
A Novel CARMIL2 Immunodeficiency Identified in a Subset of Cavalier King Charles Spaniels with Pneumocystis and Bordetella Pneumonia


 ,
,  , , ,
, , ,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. CARMIL2 Variant Discovery
2.2. Phenotype Cohorts
2.3. CARMIL2 Genotyping
3. Results
3.1. Whole Canine Genome Sequencing and Variant Filtering
3.2. CARMIL2 Genotyping
3.3. Clinical Summary for CKCSs Homozygous for the CARMIL2 Variant Allele
3.4. Clinical Summary for CKCSs with Suspected PI Clear of the CARMIL2 Variant Allele
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aliouat-Denis, C.-M.; Chabé, M.; Demanche, C.; Aliouat, E.M.; Viscogliosi, E.; Guillot, J.; Delhaes, L.; Dei-Cas, E. Pneumocystis Species, Co-Evolution and Pathogenic Power. Infect. Genet. Evol. 2008, 8, 708–726. [Google Scholar] [CrossRef]
- Sokulska, M.; Kicia, M.; Wesołowska, M.; Hendrich, A.B. Pneumocystis Jirovecii—From a Commensal to Pathogen: Clinical and Diagnostic Review. Parasitol. Res. 2015, 114, 3577–3585. [Google Scholar] [CrossRef]
- Morris, A.; Norris, K.A. Colonization by Pneumocystis Jirovecii and Its Role in Disease. Clin. Microbiol. Rev. 2012, 25, 297–317. [Google Scholar] [CrossRef] [PubMed]
- Fillatre, P.; Decaux, O.; Jouneau, S.; Revest, M.; Gacouin, A.; Robert-Gangneux, F.; Fresnel, A.; Guiguen, C.; Le Tulzo, Y.; Jégo, P.; et al. Incidence of Pneumocystis Jiroveci Pneumonia among Groups at Risk in HIV-Negative Patients. Am. J. Med. 2014, 127, 1242.e11–1242.e17. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Chattaraj, A.; Iqbal, Q.; Anjum, A.; Rehman, M.E.U.; Aijaz, Z.; Nasir, F.; Ansar, S.; Zangeneh, T.T.; Iftikhar, A. Pneumocystis Jiroveci Pneumonia: A Review of Management in Human Immunodeficiency Virus (HIV) and Non-HIV Immunocompromised Patients. Avicenna J. Med. 2023, 13, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Danesi, P.; Petini, M.; Falcaro, C.; Bertola, M.; Mazzotta, E.; Furlanello, T.; Krockenberger, M.; Malik, R. Pneumocystis Colonization in Dogs Is as in Humans. Int. J. Environ. Res. Public Health 2022, 19, 3192. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.J.; Wotton, P.; Eastwood, J.; Swift, S.T.; Jones, B.; Day, M.J. Immunoglobulin Deficiency in Cavalier King Charles Spaniels with Pneumocystis Pneumonia. J. Vet. Intern. Med. 2006, 20, 523–527. [Google Scholar] [CrossRef]
- Weissenbacher-Lang, C.; Fuchs-Baumgartinger, A.; Guija-De-Arespacochaga, A.; Klang, A.; Weissenböck, H.; Künzel, F. Pneumocystosis in Dogs: Meta-Analysis of 43 Published Cases Including Clinical Signs, Diagnostic Procedures, and Treatment. J. Vet. Diagn. Investig. 2018, 30, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Lobetti, R. Common Variable Immunodeficiency in Miniature Dachshunds Affected with Pneumonocystis Carinii Pneumonia. J. Vet. Diagn. Investig. 2000, 12, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Shearin, A.L.; Ostrander, E.A. Leading the Way: Canine Models of Genomics and Disease. Dis. Models Mech. 2010, 3, 27–34. [Google Scholar] [CrossRef]
- Felsburg, P.J. Overview of Immune System Development in the Dog: Comparison with Humans. Hum. Exp. Toxicol. 2002, 21, 487–492. [Google Scholar] [CrossRef]
- Pereira, M.; Valério-Bolas, A.; Saraiva-Marques, C.; Alexandre-Pires, G.; Pereira Da Fonseca, I.; Santos-Gomes, G. Development of Dog Immune System: From in Uterus to Elderly. Vet. Sci. 2019, 6, 83. [Google Scholar] [CrossRef]
- Cissé, O.H.; Ma, L.; Dekker, J.P.; Khil, P.P.; Youn, J.-H.; Brenchley, J.M.; Blair, R.; Pahar, B.; Chabé, M.; Van Rompay, K.K.A.; et al. Genomic Insights into the Host Specific Adaptation of the Pneumocystis Genus. Commun. Biol. 2021, 4, 305. [Google Scholar] [CrossRef]
- Sukura, A.; Saari, S.; Järvinen, A.-K.; Olsson, M.; Kärkkäinen, M.; Ilvesniemi, T. Pneumocystis Carinii Pneumonia in Dogs—A Diagnostic Challenge. J. Vet. Diagn. Investig. 1996, 8, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Friedenberg, S.G.; Meurs, K.M. Genotype Imputation in the Domestic Dog. Mamm. Genome 2016, 27, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.N.; Friedenberg, S.G. Whole Animal Genome Sequencing: User-Friendly, Rapid, Containerized Pipelines for Processing, Variant Discovery, and Annotation of Short-Read Whole Genome Sequencing Data. G3 Genes Genomes Genet. 2023, 13, jkad117. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J.; Zody, M.C.; et al. Genome Sequence, Comparative Analysis and Haplotype Structure of the Domestic Dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows–Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Giger, U.; Reagan, K. Hereditary and Acquired Immunodeficiencies. In Greene’s Infectious Diseases of the Dog and Cat; Elsevier/Saunders: St. Louis, MO, USA, 2021; pp. 1728–1745. [Google Scholar]
- Pejaver, V.; Urresti, J.; Lugo-Martinez, J.; Pagel, K.A.; Lin, G.N.; Nam, H.-J.; Mort, M.; Cooper, D.N.; Sebat, J.; Iakoucheva, L.M.; et al. Inferring the Molecular and Phenotypic Impact of Amino Acid Variants with MutPred2. Nat. Commun. 2020, 11, 5918. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A Method and Server for Predicting Damaging Missense Mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed]
- Köhler, S.; Gargano, M.; Matentzoglu, N.; Carmody, L.C.; Lewis-Smith, D.; Vasilevsky, N.A.; Danis, D.; Balagura, G.; Baynam, G.; Brower, A.M.; et al. The Human Phenotype Ontology in 2021. Nucleic Acids Res. 2021, 49, D1207–D1217. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.E.; McLaren, W.; Gil, L.; Thormann, A.; Schuilenburg, H.; Sheppard, D.; Parton, A.; Armean, I.M.; Trevanion, S.J.; Flicek, P.; et al. Ensembl Variation Resources. Database 2018, 2018, bay119. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.J.; Amode, M.R.; Aneja, A.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Becker, A.; Bennett, R.; Berry, A.; Bhai, J.; et al. Ensembl 2023. Nucleic Acids Res. 2023, 51, D933–D941. [Google Scholar] [CrossRef] [PubMed]
- Weissenbacher-Lang, C.; Fuchs-Baumgartinger, A.; Klang, A.; Kneissl, S.; Pirker, A.; Shibly, S.; von Ritgen, S.; Weissenböck, H.; Künzel, F. Pneumocystis Carinii Infection with Severe Pneumomediastinum and Lymph Node Involvement in a Whippet Mixed-Breed Dog. J. Vet. Diagn. Investig. 2017, 29, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Merrill, K.; Coffey, E.; Furrow, E.; Masseau, I.; Rindt, H.; Reinero, C. X-Linked CD40 Ligand Deficiency in a 1-Year-Old Male Shih Tzu with Secondary Pneumocystis Pneumonia. Vet. Intern. Med. 2021, 35, 497–503. [Google Scholar] [CrossRef]
- Shamriz, O.; Simon, A.J.; Lev, A.; Megged, O.; Ledder, O.; Picard, E.; Joseph, L.; Molho-Pessach, V.; Tal, Y.; Millman, P.; et al. Exogenous Interleukin-2 Can Rescue in-Vitro T Cell Activation and Proliferation in Patients with a Novel Capping Protein Regulator and Myosin 1 Linker 2 Mutation. Clin. Exp. Immunol. 2020, 200, 215–227. [Google Scholar] [CrossRef]
- Yonkof, J.R.; Gupta, A.; Rueda, C.M.; Mangray, S.; Prince, B.T.; Rangarajan, H.G.; Alshahrani, M.; Varga, E.; Cripe, T.P.; Abraham, R.S. A Novel Pathogenic Variant in CARMIL2 (RLTPR) Causing CARMIL2 Deficiency and EBV-Associated Smooth Muscle Tumors. Front. Immunol. 2020, 11, 884. [Google Scholar] [CrossRef]
- Kolukisa, B.; Baser, D.; Akcam, B.; Danielson, J.; Bilgic Eltan, S.; Haliloglu, Y.; Sefer, A.P.; Babayeva, R.; Akgun, G.; Charbonnier, L.; et al. Evolution and Long-term Outcomes of Combined Immunodeficiency Due to CARMIL2 Deficiency. Allergy 2022, 77, 1004–1019. [Google Scholar] [CrossRef]
- Lévy, R.; Gothe, F.; Momenilandi, M.; Magg, T.; Materna, M.; Peters, P.; Raedler, J.; Philippot, Q.; Rack-Hoch, A.L.; Langlais, D.; et al. Human CARMIL2 Deficiency Underlies a Broader Immunological and Clinical Phenotype than CD28 Deficiency. J. Exp. Med. 2023, 220, e20220275. [Google Scholar] [CrossRef]
- Schober, T.; Magg, T.; Laschinger, M.; Rohlfs, M.; Linhares, N.D.; Puchalka, J.; Weisser, T.; Fehlner, K.; Mautner, J.; Walz, C.; et al. A Human Immunodeficiency Syndrome Caused by Mutations in CARMIL2. Nat. Commun. 2017, 8, 14209. [Google Scholar] [CrossRef]
- Stark, B.C.; Lanier, M.H.; Cooper, J.A. CARMIL Family Proteins as Multidomain Regulators of Actin-Based Motility. Mol. Biol. Cell 2017, 28, 1713–1723. [Google Scholar] [CrossRef]
- Roncagalli, R.; Cucchetti, M.; Jarmuzynski, N.; Grégoire, C.; Bergot, E.; Audebert, S.; Baudelet, E.; Menoita, M.G.; Joachim, A.; Durand, S.; et al. The Scaffolding Function of the RLTPR Protein Explains Its Essential Role for CD28 Co-Stimulation in Mouse and Human T Cells. J. Exp. Med. 2016, 213, 2437–2457. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Cucchetti, M.; Roncagalli, R.; Yokosuka, T.; Malzac, A.; Bertosio, E.; Imbert, J.; Nijman, I.J.; Suchanek, M.; Saito, T.; et al. The Lymphoid Lineage–Specific Actin-Uncapping Protein Rltpr Is Essential for Costimulation via CD28 and the Development of Regulatory T Cells. Nat. Immunol. 2013, 14, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Ménard, S.; Marques, C.; Berry, A.; Iriart, X. Immune Response in Pneumocystis Infections According to the Host Immune System Status. JoF 2021, 7, 625. [Google Scholar] [CrossRef] [PubMed]
- Alazami, A.M.; Al-Helale, M.; Alhissi, S.; Al-Saud, B.; Alajlan, H.; Monies, D.; Shah, Z.; Abouelhoda, M.; Arnaout, R.; Al-Dhekri, H.; et al. Novel CARMIL2 Mutations in Patients with Variable Clinical Dermatitis, Infections, and Combined Immunodeficiency. Front. Immunol. 2018, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Opata, M.M.; Hollifield, M.L.; Lund, F.E.; Randall, T.D.; Dunn, R.; Garvy, B.A.; Feola, D.J. B Lymphocytes Are Required during the Early Priming of CD4+ T Cells for Clearance of Pneumocystis Infection in Mice. J. Immunol. 2015, 195, 611–620. [Google Scholar] [CrossRef]
- Kolls, J.K. An Emerging Role of B Cell Immunity in Susceptibility to Pneumocystis Pneumonia. Am. J. Respir. Cell Mol. Biol. 2017, 56, 279–280. [Google Scholar] [CrossRef]
- Di Tosto, M.; Callegari, C.; Matiasek, K.; Lacava, G.; Salvatore, G.; Muñoz Declara, S.; Betti, B.; Tirrito, F. Case Report: Atypical and Chronic Masticatory Muscle Myositis in a 5-Month Old Cavalier King Charles Spaniel. Clinical and Diagnostic Findings, Treatment and Successful Outcome. Front. Vet. Sci. 2022, 9, 955758. [Google Scholar] [CrossRef]
- Pitcher, G.D.C.; Hahn, C.N. Atypical Masticatory Muscle Myositis in Three Cavalier King Charles Spaniel Littermates. J. Small Anim. Pract. 2007, 48, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Henthorn, P.; Somberg, R.; Fimiani, V.; Puck, J.; Patterson, D.; Felsburg, P. IL-2Rγ Gene Microdeletion Demonstrates That Canine X-Linked Severe Combined Immunodeficiency Is a Homologue of the Human Disease. Genomics 1994, 23, 69–74. [Google Scholar] [CrossRef]
- Somberg, R.L.; Pullen, R.P.; Casal, M.L.; Patterson, D.F.; Felsburg, P.J.; Henthorn, P.S. A Single Nucleotide Insertion in the Canine Interleukin-2 Receptor Gamma Chain Results in X-Linked Severe Combined Immunodeficiency Disease. Vet. Immunol. Immunopathol. 1995, 47, 203–213. [Google Scholar] [CrossRef]
- Donner, J.; Freyer, J.; Davison, S.; Anderson, H.; Blades, M.; Honkanen, L.; Inman, L.; Brookhart-Knox, C.A.; Louviere, A.; Forman, O.P.; et al. Genetic Prevalence and Clinical Relevance of Canine Mendelian Disease Variants in over One Million Dogs. PLoS Genet. 2023, 19, e1010651. [Google Scholar] [CrossRef]
- Letko, A.; Minor, K.M.; Jagannathan, V.; Seefried, F.R.; Mickelson, J.R.; Oliehoek, P.; Drögemüller, C. Genomic Diversity and Population Structure of the Leonberger Dog Breed. Genet. Sel. Evol. 2020, 52, 61. [Google Scholar] [CrossRef] [PubMed]
- Karousis, E.D.; Mühlemann, O. Nonsense-Mediated mRNA Decay Begins Where Translation Ends. Cold Spring Harb. Perspect. Biol. 2019, 11, a032862. [Google Scholar] [CrossRef] [PubMed]


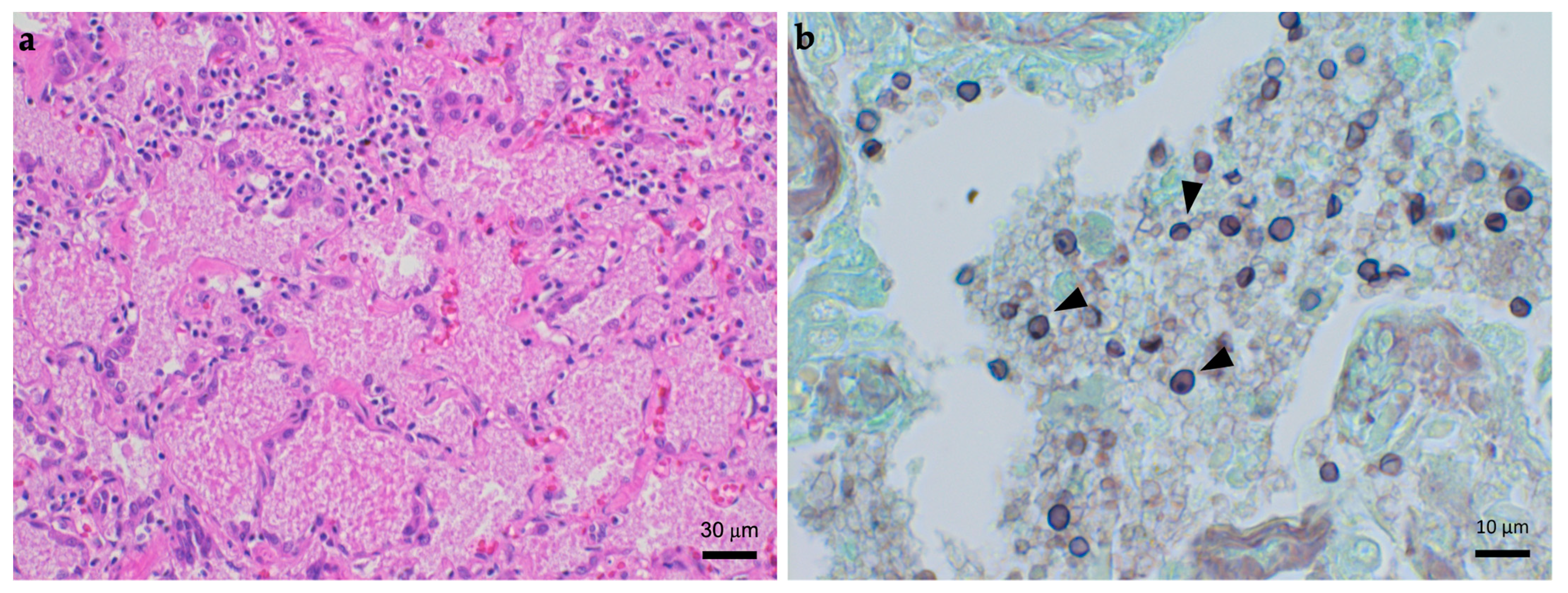{kind=link}
| Genomic Alteration (CanFam3.1) | Gene | UniProt ID | Protein Alteration | PolyPhen-2 HumVar † | MutPred2 † |
|---|---|---|---|---|---|
| CFA5 g.81801920G>A | CARMIL2 | A0A8I3NGS3 | p.R291* | NA | NA |
| CFA7 g.39301677C>T | LBR | A0A8C0SLC1 | p.P356L | 0.07 | 0.29 |
| CFA7 g.40051413G>A | NVL | A0A8C0M6L5 | p.R134Q | 0.00 | 0.05 |
| CFA14 g.32122502C>G | ENSCAFG00000002455 | A0A8I3N264 | p.R90Q | 0.00 | 0.31 |
| CFA23 g.9564400C>G | ENSCAFG00000048903 ‡ | NA ‡ | p.Q618H | 0.54 | 0.47 |
| CFA30 g.38173284A>C | C30H15orf39 | A0A8C0TAG0 | p.E61A | 0.12 | 0.27 |
| CFA30 g.38174844G>A | C30H15orf39 | A0A8C0TAG0 | p.A581T | 0.03 | 0.05 |
| CARMIL2 p.R291* Genotypes # | Total Number of Dogs | Variant Allele Frequency (var Alleles/Total Alleles) | |||
|---|---|---|---|---|---|
| var/var | var/ref | ref/ref | |||
| PI suspects * | |||||
| CKCS | 3 (2) | 0 | 9 (3) | 12 | 25% |
| Other/unknown breeds | 0 | 0 | 15 (15) | 15 | 0% |
| Breed/population controls | |||||
| CKCS | 0 | 13 | 123 | 136 | 4.8% |
| Other/unknown breed | 0 | 0 | 559 | 559 | 0% |
| Age and Sex | Diagnosis | Confirmation of Diagnosis | Other Conditions | Outcome | |
|---|---|---|---|---|---|
| D1 | 1.5 years, MI | PCP | Cytology and immunocytochemistry (BAL, lung FNA); PCR, immunohistochemistry, electron microscopy (lung tissue, post-mortem) | Skin abscess, early onset GI disease, erosive oral lesions | Died on the day of TMS therapy initiation |
| D2 | 16 months, MN | PCP | PCR (deep oropharyngeal swab) | Atopic dermatitis, chronic diarrhea, GI parasitism | Complete response to TMS and prednisone |
| D3 | 5 months, MI | Refractory Bordetella pneumonia | Aerobic culture (BAL fluid) and PCR (oropharyngeal swab) | MMM, hypocholesterolemia, chronic diarrhea, GI parasitism | Partial clinical response to inhaled gentamicin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coffey, E.L.; Ma, L.; Cissé, O.H.; Kovacs, J.A.; Minor, K.M.; Sukura, A.; Danesi, P.; Friedenberg, S.G.; Cullen, J.N.; Weissenbacher-Lang, C.; et al. A Novel CARMIL2 Immunodeficiency Identified in a Subset of Cavalier King Charles Spaniels with Pneumocystis and Bordetella Pneumonia. J. Fungi 2024, 10, 198. https://doi.org/10.3390/jof10030198
Coffey EL, Ma L, Cissé OH, Kovacs JA, Minor KM, Sukura A, Danesi P, Friedenberg SG, Cullen JN, Weissenbacher-Lang C, et al. A Novel CARMIL2 Immunodeficiency Identified in a Subset of Cavalier King Charles Spaniels with Pneumocystis and Bordetella Pneumonia. Journal of Fungi. 2024; 10(3):198. https://doi.org/10.3390/jof10030198
Chicago/Turabian StyleCoffey, Emily L., Liang Ma, Ousmane H. Cissé, Joseph A. Kovacs, Katie M. Minor, Antti Sukura, Patrizia Danesi, Steven G. Friedenberg, Jonah N. Cullen, Christiane Weissenbacher-Lang, and et al. 2024. "A Novel CARMIL2 Immunodeficiency Identified in a Subset of Cavalier King Charles Spaniels with Pneumocystis and Bordetella Pneumonia" Journal of Fungi 10, no. 3: 198. https://doi.org/10.3390/jof10030198
APA StyleCoffey, E. L., Ma, L., Cissé, O. H., Kovacs, J. A., Minor, K. M., Sukura, A., Danesi, P., Friedenberg, S. G., Cullen, J. N., Weissenbacher-Lang, C., Nadeau, J. C., Graham, A. M., Granick, M. N., Branson, N. K., Branson, K. C., Blasi, B., Jacobs, C. M., & Furrow, E. (2024). A Novel CARMIL2 Immunodeficiency Identified in a Subset of Cavalier King Charles Spaniels with Pneumocystis and Bordetella Pneumonia. Journal of Fungi, 10(3), 198. https://doi.org/10.3390/jof10030198