
Effects of Acrylamide-Induced Vasorelaxation and Neuromuscular Blockage: A Rodent Study

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Assessment of Vasorelaxation
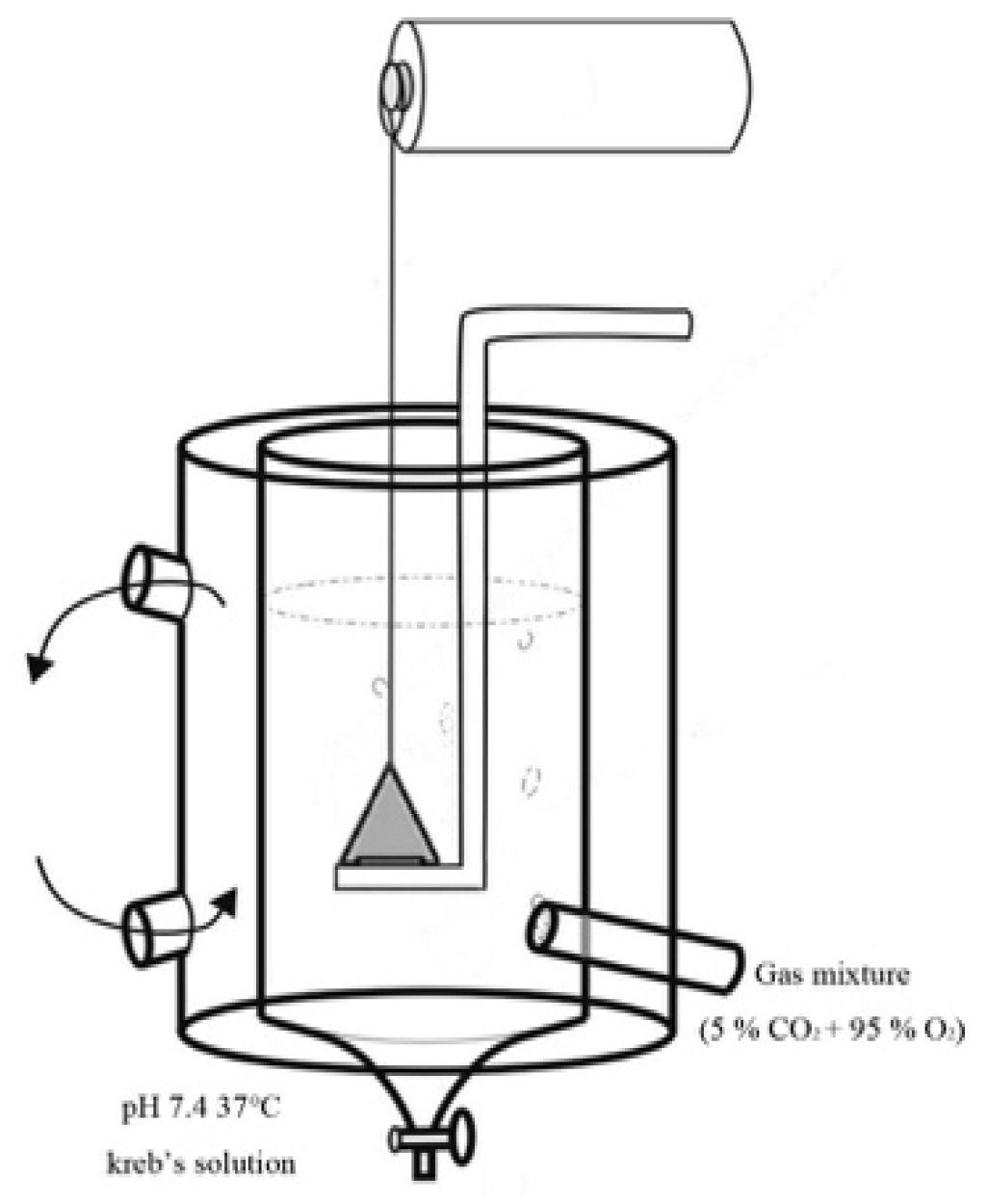
2.3.1. Rat Aortic Ring Preparation
2.3.2. Dosage Selection
2.3.3. Effects of ACR on Rat Aortic Rings
2.3.4. Effects of Nitric Oxide Synthase and AChR Inhibitors on the ACR-Relaxed Aortic Ring of Rats
2.3.5. Effects of ACR-Induced Relaxation with/without Ca2+
2.4. Assessment of Neuromuscular Toxicity
2.4.1. Preparation and Treatment of the Phrenic Nerve–Diaphragm Model
2.4.2. Effects of ACR on Phrenic Nerve–Diaphragm in Mice
2.4.3. Effects of AChR Inhibitor on ACR-Induced Phrenic Nerve–Diaphragm Changes
2.4.4. Effects of Extracellular Ca on ACR-Induced Phrenic Nerve–Diaphragm Changes
2.5. Statistical Analysis
3. Results
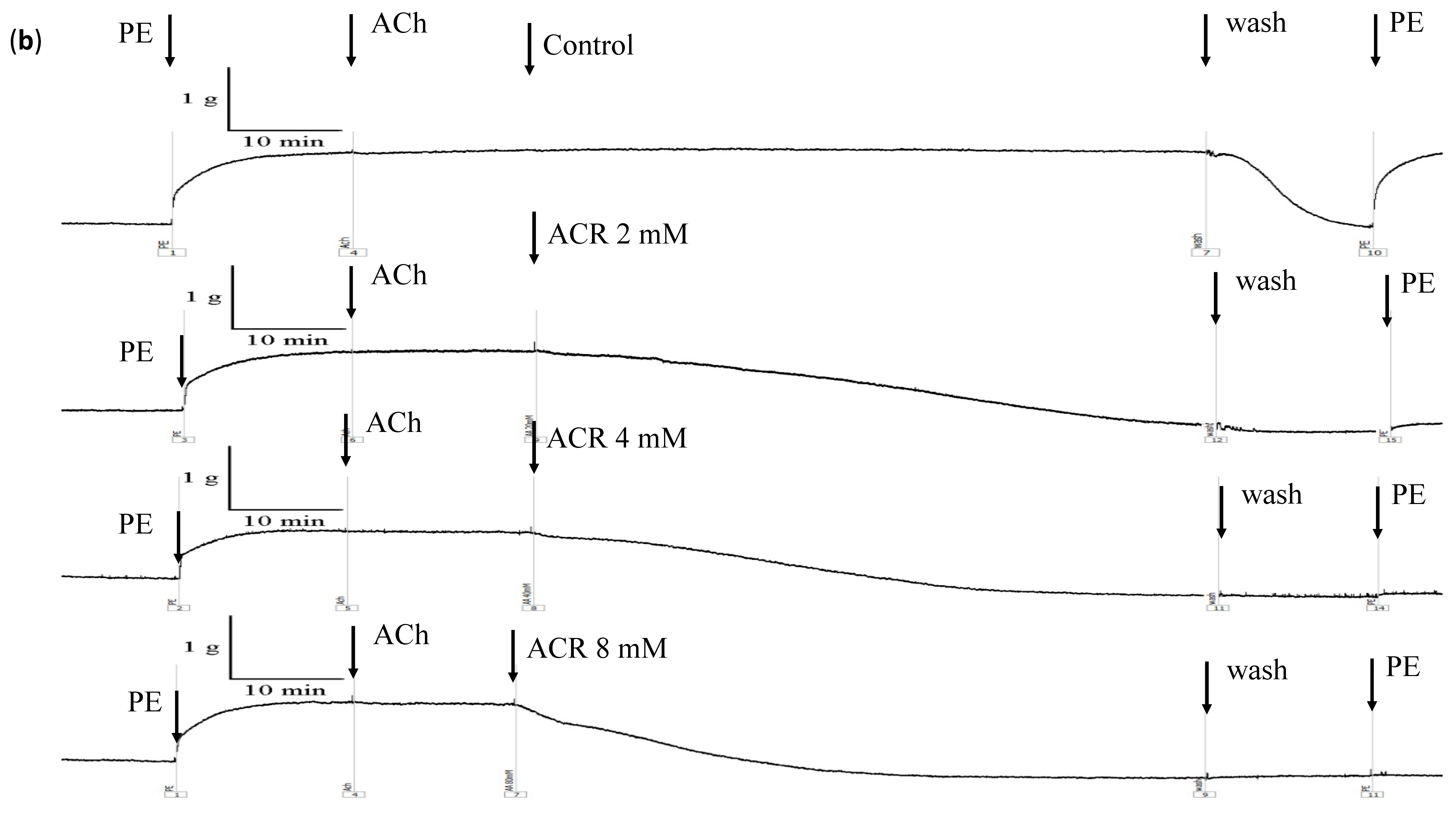
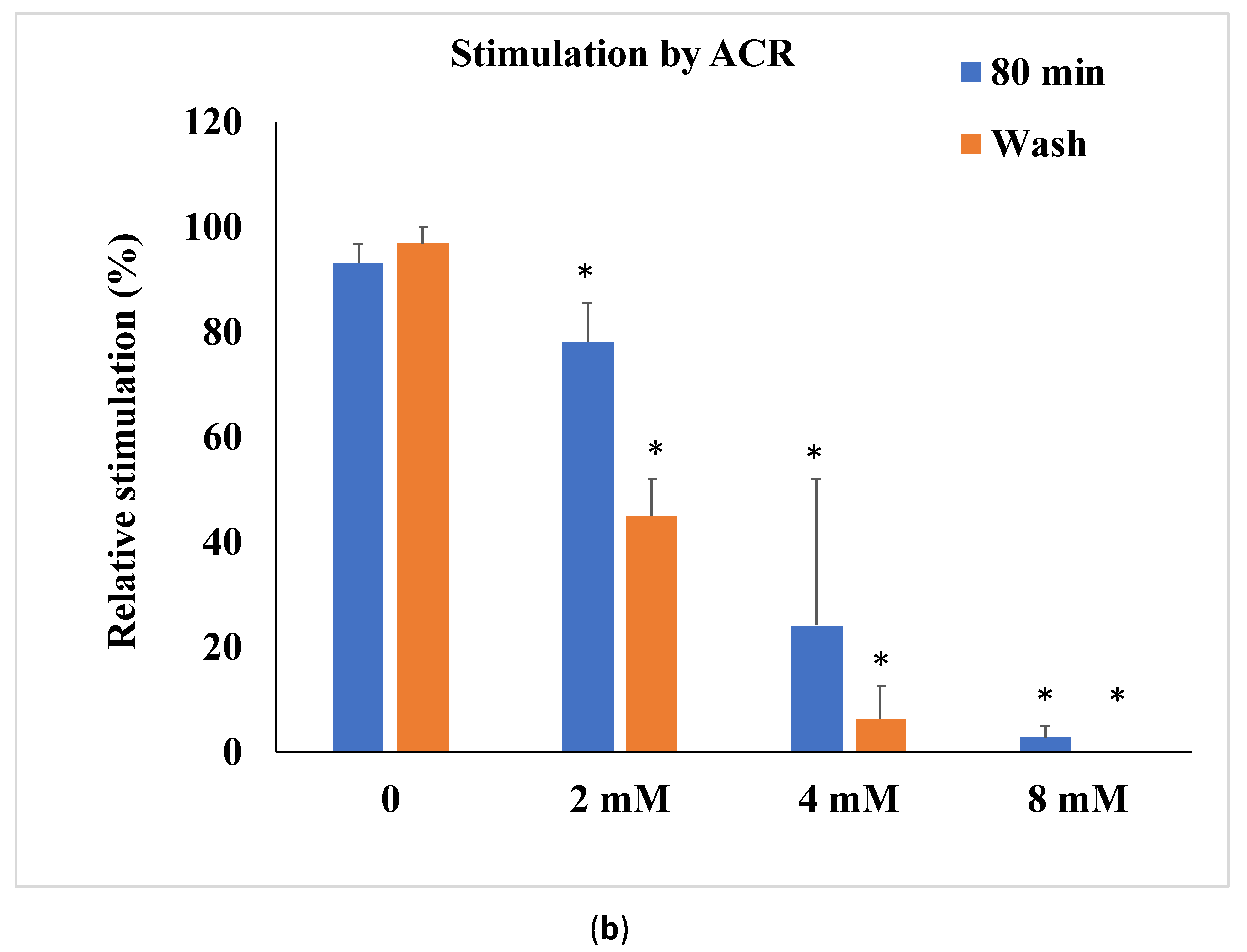
3.1. ACR-Induced Aortic Ring Vasorelaxation
3.2. Effects of L-NAME, Atropine, and Mecamylamine on ACR-Induced Vasorelaxation
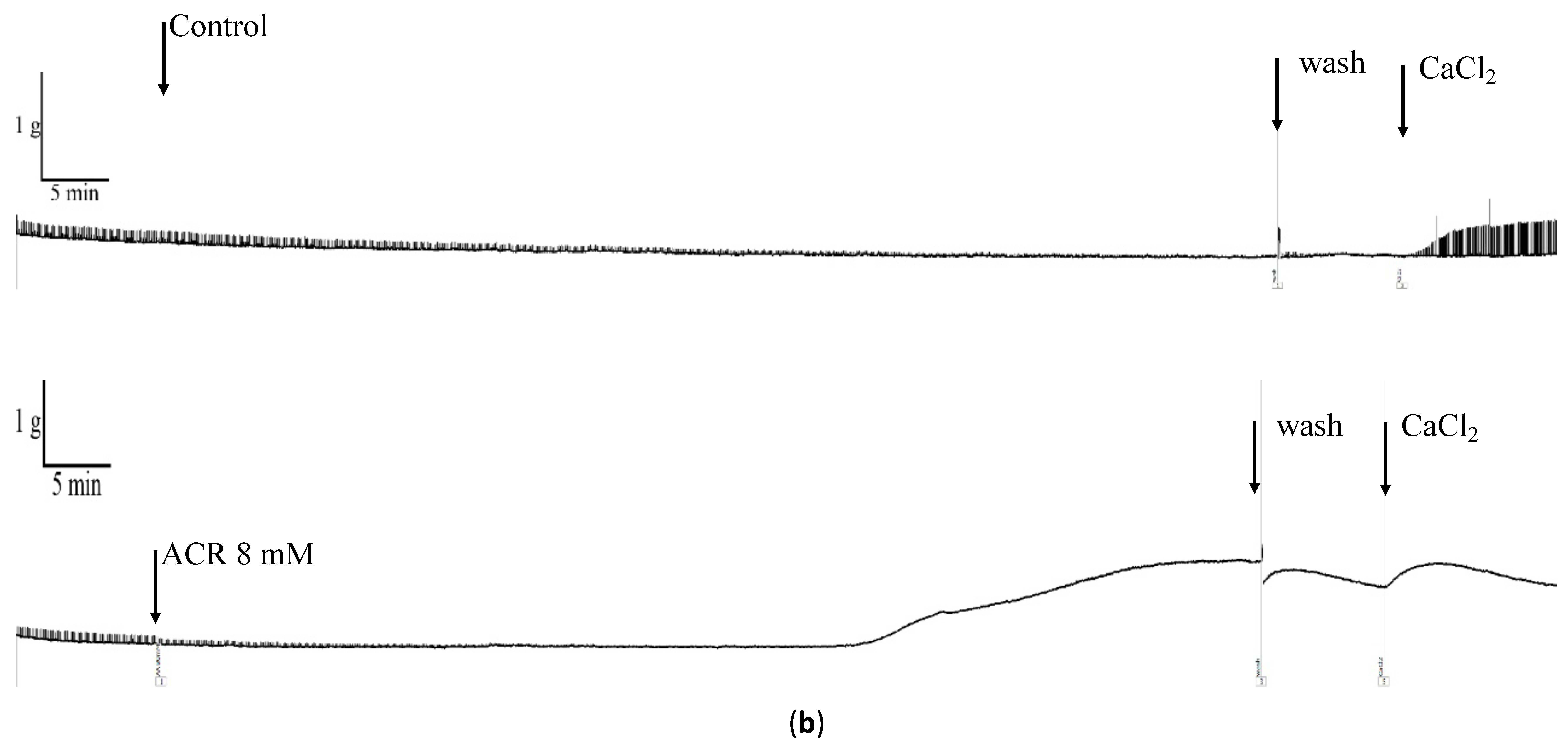
3.3. Effects of Ca2+ on ACR-Induced Vasorelaxation
3.4. Effects of ACR on Isolated Mouse Phrenic Nerve–Diaphragm
3.5. Effects of NTX on ACR-Induced Phrenic Nerve Diaphragm Contraction
3.6. Effects of Extracellular Ca2+ on ACR-Induced Changes in Phrenic Nerve–Diaphragm
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Rifai, L.; Saleh, F.A. A review on acrylamide in food: Occurrence, toxicity, and mitigation strategies. Int. J. Toxicol. 2020, 39, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.; Brabec, M.J.; Buelk-Sam, J.; Carlson, G.P.; Chapin, R.E.; Favor, J.B.; Fischer, L.J.; Hattis, D.; Lees, P.S.J.; Perreault-Darney, S.; et al. NTP-CERHR expert panel report on the repoductive and developmental toxicity of acrylamide. Birth Defects Res. B Dev. Reprod. Toxicol. 2005, 71, 17–113. [Google Scholar] [CrossRef] [PubMed]
- Erkekoglu, P.; Baydar, T. Acrylamide neurotoxicity. Nutr. Neurosci. 2014, 17, 49–57. [Google Scholar] [CrossRef]
- Exon, J.H. A Review of the Toxicology of Acrylamide. J. Toxicol. Environ. Health B Crit Rev. 2016, 9, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Krishna, G. Oral supplements of combined fructo-and xylo-oligosaccharides during perinatal period significantly offsets acrylamide-induced oxidative impairments and neurotoxicity in rats. J. Physiol. Pharmacol. 2018, 69, 801–814. [Google Scholar]
- Krishna, G.; Divyashri, G.; Prapulla, S.G. A combination supplement of fructo- and xylo-oligosaccharides significantly abrogates oxidative impairments and neurotoxicity in maternal/fetal milieu following gestational exposure to acrylamide in rat. Neurochem. Res. 2015, 40, 1904–1918. [Google Scholar] [CrossRef]
- Lo Pachin, R.M.; Gavin, T. Acrylamide-induced nerve terminal damage: Relevance to neurotoxic and neurodegenerative mechanisms. J. Agric. Food Chem. 2008, 56, 5994–6003. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abd Eldaim, M.A.; Hassan, A.G.A. Trigonella foenum-graecum ameliorates acrylamide-induced toxicity in rats: Role of oxidative stress, proinflammatory cytokines, and DNA damage. Biochem. Cell Biol. 2015, 93, 192–198. [Google Scholar] [CrossRef]
- Elhelaly, A.E.; AlBasher, G.; Alfarraj, S.; Almeer, R.; Bahbah, E.I.; Fouda, M.M.; Bungău, S.G.; Aleya, L.; Abdel-Daim, M.M. Protective effects of hesperidin and diosmin against acrylamide-induced liver, kidney, and brain oxidative damage in rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 35151–35162. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; El-Ela, F.I.A.; Alshahrani, F.K.; Bin-Jumah, M.; Al-Zharani, M.; Almutairi, B.; Alyousif, M.S.; Bungau, S.; Aleya, L.; Alkahtani, S. Protective effect of thymoquinone against acrylamide-induced liver, kidney and brain oxidative damage in rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 37709–37717. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Lu, P.C.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Association between acrylamide metabolites and cardiovascular risk in children with early stages of chronic kidney disease. Int. J. Mol. Sci. 2020, 21, 5855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, M.; Zhuang, P.; Jiao, J.; Chen, X.; Eang, J.; Wu, Y. Exposure to acrylamide and the risk of cardiovascular diseases in the National Health and Nurtition Examination survey 2003–2006. Environ. Int. 2018, 117, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Nurullahoglu-Atalik, E.; Okudan, N.; Belviranli, M.; Esen, H.; Yener, Y.; Oznurlu, Y. Acrylamide-treatment and responses to phenylephrine and potassium in rat aorta. Acta Physiol. Hung. 2012, 99, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Maynard, K.I.; Lincoln, J.; Milner, P.; Burnstock, G. Change in sympathetic and endothelium-mediated responses in the rabbit central ear artery after acrylamide treatment. J. Auton. Nerv. Syst. 1991, 36, 55–63. [Google Scholar] [CrossRef]
- Attoff, K.; Kertika, D.; Lundqvist, J.; Oredsson, S.; Forsby, A. Acrylamide affacted proliferation and differentiation of the neural progenitor cell line C17.2 and the neuroblastoma cell line SH-SY5Y. Toxicol. In Vitro 2016, 35, 100–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennisi, M.; Malaguarnera, G.; Puglisi, V.; Vinciguerra, L.; Vacante, M.; Malaguarnera, M. Neurotoxicity of acrylamide in exposed workers. Int. J. Environ. Res. Public Health 2013, 10, 3843–3854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radad, K.; El-Amir, Y.; Al-Eman, A.; Al-Shraim, M.; Bin-Jaliah, I.; Krewenka, C.; Moldzio, R. Minocycin protects against acrylamide-induced neurotoxicity and testicular damage in Sprague-Dawley rats. J. Toxicol. Pathol. 2020, 33, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.E.; Macun, I. Acrylamide neuropathy in a South African factory: An epidemiologic investigation. Am. J. Ind. Med. 1991, 19, 487–493. [Google Scholar] [CrossRef]
- Hogervorst, J.G.; Schouten, L.J.; Konings, E.J.; Goldbohm, R.A.; van den Brandt, P.A. A prospective study of. dietary acrylamide intake and the risk of endometrial, ovarian, and breast cancer. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 2304–2313. [Google Scholar] [CrossRef] [Green Version]
- Hogervorst, J.G.; Schouten, L.J.; Konings, E.J.; Goldbohm, R.A.; van den Brandt, P.A. Dietary acrylamide intake. and the risk of renal cells, bladder, and prostate cancer. Am. J. Clin. Nutr. 2008, 87, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Olesen, P.T.; Olsen, A.; Frandsen, H.; Frederiksen, K.; Overvad, K.; Tjonneland, A. Acrylamide exposure and. incidence of breast cancer among postmenopausal woman in the Danish Diet, Cancer and Health Study. Int. J. Cancer 2008, 122, 2094–2100. [Google Scholar] [CrossRef]
- Hogervorst, J.G.; Baars, B.J.; Schouten, L.J.; Konings, E.J.; Goldbohm, R.A.; van den Brandt, P.A. The carcinogenicity of dietary acrylamide intake: A comparative discussion of epidemiological and experimental animal research. Crit. Rev. Toxicol. 2006, 36, 481–608. [Google Scholar] [CrossRef] [PubMed]
- Shipp, A.; Lawrence, G.; Gentry, R.; McDonald, T.; Bartow, H.; Bounds, J.; Macdonald, N.; Clewell, H.; Allen, B.; Van Landingham, C. Acrylamide: Review of toxicity data and dose-response analyses for cancer and noncancer effects. Crit. Rev. Toxicol. 2010, 40, 485–512. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Brit. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. Osteoarthr. Cartil. 2012, 20, 256–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jespersen, B.; Tykocki, N.R.; Watts, S.W.; Cobbett, P.J. Measurement of smooth muscle function in the. isolated tissue bath-applications to pharmacology research. J. Vis. Exp. 2015, 95, 52324. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.T.; Hung, D.Z.; Chou, C.C.; Kang, J.J.; Cheng, Y.W.; Hu, C.M.; Liao, J.W. Vasorelaxation effects of 2-chloroethanol and chloroacetaldehyde in the isolated rat aortic rings. J. Health Sci. 2009, 55, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Paulsson, B.; Larsen, K.O.; Tornqvist, M. Hemoglobin adducts in the assessment of potential occupational exposure to acrylamides-three case studies. Scand. J. Work Environ. Health 2006, 32, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.W.; Kang, J.J.; Liu, S.H.; Jeng, C.R.; Cheng, Y.W.; Hu, C.M. Effects of Cartap on Isolated Mouse Phrenic Nerve Diaphragm and Its Related Mechanism. Toxicol. Sci. 2000, 55, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, G.; Negro, S.; Pirazzini, M.; Caccin, P. Mouse Phrenic Nerve Hemidiaphragm Assay (MPN). Bio Protoc. 2018, 8, e2759. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.; Griffin, S.; Emms, V.; Warren, N.; Cocker, J.; Farmer, P. Correlation of. hemoglobin-acrylamide adducts with air-borne exposure: An occupational survey. Toxicol. Lett. 2006, 162, 174–180. [Google Scholar] [CrossRef]
- Hagmar, L.; Tornqvist, M.; Nordander, C.; Rosen, I.; Bruze, M.; Kautiainen, A.; Magnusson, A.L.; Malmberg, B.; Aprea, P.; Granath, F.; et al. Health effects of occupational exposure to acrylamide using hemoglobin adducts as biomarkers of internal dose. Scand. J. Work Environ. Health 2001, 27, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calleman, C.J.; Wu, Y.; He, F.; Tian, G.; Bergmark, E.; Zhang, S.; Deng, H.; Wang, Y.; Crofton, K.M.; Fennell, T.; et al. Relationships between biomarkers of exposure and neurological effects in a group of workers exposed to acrylamide. Toxicol. Appl. Pharmacol. 1994, 126, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Lyn-Cook, L.E., Jr.; Tareke, E.; Word, B.; Starlard-Davenport, A.; Lyn-Cook, B.D.; Hammons, G.J. Food contaminant acrylamide increase expression of Cox-2 and nitric oxide synthase in breast epithelial cells. Toxicol. Ind. Health 2011, 27, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, M.; Mukosera, G.T.; Borchardt, D.; Li, Q.; Tipple, T.E.; Ishtiaq Ahmed, A.S.; Power, G.G.; Blood, A.B. L-NAME release nitric oxide and potentiates subsequent nitroglycerin-mediated vasodilation. Redox Biol. 2019, 26, 101238. [Google Scholar] [CrossRef]
- Wilson, C.; Lee, M.D.; McCarron, J.G. Acetylcholine released by endothelial cells facilitates flow-mediated dilatation. J. Physiol. 2016, 594, 7267–7307. [Google Scholar] [CrossRef]
- Nevala, R. Effects of Genistein and Daidzein on Arterial Tone and Blood Pressure in Rats; Helsinki Yliopistopaino: Helsinki, Finland, 2001; pp. 25–26. [Google Scholar]
- Ferreira, S.H.; Lorenzetti, B.B.; Cunha, F.Q.; Poole, S. Bradykinin release of TNF-alpha plays a key role in the development of inflammatory hyperalgesia. Agents Actions 1993, 38, C7–C9. [Google Scholar] [CrossRef]
- Tiffany, C.W.; Burch, R.M. Bradykinin stimulates tumor necrosis factor and interleukin-1 release from macrophages. FEBS Lett. 1989, 247, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Santhanasabapathy, R.; Vasudevan, S.; Anupriya, K.; Pabitha, R.; Sudhandiran, G. Farnesol quells oxidative stress, reactive gliosis and inflammation during acrylamide-induced neurotoxicity: Behavioral and biochemical evidence. Neuro Sci. 2015, 308, 212–227. [Google Scholar] [CrossRef]
- Kume, T.; Takada-Takatori, Y. Nicotinic Acetylcholine Receptor Signaling: Roles in Neuroprotection. In Nicotinic Acetylcholine Receptor Signaling in Neuroprotection; Springer: Singapore, 2018; pp. 59–71. [Google Scholar]
- Papke, R.; Sanberg, P.; Shytle, R. Analysis of mecamylamine stereoisomers on human nicotinic receptor subtypes. J. Pharmacol. Exp. Ther. 2001, 297, 646–656. [Google Scholar]
- Kopanska, M.; Lukac, N.; Kapusta, E.; Formicki, G. Acrylamide influence on activity of acetylcholinesterase, thiol groups, and malondialdehyde content in the brain of Swiss mice. J. Biochem. Mol. Toxicol. 2015, 29, 472–478. [Google Scholar] [CrossRef]
- Kopanska, M.; Muchacka, R.; Czech, J.; Batoryna, M.; Formicki, G. Acrylamide toxicity and cholinergic nervous system. J. Physiol. Pharmacol. 2018, 69, 847–858. [Google Scholar]
- Gaspersic, R.; Koritnik, B.; Crne-Finderle, N.; Sketelj, J. Acetylcholinesterase in the neuromuscular junction. Chem. Biol. Interact. 1999, 119, 301–308. [Google Scholar] [CrossRef]
- Couraud, J.Y.; Di Giamberardino, L.; Chretien, M.; Souyri, F.; Fardeau, M. Acrylamide neuropathy and changes in the axonal transport and muscular content of the molecular forms of acetylcholinesterase. Muscle Nerve 1982, 5, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.B.; Krajacic, P.; Sawamoto, K.; Bunya, N.; Loro, E.; Khurana, T.S. Pharmacotherapy to protect the neuromuscular junction after acute organophosphorus pesticide poisoning. Ann. N. Y. Acad. Sci. 2016, 1374, 86–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Yang, C.; Zheng, X.; Guan, Q.; Yu, S. Acrylamide treatment alters the level of Ca2+ and Ca2+ related protein kinase in spinal cords of rats. Toxicol. Ind. Health 2021, 37, 113–123. [Google Scholar] [CrossRef]
- Fleischer, S.; Inui, M. Biochemistry and Biophysics of Excitation-Contraction Coupling. Annu. Rev. Biophys. Biophys. Chem. 1989, 18, 333–364. [Google Scholar] [CrossRef] [PubMed]
- Erkekoglu, P.; Baydar, T. Toxicity of acrylamide and evaluation of its exposure in body foods. Nutr. Res. Rev. 2010, 23, 323–333. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | % of PE Precontracted 1 | |
|---|---|---|
| 10 min | 60 min | |
| Control | 22.8 ± 3.9 | 127.0 ± 6.7 |
| L-NAME | 4.2 ± 1.3 * | 114.3 ± 4.2 * |
| Atropine | 37.6 ± 7.4 * | 138.4 ± 15.4 |
| Mecamylamine | 19.7 ± 7.0 | 96.8 ± 1.8 * |
| Group | % of PE Precontracted 1 | |
|---|---|---|
| 10 min | 60 min | |
| Control | 170.0 ± 38.2 | 226.9 ± 53.3 |
| ACR | 231.5 ± 60.4 | −136.1 ± 49.7 * |
| Relative Stimulation (%) | Baseline Change (%) | |||
|---|---|---|---|---|
| Control | ACR | Control | ACR | |
| 80 min | 9.1 ± 5.1 | 0.5 ± 0.8 * | −19.4 ± 4.2 | −4.3 ± 6.7 * |
| wash | 76.7 ± 11.6 | 62.0 ± 9.8 | −21.4 ± 5.0 | −4.8 ± 8.2 * |
| Baseline Change (%) | ||
|---|---|---|
| Control | ACR | |
| 80 min | −16.9 ± 3.2 | 123.8 ± 86.7 |
| Wash | −16.2 ± 3.7 | 88.3 ± 46.2 * |
| Ca2+ | −13.4 ± 3.4 | 71.8 ± 9.5 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-D.; Ou, C.-C.; Hsiao, S.-H.; Chang, C.-H.; Tsai, F.-J.; Liao, J.-W.; Chen, Y.-T. Effects of Acrylamide-Induced Vasorelaxation and Neuromuscular Blockage: A Rodent Study. Toxics 2021, 9, 117. https://doi.org/10.3390/toxics9060117
Lin W-D, Ou C-C, Hsiao S-H, Chang C-H, Tsai F-J, Liao J-W, Chen Y-T. Effects of Acrylamide-Induced Vasorelaxation and Neuromuscular Blockage: A Rodent Study. Toxics. 2021; 9(6):117. https://doi.org/10.3390/toxics9060117
Chicago/Turabian StyleLin, Wei-De, Chu-Chyn Ou, Shih-Hao Hsiao, Chih-Han Chang, Fuu-Jen Tsai, Jiunn-Wang Liao, and Yng-Tay Chen. 2021. "Effects of Acrylamide-Induced Vasorelaxation and Neuromuscular Blockage: A Rodent Study" Toxics 9, no. 6: 117. https://doi.org/10.3390/toxics9060117
APA StyleLin, W. -D., Ou, C. -C., Hsiao, S. -H., Chang, C. -H., Tsai, F. -J., Liao, J. -W., & Chen, Y. -T. (2021). Effects of Acrylamide-Induced Vasorelaxation and Neuromuscular Blockage: A Rodent Study. Toxics, 9(6), 117. https://doi.org/10.3390/toxics9060117