. [Google Scholar] [CrossRef]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manag. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef]
- Gough, H.J.; Collins, L.G.; Everett, C.J.; Wilkinson, W. Acute Contact and Oral Toxicity to Honey Bees (Apis mellifera); ICI Americas Inc.: Wilmington, DE, USA, 2018. [Google Scholar]
- EPA. Pesticide Fact Sheet Number 164: Cyfluthrin; Office of Pesticide Programs, EPA U.S. Environmental Protection Agency: Washington, DC, USA, 1987.
- FAO. Specifications and Evaluations for Plant Protection Products: Beta-Cyfluthrin; FAO, Food and Agriculture Organization of the United Nations: Rome, Italy, 1999. [Google Scholar]
- Schmuck, R.; Schoning, R.; Stork, A.; Schramel, O. Risk posed to honey bees (Apis mellifera L., Hymenoptera) by an imidacloprid seed dressing of suflowers. Pest Manag. Sci. 2001, 57, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Syngenta. Thiamethoxam: Used to formulate Patinum®, Actara®, Centric®, Cruiser®, Flagship®, and Helix®. (Envirofacts: Syngenta Crop Protection Fact Sheet); Syngenta Crop Protection, Inc.: Greensboro, NC, USA, 2005; p. 7. [Google Scholar]
- Biddinger, D.J.; Robertson, J.L.; Mullin, C.; Frazier, J.; Ashcraft, S.A.; Rajotte, E.G.; Joshi, N.K.; Vaughn, M. Comparative toxicities and synergism of apple orchard pesticides to Apis mellifera (L.) and Osmia cornifrons (Radoszkowski). PLoS ONE 2013, 8, e72587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, J.L.; Russell, R.M.; Preisler, H.K.; Savin, N.E. Bioassays with Arthropods; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- EPA. Ecological Risk Assessment for Section 3 Registration for Fruit, Vegetable, Selected Field Crop, Turf and Ornamental Uses of Chlorantraniliprole (PC Code 090100); Office of Pesticide Programs, EPA U.S. Environmental Protection Agency: Washington, DC, USA, 2008; p. 37.
- CDPR. California Department of Pesticide Regulation Public Report: Methoxyfenozide; California Department of Pesticide Regulation: Sacramento, CA, USA.
- CDPR. California Department of Pesticide Regulation Public Report: Active Ingredient: Spinetoram; California Department of Pesticide Regulation: Sacramento, CA, USA.
- Smagghe, G.; Deknopper, J.; Meeus, I.; Mommaerts, V. Dietary chlorantraniliprole suppresses reproduction in worker bumblebees. Pest Manag. Sci. 2013, 69, 787–791. [Google Scholar] [CrossRef] [PubMed]
- IRAC. IRAC Mode of Action Classification Scheme; Version 8.4; Insecticide Resistance Action Committee: Brussels, Belgium, 20 May 2018; pp. 1–26. [Google Scholar]
- EPA. EFED Risk Assessment for the Proposed IR-4 Use of the Spinosad Product Entrust® on Pomegranate and Dates; EPA U.S. Environmental Protection Agency: Washington, DC, USA, 2009; pp. 1–85.
- Suiter, D.R.; Scharf, M.E. Insecticide Basics for the Pest Management Professional; University of Georgia: Athens, GA, USA, 2015; p. 1352. [Google Scholar]
- Mommaerts, V.; Sterk, G.; Smagghe, G. Bumblebees can be used in combination with juvenile hormone analogues and ecdysone agonists. Ecotoxicology 2006, 15, 513–521. [Google Scholar] [CrossRef]
- Besard, L.; Mommaerts, V.; Abdu-Alla, G.; Smagghe, G. Lethal and sublethal side-effect assessment supports a more benign profile of spinetoram compared with spinosad in the bumblebee Bombus terrestris. Pest Manag. Sci. 2011, 67, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Shimokawatoko, Y.; Yamaguchi, T.; Tanaka, H. Development of the Novel Insecticide Spinetoram (DIANA®). Sumitomo Kagaku 2012, 2012, 14. Available online: https://www.sumitomo-chem.co.jp/english/rd/report/files/docs/01_2012e.pdf (accessed on 16 March 2021).
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Acaricide, fungicide and drug interactions in honey bees (Apis mellifera). PLoS ONE 2013, 8, e54092. [Google Scholar] [CrossRef] [Green Version]
- EPA. Leverage 360® EPA Regulated Insecticide Label; Bayer Crop Science: Leverkusen, Germany, 2015. [Google Scholar]
- James, R.; Pitts-Singer, T.L. Bee Pollination in Agricultural Ecosystems; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Leskey, T.C.; Short, B.D.; Lee, D.-H. Efficacy of insecticide residues on adult Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) mortality and injury in apple and peach orchards: Residual insecticide efficacy on Halyomorpha halys. Pest Manag. Sci. 2013, 70, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Tanigoshi, L.K.; Gerdeman, B.S.; Hollis Spitler, G. Overview of Our First Season’s Experiences to Chemically Manage the Spotted Wing Drosophila through Lab and Field Research on Red Raspberry in Western Washington; Washington State University: Mount Vernon, WA, USA, 2011; pp. 15–16. [Google Scholar]
- Tanigoshi, L.K.; Spitler Hollis, G.; Gerdeman, B.S. Field Efficacy of Severaled Labeled and Experimental Insecticides for Spotted Wing Drosophila Control in Blueberry; Washington State University: Mount Vernon, WA, USA, 2013; pp. 105–107. [Google Scholar]
- Alston, D.; Murray, M. Leafrollers in Fruit Orchards; Utah State University: Logan, UT, USA, 2017; p. 5. [Google Scholar]
- Krawczyk, G.; Biddinger, D. Entomology. In Penn State Tree Fruit Production Guide; Penn State Extension: State College, PA, USA, 2018; p. 436. [Google Scholar]
- Doering, J.; Maus, C.; Schoening, R. Residues of Imidacloprid WG 5 in Blossom and Leaf Samples of Apple Trees After Soil Treatment in the Field; Bayer CropScience AG Report, No. G201819, Application: 2003, Sampling: 2004; Bayer AG: Monheim, Germany, 2004. [Google Scholar]
- Oliver, J.B.; Fare, D.C.; Yousef, N.; Scholl, S.S.; Reding, M.E.; Ranger, C.M.; Moyseenko, J.J.; Halcomb, M.A. Evaluation of a single application of neonicotinoid and multi-application contact insecticides for flatheaded borer management in field grown red maple cultivars. J. Environ. Hortic. 2010, 28, 135–149. [Google Scholar] [CrossRef]
- Mogren, C.L.; Lundgren, J.G. Neonicotinoid-contaminated pollinator strips adjacent to cropland reduce honey bee nutritional status. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Girolami, V.; Mazzon, L.; Squatini, A.; Mori, N.; Marzaro, M.; Dibernardo, A.; Greatti, M.; Giorio, C.; Tapparo, A. Translocation of neonicotinoid insecticides from coated seeds to seedling guttation drops: A novel way of intoxication for bees. J. Econ. Entomol. 2009, 102, 1808–1815. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.J.; Castle, S.J. Imidacloprid in melon guttation fluid: A potential mode of exposure for pest and beneficial organisms. J. Econ. Entomol. 2012, 105, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopwood, J.; Code, A.; Vaughan, M.; Biddinger, D.; Shepherd, M.; Black, S.H.; Lee-Mäder, E.; Mazzacano, C. How Neonicotinoids Can Kill Bees: The Science behind the Role These Insecticides Play in Harming Bees, 2nd ed.; The Xerces Society for Invertebrate Conservation: Portland, OR, USA, 2016; p. 84. [Google Scholar]
- Rodrigo, J.; Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Sci. Hortic. 2002, 92, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Horth, L.; Campbell, L.A. Supplementing small farms with native mason bees increases strawberry size and growth rate. J. Appl. Ecol. 2018, 55, 591–599. [Google Scholar] [CrossRef]
- Herrmann, J.D.; Beye, H.; de la Broise, C.; Hartlep, H.; Diekötter, T. Positive effects of the pollinators Osmia cornuta (Megachilidae) and Lucilia sericata (Calliphoridae) on strawberry quality. Arthropod Plant Interact. 2018, 13, 71–77. [Google Scholar] [CrossRef]
- West, T.; McCutcheon, T.W. Evaluating Osmia cornifrons as pollinators of highbush blueberry. Int. J. Fruit Sci. 2009, 9, 115–125. [Google Scholar] [CrossRef]
- Melito, S.; La Bella, S.; Martinelli, F.; Cammalleri, I.; Tuttolomondo, T.; Leto, C.; Mulas, M. Phenology study of Myrtus communis accessions selected from wild populations of Sicily: Preliminary results. ISHA Acta Hortic. 2015, 1172, 231–236. [Google Scholar]
- Dively, G.P.; Embrey, M.S.; Kamel, A.; Hawthorne, D.J.; Pettis, J.S. Assessment of chronic sublethal effects of imidacloprid on honey bee colony health. PLoS ONE 2015, 10, e0118748. [Google Scholar] [CrossRef]
- Sharma, A.; Reddy, G.V.P. IPM and pollinator protection in canola production in the USA. In Integrative Biological Control. Progress in Biological Control; Gao, Y., Hokkanen, H., Menzler-Hokkanen, I., Eds.; Springer: Berlin, Germany, 2020; Volume 20. [Google Scholar]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandrock, C.; Tanadini, L.G.; Pettis, J.S.; Biesmeijer, J.C.; Potts, S.G.; Neumann, P. Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success: Loss of pollinator fitness. Agric. For. Entomol. 2014, 16, 119–128. [Google Scholar] [CrossRef]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.; Gao, Z.; Zumkier, U. Large-scale monitoring of effects of clothianidin-dressed oilseed rape seeds on pollinating insects in Northern Germany: Effects on red mason bees (Osmia bicornis). Ecotoxicology 2016, 25, 1679–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Formulation | Active Ingredient(s) | Mode of Action 1st Insecticide | Mode of Action 2nd Insecticide | Manufacturer |
|---|---|---|---|---|---|
| A | Control (distilled water) | - | - | - | - |
| B | Endigo ZC® | Thiamethoxam (12.6%) + Lambda-cyhalothrin (9.48%) | Thiamethoxam -Nicotinic acetylcholine receptor (nAChR) competitive modulator | Lambda-cyhalothrin -Sodium channel modulator | Syngenta Crop Protection, LLC Greensboro, NC |
| C | Leverage 360® | Imidacloprid (21.0%) + Beta cyfluthrin (10.5%) | Imidacloprid -Nicotinic acetylcholine receptor (nAChR) competitive modulator | Beta-cyfluthrin -Sodium channel modulator | BayerCropScience LP Research Triangle Park, NC |
| D | Besiege® | Chlorantraniliprole (9.26%) + Lambda-cyhalothrin (4.63%) | Chlorantraniliprole -Ryanodine receptor modulator | Lambda-cyhalothrin -Sodium channel modulator | Syngenta Crop Protection, LLC Greensboro, NC |
| E | Intrepid Edge® | Methoxyfenozide (28.3%) + Spinetoram (5.66%) | Methoxyfenozide -Ecdysone receptor agonist (molt accelerating compound) | Spinetoram -Nicotinic acetylcholine receptor (nAChR) allosteric modulator | Dow AgroSciences LLC Indianapolis, IN |
| Treatment | Active Ingredient(s) | Application Concentration Administered (mL/Ha) | Highest Application Concentration on Label (mL/Ha) * | AI Concentration (ppm) in Formulated Product (per L) |
|---|---|---|---|---|
| A | Control (distilled water) | - | - | - |
| B | Thiamethoxam (12.6%) + Lambda-cyhalothrin (9.48%) | 401.76 | 438.28 | Thiamethoxam = 120 Lambda-cyhalothrin = 91 |
| C | Imidacloprid (21.0%) + Beta cyfluthrin (10.5%) | 189.92 | 204.53 | Imidacloprid = 98 Beta-cyfluthrin = 49 |
| D | Chlorantraniliprole (9.26%) + Lambda-cyhalothrin (4.63%) | 657.42 | 876.56 | Chlorantraniliprole = 56 Lambda-cyhalothrin = 28 |
| E | Methoxyfenozide (28.3%) + Spinetoram (5.66%) | 657.42 | 876.56 | Methoxyfenozide = 140 Spinetoram = 28 |
| Treatment | Formulation | Active Ingredient | Insecticide Mode of Action | Manufacturer |
|---|---|---|---|---|
| A | Control (distilled water) | - | - | - |
| B | Admire Pro® | Imidacloprid (42.8%) | Imidacloprid -Nicotinic acetylcholine receptor (nAChR) competitive modulator | BayerCropScience LP Research Triangle Park, NC |
| C | Baythroid XL® | Beta-cyfluthrin (12.7%) | Beta-cyfluthrin -Sodium channel modulator | BayerCropScience LP Research Triangle Park, NC |
| D | (1:1 binary combination by volume) of Treatment B and Treatment C | Imidacloprid + Beta-cyfluthrin | - | - |
| Treatment | Active Ingredient | Application Concentration Administered (mL/Ha) * | Highest Application Concentration on Label (mL/Ha) * | AI Concentration (ppm) in Formulated Product (per L) |
|---|---|---|---|---|
| A | Control (distilled water) | - | - | - |
| B | Imidacloprid (42.8%) | 102.27 | 204.53 | Imidacloprid = 120 |
| C | Beta-cyfluthrin (12.7%) | 102.27 | 204.53 | Beta-cyfluthrin = 26 |
| D ** | Imidacloprid + Beta-cyfluthrin | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belsky, J.; Biddinger, D.J.; Joshi, N.K. Whole-Body Acute Contact Toxicity of Formulated Insecticide Mixtures to Blue Orchard Bees (Osmia lignaria). Toxics 2021, 9, 61. https://doi.org/10.3390/toxics9030061
Belsky J, Biddinger DJ, Joshi NK. Whole-Body Acute Contact Toxicity of Formulated Insecticide Mixtures to Blue Orchard Bees (Osmia lignaria). Toxics. 2021; 9(3):61. https://doi.org/10.3390/toxics9030061
Chicago/Turabian StyleBelsky, Joseph, David J. Biddinger, and Neelendra K. Joshi. 2021. "Whole-Body Acute Contact Toxicity of Formulated Insecticide Mixtures to Blue Orchard Bees (Osmia lignaria)" Toxics 9, no. 3: 61. https://doi.org/10.3390/toxics9030061
APA StyleBelsky, J., Biddinger, D. J., & Joshi, N. K. (2021). Whole-Body Acute Contact Toxicity of Formulated Insecticide Mixtures to Blue Orchard Bees (Osmia lignaria). Toxics, 9(3), 61. https://doi.org/10.3390/toxics9030061