
Isolation of a Novel Pythium Species, P. thermoculicivorax, and Trichoderma sp. from Natural Enzootic Mosquito Larval Infections


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collecting Mosquito Larvae
2.2. Isolation of Pythium and Trichoderma Strains
2.3. Pythium Temperature Growth Tolerance Assays
2.4. DNA Extraction, PCR, and Sequencing
2.5. Generating Phylogenies
2.6. Microscopy
3. Results
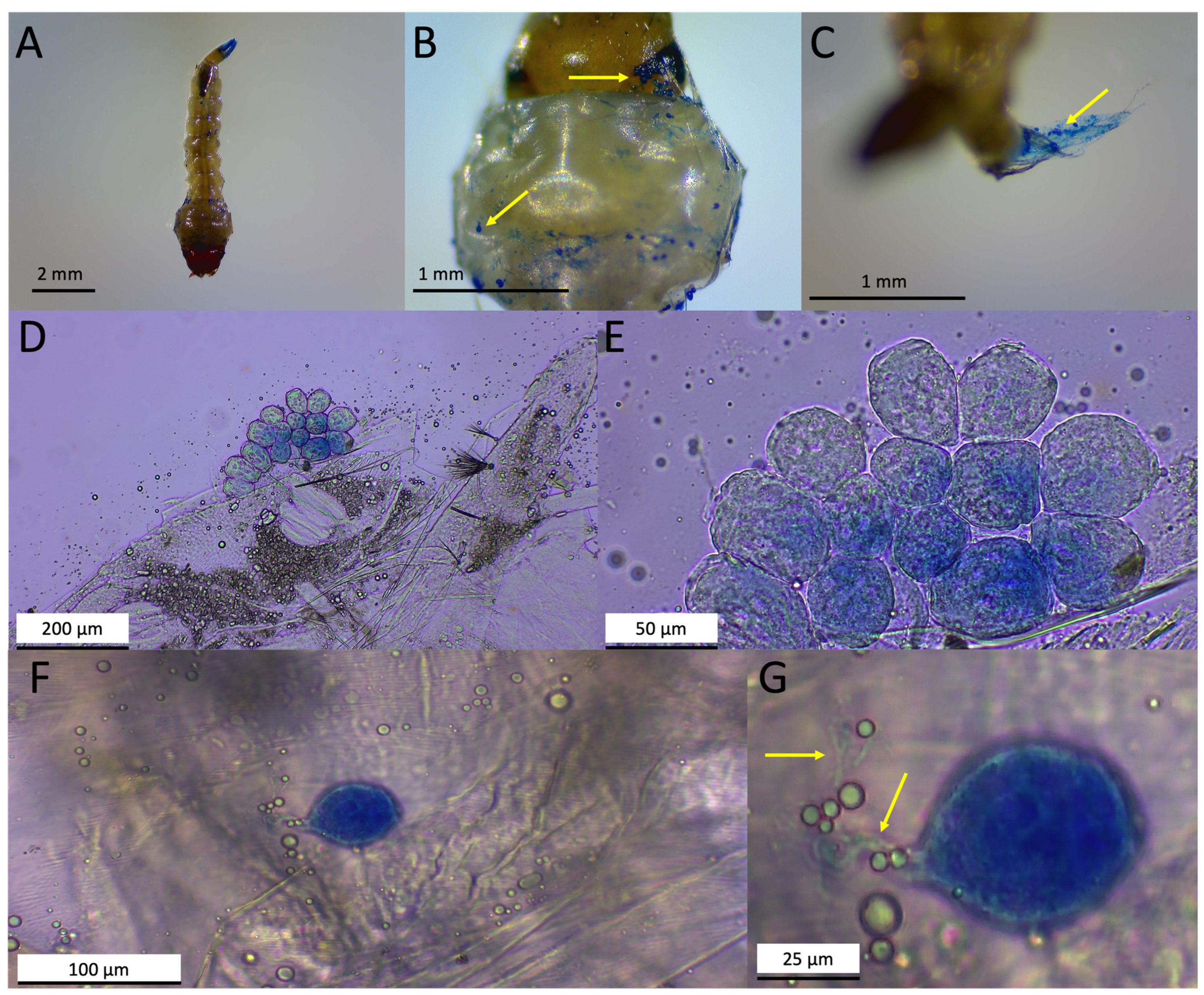
3.1. Sampling Sites and Macroscopic and Microscopic Characterizations of Infected Mosquitoes
3.2. Phenotypic and Molecular Characterizations of LCMP-P1—Pythium thermoculicivorax
3.3. Characterization of Isolates LCMP-T1, LCMP-T2, and LCMP-T3—Trichoderma sp.
3.4. P. thermoculcivorax Zoospores
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Kohl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef]
- Marrone, P.G. Status of the biopesticide market and prospects for new bioherbicides. Pest Manag. Sci. 2023, 80, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Koller, J.; Sutter, L.; Gonthier, J.; Collatz, J.; Norgrove, L. Entomopathogens and Parasitoids Allied in Biocontrol: A Systematic Review. Pathogens 2023, 12, 957. [Google Scholar] [CrossRef] [PubMed]
- Elya, C.; De Fine Licht, H.H. The genus Entomophthora: Bringing the insect destroyers into the twenty-first century. IMA Fungus 2021, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Kyle, C.H.; Liu, J.; Gallagher, M.E.; Dukic, V.; Dwyer, G. Stochasticity and Infectious Disease Dynamics: Density and Weather Effects on a Fungal Insect Pathogen. Am. Nat. 2020, 195, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Touray, M.; Bakirci, S.; Ulug, D.; Gulsen, S.H.; Cimen, H.; Yavasoglu, S.I.; Simsek, F.M.; Ertabaklar, H.; Ozbel, Y.; Hazir, S. Arthropod vectors of disease agents: Their role in public and veterinary health in Turkiye and their control measures. Acta Trop. 2023, 243, 106893. [Google Scholar] [CrossRef]
- Onen, H.; Luzala, M.M.; Kigozi, S.; Sikumbili, R.M.; Muanga, C.K.; Zola, E.N.; Wendji, S.N.; Buya, A.B.; Balciunaitiene, A.; Viskelis, J.; et al. Mosquito-Borne Diseases and Their Control Strategies: An Overview Focused on Green Synthesized Plant-Based Metallic Nanoparticles. Insects 2023, 14, 221. [Google Scholar] [CrossRef]
- Giunti, G.; Becker, N.; Benelli, G. Invasive mosquito vectors in Europe: From bioecology to surveillance and management. Acta Trop. 2023, 239, 106832. [Google Scholar] [CrossRef]
- Duval, P.; Antonelli, P.; Aschan-Leygonie, C.; Valiente Moro, C. Impact of Human Activities on Disease-Spreading Mosquitoes in Urban Areas. J. Urban Health 2023, 100, 591–611. [Google Scholar] [CrossRef] [PubMed]
- Arnal, A.; Roche, B.; Gouagna, L.C.; Dujon, A.; Ujvari, B.; Corbel, V.; Remoue, F.; Poinsignon, A.; Pompon, J.; Giraudeau, M.; et al. Cancer and mosquitoes—An unsuspected close connection. Sci. Total Environ. 2020, 743, 140631. [Google Scholar] [CrossRef]
- Vatandoost, H.; Hanafi-Bojd, A.A.; Nikpoor, F.; Raeisi, A.; Abai, M.R.; Zaim, M. Situation of insecticide resistance in malaria vectors in the World Health Organization of Eastern Mediterranean region 1990–2020. Toxicol. Res. 2022, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tepa, A.; Kengne-Ouafo, J.A.; Djova, V.S.; Tchouakui, M.; Mugenzi, L.M.J.; Djouaka, R.; Pieme, C.A.; Wondji, C.S. Molecular Drivers of Multiple and Elevated Resistance to Insecticides in a Population of the Malaria Vector Anopheles gambiae in Agriculture Hotspot of West Cameroon. Genes 2022, 13, 1206. [Google Scholar] [CrossRef] [PubMed]
- Sonhafouo-Chiana, N.; Nkahe, L.D.; Kopya, E.; Awono-Ambene, P.H.; Wanji, S.; Wondji, C.S.; Antonio-Nkondjio, C. Rapid evolution of insecticide resistance and patterns of pesticides usage in agriculture in the city of Yaounde, Cameroon. Parasites Vectors 2022, 15, 186. [Google Scholar] [CrossRef] [PubMed]
- Meier, C.J.; Rouhier, M.F.; Hillyer, J.F. Chemical Control of Mosquitoes and the Pesticide Treadmill: A Case for Photosensitive Insecticides as Larvicides. Insects 2022, 13, 1093. [Google Scholar] [CrossRef] [PubMed]
- Katak, R.M.; Cintra, A.M.; Burini, B.C.; Marinotti, O.; Souza-Neto, J.A.; Rocha, E.M. Biotechnological Potential of Microorganisms for Mosquito Population Control and Reduction in Vector Competence. Insects 2023, 14, 718. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.F.V.; N’Guessan, R.; Koenraadt, C.J.M.; Asidi, A.; Farenhorst, M.; Akogbeto, M.; Thomas, M.B.; Knols, B.G.J.; Takken, W. The entomopathogenic fungus Beauveria bassiana reduces instantaneous blood feeding in wild multi-insecticide-resistant Culex quinquefasciatus mosquitoes in Benin, West Africa. Parasites Vectors 2010, 3, 87. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.F.V.; Koenraadt, C.J.M.; Farenhorst, M.; Knols, B.G.J.; Takken, W. Pyrethroid resistance in Anopheles gambiae leads to increased susceptibility to the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana. Malar. J. 2010, 9, 168. [Google Scholar] [CrossRef] [PubMed]
- Scholte, E.J.; Knols, B.G.; Samson, R.A.; Takken, W. Entomopathogenic fungi for mosquito control: A review. J. Insect Sci. 2004, 4, 19. [Google Scholar] [CrossRef]
- Bilgo, E.; Lovett, B.; Bayili, K.; Millogo, A.S.; Sare, I.; Dabire, R.K.; Sanon, A.; St Leger, R.J.; Diabate, A. Transgenic Metarhizium pingshaense synergistically ameliorates pyrethroid-resistance in wild-caught, malaria-vector mosquitoes. PLoS ONE 2018, 13, e0203529. [Google Scholar] [CrossRef]
- Fang, W.G.; Vega-Rodriguez, J.; Ghosh, A.K.; Jacobs-Lorena, M.; Kang, A.; St Leger, R.J. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 2011, 331, 1074–1077. [Google Scholar] [CrossRef]
- Clark, T.B.; Kellen, W.R.; Lindegren, J.E.; Sanders, R.D. Pythium sp. (Phycomycetes:Pythiales) pathogenic to mosquito larvae. J. Invertebr. Pathol. 1966, 8, 351–354. [Google Scholar] [CrossRef]
- Seymour, R.L. Leptolegnia Chapmanii, an oomycete pathogen of mosquito larvae. Mycologia 1984, 76, 670–674. [Google Scholar] [CrossRef]
- Vilela, R.; Humber, R.A.; Taylor, J.W.; Mendoza, L. Phylogenetic and physiological traits of oomycetes originally identified as Lagenidium giganteum from fly and mosquito larvae. Mycologia 2019, 111, 408–422. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.C.; Roberts, D.W. Effects of salinity, pH, organic solutes, anaerobic conditions, and the presence of other microbes on production and survival of Lagenidium giganteum (Oomycetes: Lagenidiales) zoospores. J. Invertebr. Pathol. 1985, 45, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, J.L.; Washino, R.K. Field evaluation of Lagenidium giganteum (Oomycetes: Lagenidiales) and description of a natural epizootic involving a new isolate of the fungus. J. Med. Entomol. 1988, 25, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, J.L. Oomycetes: Lagenidium giganteum. J. Am. Mosq. Control Assoc. 2007, 23, 50–57. [Google Scholar] [CrossRef]
- Vilela, R.; Taylor, J.W.; Walker, E.D.; Mendoza, L. Lagenidium giganteum pathogenicity in mammals. Emerg. Infect. Dis. 2015, 21, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Spies, C.F.J.; Grooters, A.M.; Levesque, C.A.; Rintoul, T.L.; Redhead, S.A.; Glockling, S.L.; Chen, C.Y.; de Cock, A. Molecular phylogeny and taxonomy of Lagenidium-like oomycetes pathogenic to mammals. Fungal Biol. 2016, 120, 931–947. [Google Scholar] [CrossRef]
- Lord, J.C.; Fukuda, T. An ultrastructural study of the invasion of Culex quinquefasciatus larvae by Leptolegnia chapmanii (Oomycetes: Saprolegniales). Mycopathologia 1988, 104, 67–73. [Google Scholar] [CrossRef]
- Pelizza, S.A.; Scorsetti, A.C.; Lastra, C.C.; Garcia, J.J. Production of oogonia and oospores of Leptolegnia chapmanii Seymour (Straminipila: Peronosporomycetes) in Aedes aegypti (L.) larvae at different temperatures. Mycopathologia 2010, 169, 71–74. [Google Scholar] [CrossRef]
- Paramo, M.E.R.; Falvo, M.; Garcia, J.; Lastra, C.C.L. Compatibility between Leptolegnia chapmanii and diflubenzuron and neem oil for the control of Aedes aegypti. Rev. Argent. Microbiol. 2020, 52, 240–244. [Google Scholar] [CrossRef]
- Su, X.; Zou, F.; Guo, Q.; Huang, J.; Chen, T. A report on a mosquito-killing fungus, Pythium carolinianum. Fungal Divers. 2001, 7, 129–133. [Google Scholar]
- Huang, S.-W.; Su, X.-Q. Biological Studies on Pythium guiyangense, a fungal pathogen of mosquito larvae. Mycosystema 2007, 23, 380–388. [Google Scholar]
- Vilela, R.; Montalva, C.; Luz, C.; Humber, R.A.; Mendoza, L. Pythium insidiosum isolated from infected mosquito larvae in central Brazil. Acta Trop. 2018, 185, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Dong, Y.; Wei, Y.; Zhang, M.; Wang, J.; Tang, Z.; Xia, Q.; Nyawira, K.T.; Jing, M.; Dou, D.; et al. Genome-wide and functional analyses of tyrosine kinase-like family genes reveal potential roles in development and virulence in mosquito pathogen Pythium guiyangense. Fungal Genet. Biol. 2019, 130, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Tang, Z.; Wang, C.; Wang, J.; Dong, Y.; Chen, Y.; Wei, Y.; Cheng, B.; Zhang, M.; Grenville-Briggs, L.J.; et al. Infection mechanisms and putative effector repertoire of the mosquito pathogenic oomycete Pythium guiyangense uncovered by genomic analysis. PLoS Genet. 2019, 15, e1008116. [Google Scholar] [CrossRef] [PubMed]
- Darsie, R.F., Jr.; Ward, R.A. Identification and Geographic Distribution of the Mosquitoes of North America, North of Mexico; University of Florida Press: Gainesville, FL, USA, 2005. [Google Scholar]
- Dou, K.; Lu, Z.; Wu, Q.; Ni, M.; Yu, C.; Wang, M.; Li, Y.; Wang, X.; Xie, H.; Chen, J.; et al. MIST: A Multilocus Identification System for Trichoderma. Appl. Environ. Microbiol. 2020, 86, e01532-20. [Google Scholar] [CrossRef] [PubMed]
- Robideau, G.P.; De Cock, A.W.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Desaulniers, N.; Eggertson, Q.A.; Gachon, C.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.; Zhu, H.; Guan, Y.; Lam, T.T.Y. ggtree: An R package for visualization and annotation of phylogenetic trees with different types of meta-data. Methods Ecol. Evol. 2024, 8, 28–36. [Google Scholar] [CrossRef]
- Yadav, M.; Dahiya, N.; Sehrawat, N. Mosquito gene targeted RNAi studies for vector control. Funct. Integr. Genom. 2023, 23, 180. [Google Scholar] [CrossRef]
- dos Santos, D.R.; Chaves, L.L.; Pires, V.C.; Rodrigues, J.S.; de Assunção, M.A.S.; Faierstein, G.B.; Neto, A.G.B.; de Souza Rebouças, J.; de Magalhães Cabral Albuquerque, E.C.; de Melo, S.A.B.V.; et al. New weapons against the disease vector Aedes aegypti: From natural products to nanoparticles. Int. J. Pharm. 2023, 643, 123221. [Google Scholar] [CrossRef]
- Piombo, E.; Kelbessa, B.G.; Sundararajan, P.; Whisson, S.C.; Vetukuri, R.R.; Dubey, M. RNA silencing proteins and small RNAs in oomycete plant pathogens and biocontrol agents. Front. Microbiol. 2023, 14, 1076522. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.E.I.; Paula, A.R.; Ribeiro, A.; Butt, T.M.; Silva, C.P.; Samuels, R.I. A new method of deploying entomopathogenic fungi to control adult Aedes aegypti mosquitoes. J. Appl. Entomol. 2018, 142, 59–66. [Google Scholar] [CrossRef]
- Bukhari, T.; Takken, W.; Koenraadt, C.J.M. Development of Metarhizium anisopliae and Beauveria bassiana formulations for control of malaria mosquito larvae. Parasites Vector 2011, 4, 23. [Google Scholar] [CrossRef]
- Qin, Y.; Liu, X.; Peng, G.; Xia, Y.; Cao, Y. Recent Advancements in Pathogenic Mechanisms, Applications and Strategies for Entomopathogenic Fungi in Mosquito Biocontrol. J. Fungi 2023, 9, 746. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Bergamini, C.; Montalva, C.; Humber, R.A.; Luz, C. Simple method to detect and to isolate entomopathogenic fungi (Hypocreales) from mosquito larvae. J. Invertebr. Pathol. 2021, 182, 107581. [Google Scholar] [CrossRef]
- Leles, R.N.; Lopez Lastra, C.C.; Garcia, J.J.; Fernandes, E.K.; Luz, C. A simple method for the detection of Leptolegnia chapmanii from infected Aedes aegypti larvae. Can. J. Microbiol. 2013, 59, 425–429. [Google Scholar] [CrossRef]
- Accoti, A.; Engdahl, C.S.; Dimopoulos, G. Discovery of Novel Entomopathogenic Fungi for Mosquito-Borne Disease Control. Front. Fungal Biol. 2021, 2, 637234. [Google Scholar] [CrossRef] [PubMed]
- Kikankie, C.K.; Brooke, B.D.; Knols, B.G.J.; Koekemoer, L.L.; Farenhorst, M.; Hunt, R.H.; Thomas, M.B.; Coetzee, M. The infectivity of the entomopathogenic fungus Beauveria bassiana to insecticide-resistant and susceptible Anopheles arabiensis mosquitoes at two different temperatures. Malar. J. 2010, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Farenhorst, M.; Knols, B.G.J.; Thomas, M.B.; Howard, A.F.V.; Takken, W.; Rowland, M.; N’Guessan, R. Synergy in efficacy of fungal entomopathogens and permethrin against West African insecticide-resistant Anopheles gambiae mosquitoes. PLoS ONE 2010, 5, e12081. [Google Scholar] [CrossRef] [PubMed]
- Farenhorst, M.; Mouatcho, J.C.; Kikankie, C.K.; Brooke, B.D.; Hunt, R.H.; Thomas, M.B.; Koekemoer, L.L.; Knols, B.G.J.; Coetzee, M. Fungal infection counters insecticide resistance in African malaria mosquitoes. Proc. Natl. Acad. Sci. USA 2009, 106, 17443–17447. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.N.; Wang, Z.L.; Huang, Y.; Keyhani, N.O.; Huang, Z. Lack of resistance development in Bemisia tabaci to Isaria fumosorosea after multiple generations of selection. Sci. Rep. 2017, 7, 42727. [Google Scholar] [CrossRef] [PubMed]
- Bickel, J.T.; Koehler, A.M. Review of Pythium species causing damping-off in corn. Plant Health Prog. 2021, 22, 219–225. [Google Scholar] [CrossRef]
- Paul, B.; Bouchet, P.; Bouchet, F. Some species of Pythium isolated from soil and water samples collected in western Africa. Microbiol. Res. 1994, 149, 217–222. [Google Scholar] [CrossRef]
- Broders, K.D.; Wallhead, M.W.; Austin, G.D.; Lipps, P.E.; Paul, P.A.; Mullen, R.W.; Dorrance, A.E. Association of soil chemical and physical properties with Pythium species diversity, community composition, and disease incidence. Phytopathology 2009, 99, 957–967. [Google Scholar] [CrossRef]
- Nam, B.; Choi, Y.J. Phytopythium and Pythium Species (Oomycota) Isolated from Freshwater Environments of Korea. Mycobiology 2019, 47, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sundaresha, S.; Bhardwaj, V. Biotechnological approaches in management of oomycetes diseases. 3 Biotech 2021, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- de Andrade Lourenco, D.; Branco, I.; Choupina, A. Phytopathogenic oomycetes: A review focusing on Phytophthora cinnamomi and biotechnological approaches. Mol. Biol. Rep. 2020, 47, 9179–9188. [Google Scholar] [CrossRef]
- Bozkurt, T.O.; Schornack, S.; Banfield, M.J.; Kamoun, S. Oomycetes, effectors, and all that jazz. Curr. Opin. Plant Biol. 2012, 15, 483–492. [Google Scholar] [CrossRef]
- Jiang, R.H.; Tyler, B.M. Mechanisms and evolution of virulence in oomycetes. Annu. Rev. Phytopathol. 2012, 50, 295–318. [Google Scholar] [CrossRef]
- Phillips, A.J.; Anderson, V.L.; Robertson, E.J.; Secombes, C.J.; van West, P. New insights into animal pathogenic oomycetes. Trends Microbiol. 2008, 16, 13–19. [Google Scholar] [CrossRef]
- Gaastra, W.; Lipman, L.J.; De Cock, A.W.; Exel, T.K.; Pegge, R.B.; Scheurwater, J.; Vilela, R.; Mendoza, L. Pythium insidiosum: An overview. Vet. Microbiol. 2010, 146, 1–16. [Google Scholar] [CrossRef]
- Sur, B.; Bihari, V.; Sharma, A.; Joshi, A.K. Studies on physiology, zoospore morphology and entomopathogenic potential of the aquatic oomycete: Lagenidium giganteum. Mycopathologia 2002, 154, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Jaronski, S.; Axtell, R.C. Simplified production system for fungus Lagenidium giganteum for operational mosquito control. Mosq. News 1984, 44, 377–381. [Google Scholar]
- Chen, C.-M.; Hsieh, H.-J.; Hu, B.-Y.; Fu, C.-H. Mosquito-killing water molds isolated from soil samples collected in Taiwan. Pedobiologia 2005, 49, 585–589. [Google Scholar] [CrossRef]
- Shen, D.; Wang, J.; Dong, Y.; Zhang, M.; Tang, Z.; Xia, Q.; Nyawira, K.T.; Jing, M.; Dou, D.; Xia, A. The glycoside hydrolase 18 family chitinases are associated with development and virulence in the mosquito pathogen Pythium guiyangense. Fungal Genet. Biol. 2020, 135, 103290. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, D.; Wang, J.; Chen, Y.; Dong, Y.; Tang, Z.; Xia, A. An AGC kinase, PgAGC1 regulates virulence in the entomopathogenic oomycete Pythium guiyangense. Fungal Biol. 2019, 123, 87–93. [Google Scholar] [CrossRef]
- Sparrow, F.K. Two new species of Pythium parasitic in green algae. Ann. Bot. 1931, 45, 257–277. [Google Scholar] [CrossRef]
- Middleton, J.T. The taxonomy, host range, and geographic distribution of the genus Pythium. Mem. Torrey Bot. Club 1943, 20, 1–171. [Google Scholar]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Chaverri, P.; Castlebury, L.A.; Overton, B.E.; Samuels, G.J. Hypocrea/Trichoderma: Species with conidiophore elongations and green conidia. Mycologia 2003, 95, 1100–1140. [Google Scholar] [CrossRef] [PubMed]
- Degenkolb, T.; Grafenhan, T.; Berg, A.; Nirenberg, H.I.; Gams, W.; Bruckner, H. Peptaibiomics: Screening for polypeptide antibiotics (peptaibiotics) from plant-protective Trichoderma species. Chem. Biodivers. 2006, 3, 593–610. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Quintero, C.A.; Atanasova, L.; Franco-Molano, A.E.; Gams, W.; Komon-Zelazowska, M.; Theelen, B.; Muller, W.H.; Boekhout, T.; Druzhinina, I. DNA barcoding survey of Trichoderma diversity in soil and litter of the Colombian lowland Amazonian rainforest reveals Trichoderma strigosellum sp. nov. and other species. Antonie Van Leeuwenhoek 2013, 104, 657–674. [Google Scholar] [CrossRef]
- Amanullah, F.; Khan, W.U. Trichoderma asperellum L. Coupled the Effects of Biochar to Enhance the Growth and Physiology of Contrasting Maize Cultivars under Copper and Nickel Stresses. Plants 2023, 12, 958. [Google Scholar] [CrossRef]
- Chou, H.; Xiao, Y.T.; Tsai, J.N.; Li, T.T.; Wu, H.Y.; Liu, L.D.; Tzeng, D.S.; Chung, C.L. In Vitro and in Planta Evaluation of Trichoderma asperellum TA as a Biocontrol Agent against Phellinus noxius, the Cause of Brown Root Rot Disease of Trees. Plant Dis. 2019, 103, 2733–2741. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; Marrez, D.A. Antagonistic activity of Trichoderma asperellum against Fusarium species, chemical profile and their efficacy for management of Fusarium-root rot disease in dry bean. Pest Manag. Sci. 2023, 80, 1153–1167. [Google Scholar] [CrossRef] [PubMed]
- Ramangouda, G.; Naik, M.K.; Nitnavare, R.B.; Yeshvekar, R.; Bhattacharya, J.; Bhatnagar-Mathur, P.; Sharma, M. Genetic enhancement of Trichoderma asperellum biocontrol potentials and carbendazim tolerance for chickpea dry root rot disease management. PLoS ONE 2023, 18, e0280064. [Google Scholar] [CrossRef]
- Herrera-Tellez, V.I.; Cruz-Olmedo, A.K.; Plasencia, J.; Gavilanes-Ruiz, M.; Arce-Cervantes, O.; Hernandez-Leon, S.; Saucedo-Garcia, M. The Protective Effect of Trichoderma asperellum on Tomato Plants against Fusarium oxysporum and Botrytis cinerea Diseases Involves Inhibition of Reactive Oxygen Species Production. Int. J. Mol. Sci. 2019, 20, 2007. [Google Scholar] [CrossRef] [PubMed]
- Batool, R.; Umer, M.J.; Wang, Y.; He, K.; Zhang, T.; Bai, S.; Zhi, Y.; Chen, J.; Wang, Z. Synergistic Effect of Beauveria bassiana and Trichoderma asperellum to Induce Maize (Zea mays L.) Defense against the Asian Corn Borer, Ostrinia furnacalis (Lepidoptera, Crambidae) and Larval Immune Response. Int. J. Mol. Sci. 2020, 21, 8215. [Google Scholar] [CrossRef] [PubMed]
- Saharan, R.; Patil, J.A.; Yadav, S.; Kumar, A.; Goyal, V. The nematicidal potential of novel fungus, Trichoderma asperellum FbMi6 against Meloidogyne incognita. Sci. Rep. 2023, 13, 6603. [Google Scholar] [CrossRef] [PubMed]
- Podder, D.; Ghosh, S.K. A new application of Trichoderma asperellum as an anopheline larvicide for eco friendly management in medical science. Sci. Rep. 2019, 9, 1108. [Google Scholar] [CrossRef]
- da Silveira, A.A.; Andrade, J.S.P.; Guissoni, A.C.P.; da Costa, A.C.; de Carvalho, E.S.A.; da Silva, H.G.; Brito, P.; de Souza, G.R.L.; Fernandes, K.F. Larvicidal potential of cell wall degrading enzymes from Trichoderma asperellum against Aedes aegypti (Diptera: Culicidae). Biotechnol. Prog. 2021, 37, e3182. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, R.; Darrisaw, C.; Lloyd, A.; Hoel, D.; Keyhani, N.O. Isolation of a Novel Pythium Species, P. thermoculicivorax, and Trichoderma sp. from Natural Enzootic Mosquito Larval Infections. J. Fungi 2024, 10, 199. https://doi.org/10.3390/jof10030199
Joseph R, Darrisaw C, Lloyd A, Hoel D, Keyhani NO. Isolation of a Novel Pythium Species, P. thermoculicivorax, and Trichoderma sp. from Natural Enzootic Mosquito Larval Infections. Journal of Fungi. 2024; 10(3):199. https://doi.org/10.3390/jof10030199
Chicago/Turabian StyleJoseph, Ross, Constance Darrisaw, Aaron Lloyd, David Hoel, and Nemat O. Keyhani. 2024. "Isolation of a Novel Pythium Species, P. thermoculicivorax, and Trichoderma sp. from Natural Enzootic Mosquito Larval Infections" Journal of Fungi 10, no. 3: 199. https://doi.org/10.3390/jof10030199
APA StyleJoseph, R., Darrisaw, C., Lloyd, A., Hoel, D., & Keyhani, N. O. (2024). Isolation of a Novel Pythium Species, P. thermoculicivorax, and Trichoderma sp. from Natural Enzootic Mosquito Larval Infections. Journal of Fungi, 10(3), 199. https://doi.org/10.3390/jof10030199