
One-Pot Ionic Liquid-Mediated Bioprocess for Pretreatment and Enzymatic Hydrolysis of Rice Straw with Ionic Liquid-Tolerance Bacterial Cellulase

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Biomass Preparations
2.2. Lignocellulose Pretreatment and Enzymatic Saccharification
2.3. FT-IR Spectroscopy Analysis
2.4. Preparation of IL-Tolerant Cellulase
2.5. SDS-PAGE and Zymogram Analysis
2.6. Cellulase Enzyme Assay
2.7. IL Toxicity Test to Bacteria Cells
2.8. One-Pot Process Set Up
2.9. Statistical Analysis
3. Results and Discussion
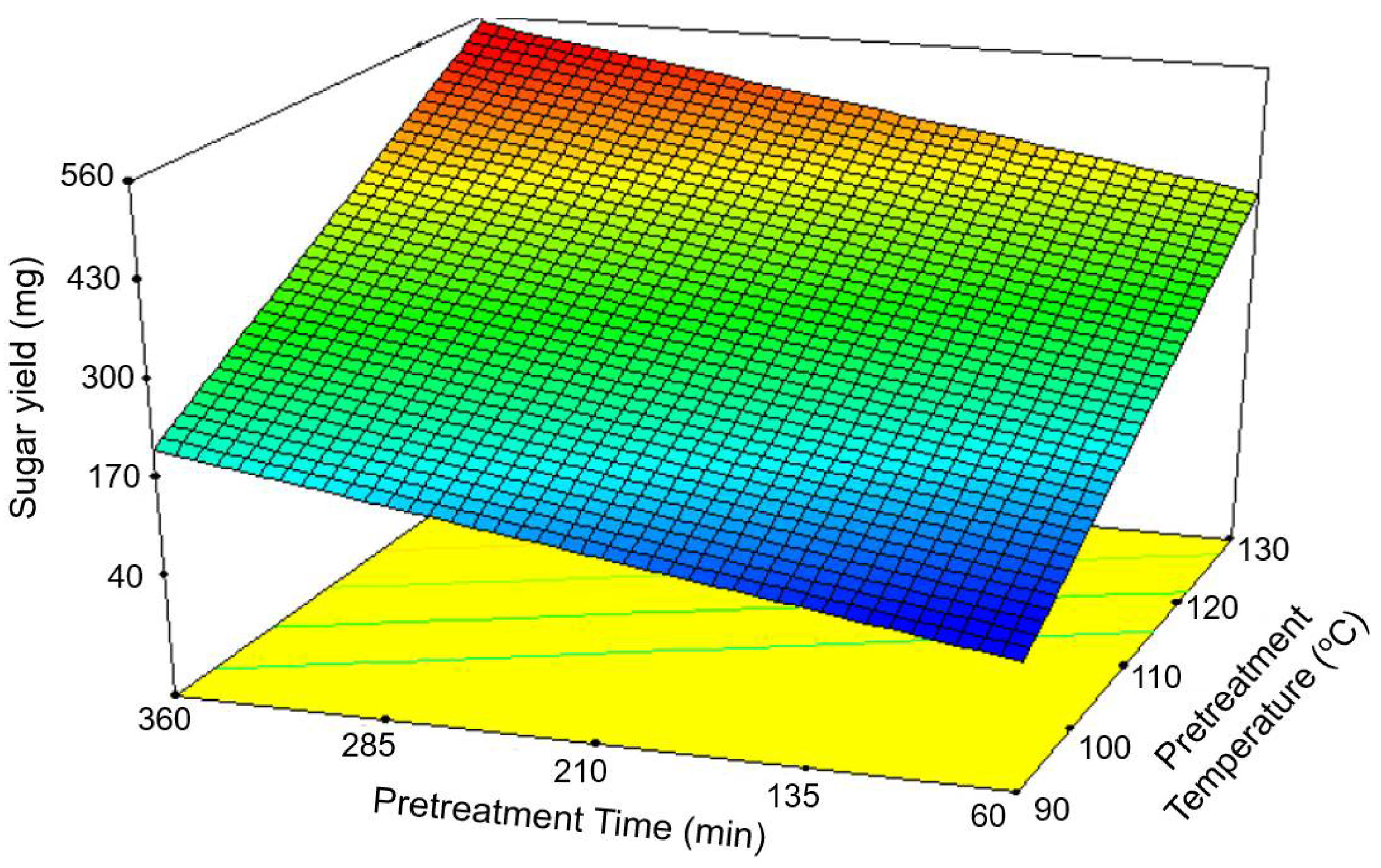
3.1. Optimization of [Ch][OAc] Pretreatment
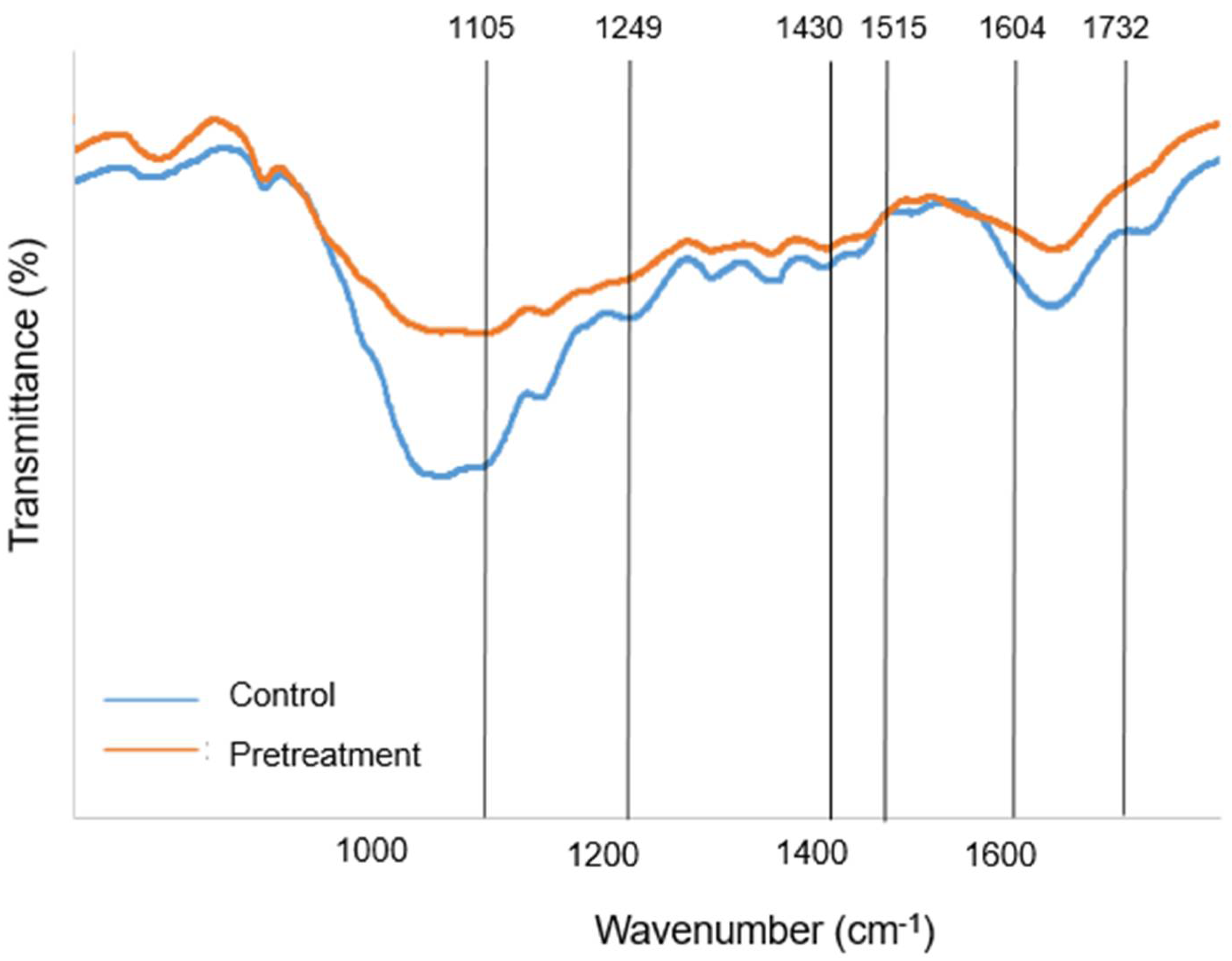
3.2. Modifications of Lignocellulose Biomass by [Ch][OAc] Pretreatment
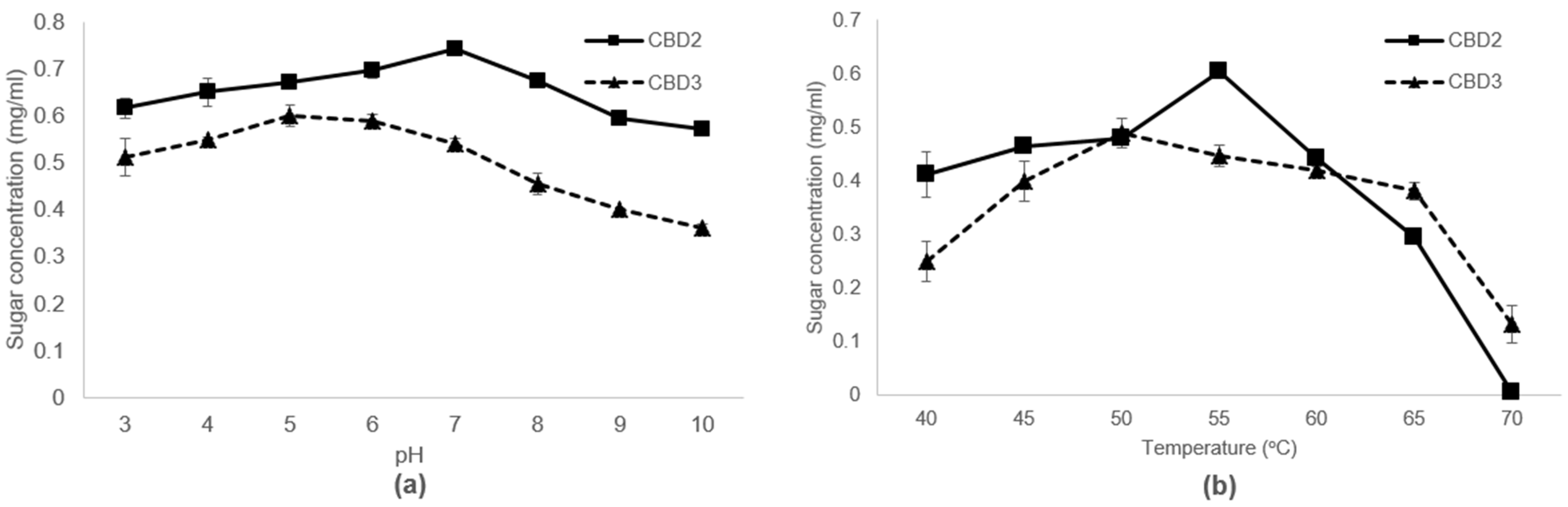
3.3. Characterization of Cellulases Produced by Bacterial Strains Isolated from Saline Soil Samples
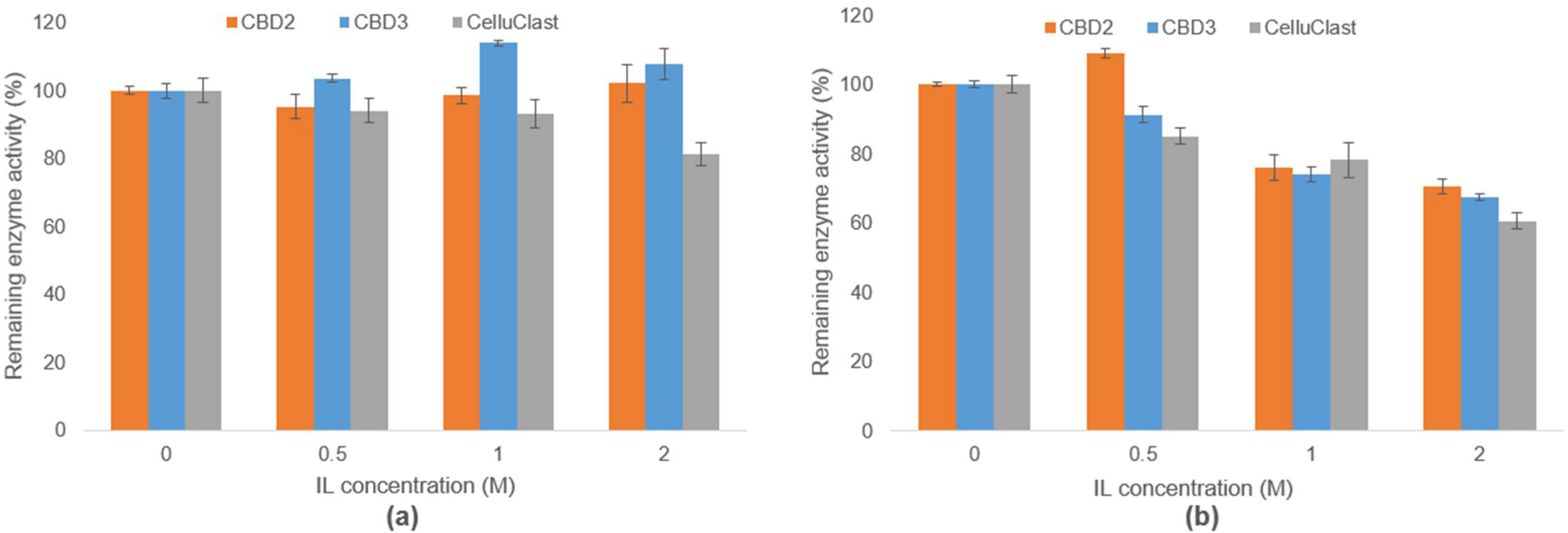
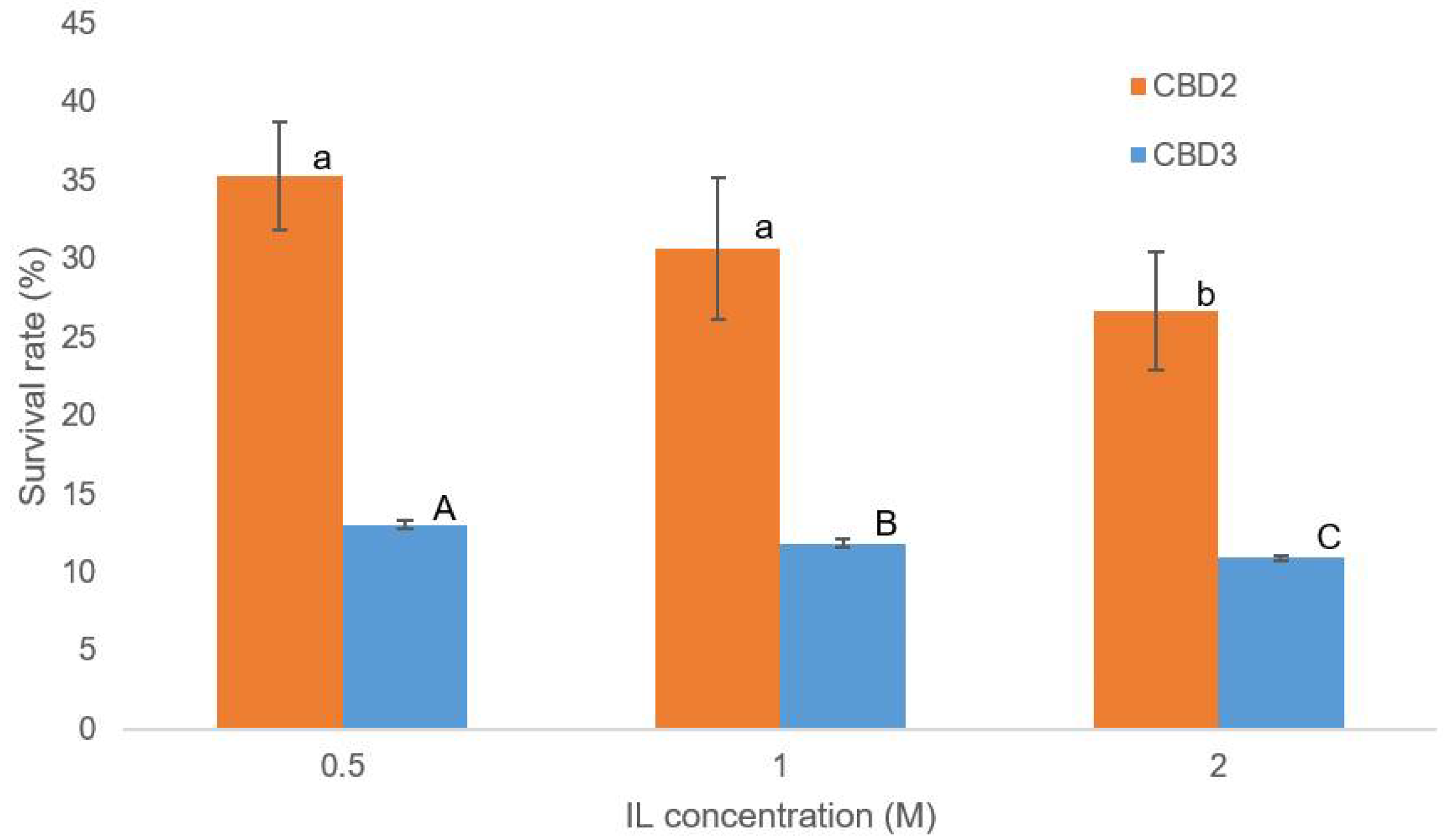
3.4. Evaluation of IL-Tolerance Properties of Bacterial Cellulases
3.5. Evaluation of IL-Tolerance Properties of Bacterial Cultures
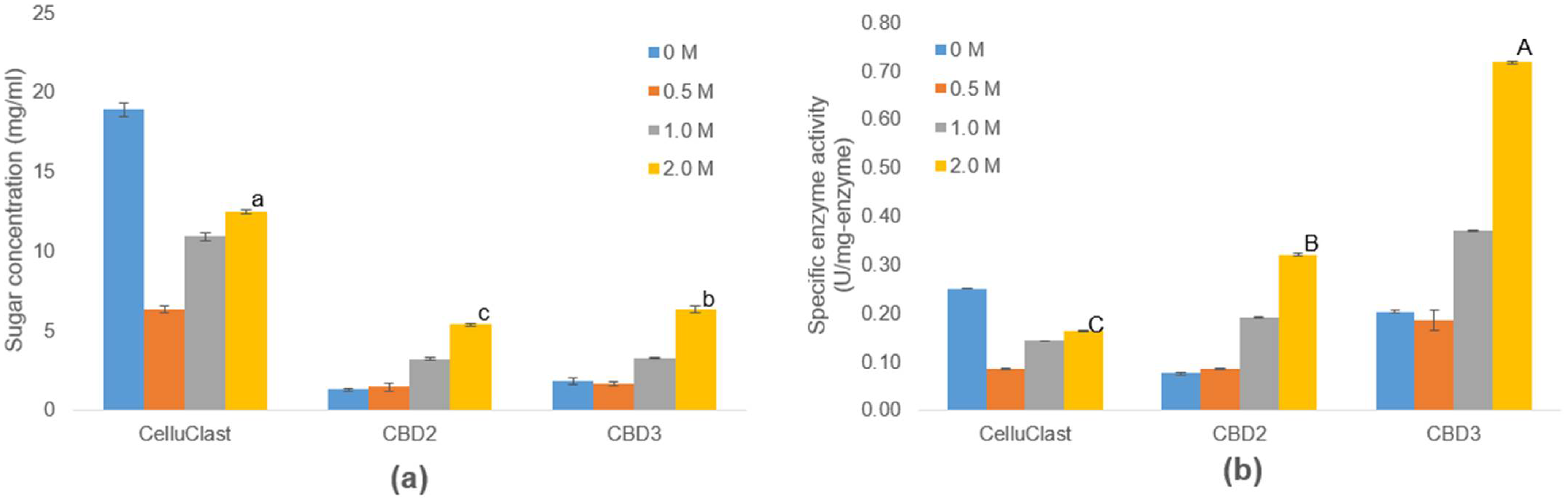
3.6. Efficiency of IL-Tolerance Cellulases in One-Pot Process
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, Y.S.; Mutrakulcharoen, P.; Chuetor, S.; Cheenkachorn, K.; Tantayotai, P.; Panakkal, E.J.; Sriariyanun, M. Recent situation and progress in biorefining process of lignocellulosic biomass: Toward green economy. Appl. Sci. Eng. Prog. 2020, 13, 299–311. [Google Scholar] [CrossRef]
- Capolupo, L.; Faraco, V. Green methods of lignocellulose pretreatment for biorefinery development. Appl. Microbiol. Biotechnol. 2016, 100, 9451–9467. [Google Scholar] [CrossRef] [Green Version]
- Rattanaporn, K.; Tantayotai, P.; Phusantisampan, T.; Pornwongthong, P.; Sriariyanun, M. Organic acid pretreatment of oil palm trunk: Effect on enzymatic saccharification and ethanol production. Bioprocess. Biosyst. Eng. 2018, 41, 467–477. [Google Scholar] [CrossRef]
- Amnuaycheewa, P.; Hengaroonprasan, R.; Rattanaporn, K.; Kirdponpattara, S.; Cheenkachorn, K.; Sriariyanun, M. Enhancing enzymatic hydrolysis and biogas production from ricestraw by pretreatment with organic acids. Ind. Crops. Prod. 2016, 84, 247–254. [Google Scholar] [CrossRef]
- Karimi, K.; Taherzadeh, M.J. A critical review of analytical methods in pretreatment of lignocelluloses: Composition, imaging, and crystallinity. Bioresour. Technol. 2016, 200, 1008–1018. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, W.C.; Li, F.X.; Yu, J.Y. Swelling and dissolution of cellulose in NaOH aqueous solvent systems. Cellul. Chem. Technol. 2013, 47, 671–679. [Google Scholar]
- Welton, T. Ionic liquid: A brief history. Biophys. Rev. 2018, 10, 691–706. [Google Scholar] [CrossRef] [Green Version]
- Gundupalli, M.P.; Rattanaporn, K.; Chuetor, S.; Rodiahwati, W.; Sriariyanun, M. Biocomposite production from Ionic liquids (IL) assisted processes using biodegradable biomass. In Toward the Value-Added Biocomposites: Technology, Innovation and Opportunity; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar] [CrossRef]
- Usmani, Z.; Sharma, M.; Gupta, P.; Karpichev, Y.; Gathergood, N.; Bhat, R.; Gupta, V.K. Ionic liquid based pretreatment of lignocellulosic biomass for enhanced bioconversion. Bioresour. Technol. 2020, 304, 123003. [Google Scholar] [CrossRef]
- Xu, H.; Peng, J.; Kong, Y.; Liu, Y.; Su, Z.; Li, B.; Song, X.; Liu, S.; Tian, W. Key process parameters for deep eutectic solvents pretreatment of lignocellulosic biomass materials: A review. Bioresour. Technol. 2020, 310, 123416. [Google Scholar] [CrossRef]
- Verma, C.; Mishra, A.; Chauhan, S.; Verma, P.; Srivastava, V.; Quraishi, M.; Ebenso, E.E. Dissolution of cellulose in ionic liquids and their mixed cosolvents: A review. Sustain. Chem. Pharm. 2019, 13, 100162. [Google Scholar] [CrossRef]
- Sriariyanun, M.; Yan, Q.; Nowik, I.; Cheenkachorn, K.; Phusantisampan, T.; Modigell, M. Efficient pretreatment of rice straw by combination of screw press and ionic liquid to enhance enzymatic hydrolysis. Kasetsart J. (Nat. Sci.) 2015, 49, 146–154. [Google Scholar]
- Bian, J.; Peng, F.; Peng, X.P.; Xiao, X.; Peng, P.; Xu, F.; Sun, R.C. Effect of [Emim]Ac pretreatment on the structure and enzymatic hydrolysis of sugarcane bagasse cellulose. Carbohydr. Polym. 2014, 100, 211–217. [Google Scholar] [CrossRef]
- Hamidh, U.; Arakawa, T.; H’ng, Y.Y.; Nakagawa-izumi, A.; Kishino, M. Recycled ionic liquid 1-ethyl-3-methylimidazolium acetate pretreatment for enhancing enzymatic saccharification of softwood without cellulose regeneration. J. Wood. Sci. 2018, 64, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Cheenkachorn, K.; Douzou, T.; Roddecha, S.; Tantayotai, P.; Sriariyanun, M. Enzymatic saccharification of rice straw under influence of recycled ionic liquid pretreatments. Energy Procedia 2016, 100, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Ninomiya, K.; Utami, A.R.I.; Tsuge, Y.; Kuroda, K.; Ogino, C.; Taima, T.; Saito, J.; Kimizu, M.; Takahashi, K. Pretreatment of bagasse with a minimum amount of cholinium ionic liquid for subsequent saccharification at high loading and co-fermentation for ethanol production. Chem. Eng. Sci. 2018, 334, 657–663. [Google Scholar] [CrossRef]
- Goncalves, A.R.T.; Paredes, X.; Cristino, A.F.; Santos, F.J.V.; Queiros, C. Ionic liquids—A review of their toxicity to living organisms. Int. J. Mol. Sci. 2021, 22, 5612. [Google Scholar] [CrossRef]
- Tantayotai, P.; Gundupalli, M.P.; Panakkal, E.J.; Sriariyanun, M.; Rattanaporn, K.; Bhattacharyya, D. Differential influence of imidazolium ionic liquid on cellulase kinetics in Saccharification of cellulose and lignocellulosic biomass substrate. Appl. Sci. Eng. Prog. 2022, 15, 5510. [Google Scholar] [CrossRef]
- Beil, S.; Markiewicz, M.; Pereira, C.S.; Stepnowski, P.; Thöming, J.; Stolte, S. Toward the proactive design of sustainable chemicals: Ionic liquids as a prime example. Chem. Rev. 2021, 121, 13132–13173. [Google Scholar] [CrossRef] [PubMed]
- Konda, N.M.; Shi, J.; Singh, S.; Blanch, H.W.; Simmons, B.A.; Klien-Marcuschamer, D. Understanding cost drivers and economic potential of two variants of ionic liquid pretreatment for cellulosic biofuel production. Biotechnol. Biofuels 2014, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Van Soest, P.; Wine, R. Use of detergents in the analysis of fibrous feeds. IV: Determination of plant cell-wall constituents. J. Assoc. Off. Anal. Chem. 1967, 50, 50–55. [Google Scholar]
- Sriariyanun, M.; Tantayotai, P.; Yasurin, P.; Pornwongthong, P.; Cheenkachorn, K. Production, purification and characterization of an ionic liquid tolerant cellulase from Bacillus sp. isolated from rice paddy field soil. Elec. J. Biotechnol. 2016, 19, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Mood, S.H.; Golfeshan, A.H.; Tabatabaei, M.; Abbasalizadeh, S.; Ardjmand, M. Comparison of different ionic liquids pretreatment for barley straw enzymatic saccharification. 3 Biotech 2013, 3, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Brandt-Talbot, A.; Gschwend, F.; Fennell, P.; Lammens, T.; Tan, B.; Weale, J.; Hallett, J. An economically viable ionic liquid for the fractionation of lignocellulose biomass. Green Chem. 2017, 19, 3078–3102. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Simmons, B.A. Ionic liquid pretreatment: Mechanism, performance, and challenges. In Aqueous Pretreatment of Plant Biomass for Biological and Bhemical Bonversion to Fuels and Chemicals; Stevens, C.V., Wyman, C.E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Namnuch, N.; Thammasittirong, A.; Thammasittirong, S.N. Lignocellulose hydrolytic enzymes production by Aspergillus flavus KUB2 using submerged fermentation of sugarcane bagasse waste. Mycology 2020, 12, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Gundupalli, M.P.; Cheng, Y.S.; Chuetor, S.; Bhattacharyya, D.; Sriariyanun, M. Effect of dewaxing on saccharification and ethanol production from different lignocellulosic biomass. Bioresour. Technol. 2021, 339, 125596. [Google Scholar] [CrossRef]
- Ping, L.; You, L.H. Preparation and characterization of cellulose nanocrystals from rice straw. Carbohyd. Polym. 2012, 87, 564–573. [Google Scholar]
- Traoré, M.; Kaal, J.; Cortizas, M.A. Differentiation between pine woods according to species and growing location using FTIR-ATR. Wood. Sci. Technol. 2018, 52, 487–504. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.L.; O’Conner, R.T. Relation of certain infrared bands to cellulose crystallinity and crystal lattice type. Part II. A new infrared ratio for estimation of crystallinity in celluloses I and II. J. Appl. Polym. Sci. 1964, 8, 1325–1341. [Google Scholar] [CrossRef]
- Tantayotai, P.; Rattanaporn, K.; Tepaamorndech, S.; Cheenkachorn, K.; Sriariyanun, M. Analysis of an ionic liquid and salt tolerant microbial consortium which is useful for enhancement of enzymatic hydrolysis and biogas production. Waste Biomass Valori. 2019, 10, 1481–1491. [Google Scholar] [CrossRef]
- Pal, S.; Sar, A.; Dam, B. Moderate halophilic bacteria, but not extreme halophilic archaea can alleviate the toxicity of short-alkyl side chain imidazolium-based ionic liquids. Ecotoxicol. Environ. Saf. 2019, 184, 109634. [Google Scholar] [CrossRef]
- Megaw, J.; Busetti, A.; Gilmore, B.F. Isolation and characterisation of 1-alkyl-3-methylimidazolium chloride ionic liquid-tolerant and biodegrading marine bacteria. PLoS ONE 2013, 8, e60806. [Google Scholar]
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Fava, F. Halo-alkalitolerant and thermostable cellulases with improved tolerance to ionic liquids and organic solvents from Paenibacillus tarimensis isolated from the Chott El Fejej, Sahara desert, Tunisia. Biores. Technol. 2013, 150, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Azadian, F.; Badoei-Dalfard, A.; Namaki-Shoushtari, A.; Hassanshahian, M. Purification and biochemical properties of a thermostable, haloalkaline cellulase from Bacillus licheniformis AMF-07 and its application for hydrolysis of different cellulosic substrates to bioethanol production. Mol. Biol. Res. Commun. 2016, 5, 143–155. [Google Scholar] [PubMed]
- Maiti, S.; Samanta, T.; Sahoo, S.; Roy, S. The Dual carboxymethyl cellulase and gelatinase activities of a newly isolated protein from Brevibacillus agri ST15c10 confer reciprocal regulations in substrate utilization. J. Mol. Microbiol. Biotechnol. 2017, 27, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, G.; Muppidi, G.L.; Gurram, R.N.; Adhikari, A.; Bischoff, K.M.; Hughes, S.R.; Apel, W.A.; Bang, S.S.; Dixon, D.J.; Sani, R.K. Isolation and characterization of cellulose-degrading bacteria from the deep subsurface of the Homestake gold mine, Lead, South Dakota, USA. J. Ind. Microbiol. Biotechnol. 2009, 36, 585–598. [Google Scholar] [CrossRef]
- Fu, D.; Mazza, G. Aqueous ionic liquid pretreatment of straw. Bioresour. Technol. 2011, 102, 7008–7011. [Google Scholar] [CrossRef]
- Pedersen, J.N.; Pérez, B.; Guo, Z. Stability of cellulase in ionic liquids: Correlations between enzyme activity and COSMO-RS descriptors. Sci. Rep. 2019, 9, 17479. [Google Scholar] [CrossRef] [Green Version]
- Minami, S.; Yusaku, O.; Shiori, A.; Keita, F.; Tomoyoshi, M.; Yukihiro, Y.; Akio, S. Effect of ionic liquids on the hatching of Artemia salina cysts. Aust. J. Chem. 2018, 71, 492–496. [Google Scholar] [CrossRef]
- Ventura, S.P.M.; de Silva, F.A.; Gonçalves, A.M.M.; Pereira, J.L.; Gonçalves, F.; Coutinho, J.L.P. Ecotoxicity analysis of cholinium-based ionic liquids to Vibrio fischeri marine bacteria. Ecotoxicol. Environ. Saf. 2014, 102, 48–54. [Google Scholar] [CrossRef]
- Gundupalli, M.P.; Sahithi, A.S.T.; Cheng, Y.S.; Tantayotai, P.; Sriariyanun, M. Differential effects of inorganic salts on cellulase kinetics in enzymatic saccharification of cellulose and lignocellulosic biomass. Bioprocess. Biosyst. Eng. 2021, 44, 2331–2344. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Pretreatment Temperature (°C) | Pretreatment Time (min) | Loading Ratio (%wt) | Sugar Yield (mg/g-Biomass) |
|---|---|---|---|---|
| 1 | 90 | 60 | 10 | 120.47 |
| 2 | 90 | 210 | 5 | 145.46 |
| 3 | 90 | 210 | 15 | 138.06 |
| 4 | 90 | 360 | 10 | 217.98 |
| 5 | 110 | 60 | 5 | 159.76 |
| 6 | 110 | 60 | 15 | 176.04 |
| 7 | 110 | 210 | 10 | 287.54 |
| 8 | 110 | 210 | 10 | 294.45 |
| 9 | 110 | 210 | 10 | 340.33 |
| 10 | 110 | 210 | 10 | 335.89 |
| 11 | 110 | 210 | 10 | 291.49 |
| 12 | 110 | 360 | 5 | 362.04 |
| 13 | 110 | 360 | 15 | 408.91 |
| 14 | 130 | 60 | 10 | 431.60 |
| 15 | 130 | 210 | 5 | 512.02 |
| 16 | 130 | 210 | 15 | 496.23 |
| 17 | 130 | 360 | 10 | 528.30 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Model | 3.014 × 105 | 2 | 1.507 × 105 | 146.21 | <0.0001 |
| A-Temp | 2.437 × 105 | 1 | 2.437 × 105 | 236.44 | <0.0001 |
| B-Time | 57,690.11 | 1 | 57,690.11 | 55.98 | <0.0001 |
| Residual | 14,427.98 | 14 | 1030.57 | ||
| Lack of Fit | 11,748.81 | 10 | 1174.88 | 1.75 | 0.3095 |
| Pure Error | 2679.17 | 4 | 669.79 | ||
| Cor Total | 3.158 × 105 | 16 |
| Sample | Sugar Yield (mg/g-Biomass) | Composition (%) | ||||
|---|---|---|---|---|---|---|
| Predicted | Experimental | Error (%) | Cellulose | Hemicellulose | Lignin | |
| Control | N.A. | 90.5 | N.A. | 40.52 ± 0.32 | 32.70 ± 0.13 | 14.95 ± 0.20 |
| Pretreat | 542.3 | 551.2 | 1.64 | 45.84 ± 0.64 | 34.03 ± 0.17 | 8.39 ± 0.26 |
| Predicted Model: Sugar yield (mg/g-biomass) = −773.09380 + 8.72626 × Time + 0.56613 × Time Optimal condition: Pretreatment temperature, 129.21 °C; Pretreatment time, 331.82 min; Loading ratio, 10.68% | ||||||
| Enzyme | Optimal Condition | Enzyme Activity (U/mL) | Specific Enzyme Activity (U/mg) |
|---|---|---|---|
| CBD2 | pH 7, 55 °C | 0.62 ± 0.023 c | 0.20 ± 0.014 C |
| CBD3 | pH 5, 50 °C | 0.73 ± 0.017 b | 0.22 ± 0.018 B |
| Celluclast 1.5 L | pH 5, 50 °C | 4.20 ± 0.033 a | 0.32 ± 0.025 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sriariyanun, M.; Kitiborwornkul, N.; Tantayotai, P.; Rattanaporn, K.; Show, P.-L. One-Pot Ionic Liquid-Mediated Bioprocess for Pretreatment and Enzymatic Hydrolysis of Rice Straw with Ionic Liquid-Tolerance Bacterial Cellulase. Bioengineering 2022, 9, 17. https://doi.org/10.3390/bioengineering9010017
Sriariyanun M, Kitiborwornkul N, Tantayotai P, Rattanaporn K, Show P-L. One-Pot Ionic Liquid-Mediated Bioprocess for Pretreatment and Enzymatic Hydrolysis of Rice Straw with Ionic Liquid-Tolerance Bacterial Cellulase. Bioengineering. 2022; 9(1):17. https://doi.org/10.3390/bioengineering9010017
Chicago/Turabian StyleSriariyanun, Malinee, Nichaphat Kitiborwornkul, Prapakorn Tantayotai, Kittipong Rattanaporn, and Pau-Loke Show. 2022. "One-Pot Ionic Liquid-Mediated Bioprocess for Pretreatment and Enzymatic Hydrolysis of Rice Straw with Ionic Liquid-Tolerance Bacterial Cellulase" Bioengineering 9, no. 1: 17. https://doi.org/10.3390/bioengineering9010017
APA StyleSriariyanun, M., Kitiborwornkul, N., Tantayotai, P., Rattanaporn, K., & Show, P. -L. (2022). One-Pot Ionic Liquid-Mediated Bioprocess for Pretreatment and Enzymatic Hydrolysis of Rice Straw with Ionic Liquid-Tolerance Bacterial Cellulase. Bioengineering, 9(1), 17. https://doi.org/10.3390/bioengineering9010017