The Application of Trichoderma Strains or Metabolites Alters the Olive Leaf Metabolome and the Expression of Defense-Related Genes

 ,
, 
 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains, Bioactive Metabolites, and Culture Conditions
2.2. Plant Material and Experimental Design
2.3. Relative Quantification of Gene Expression
2.4. Extraction of Phenolic Compound from Olive Leaves
2.5. Determination of Phenolic Compounds by LC–MS Q-TOF
2.6. Data Analysis and Compounds Identification
3. Results
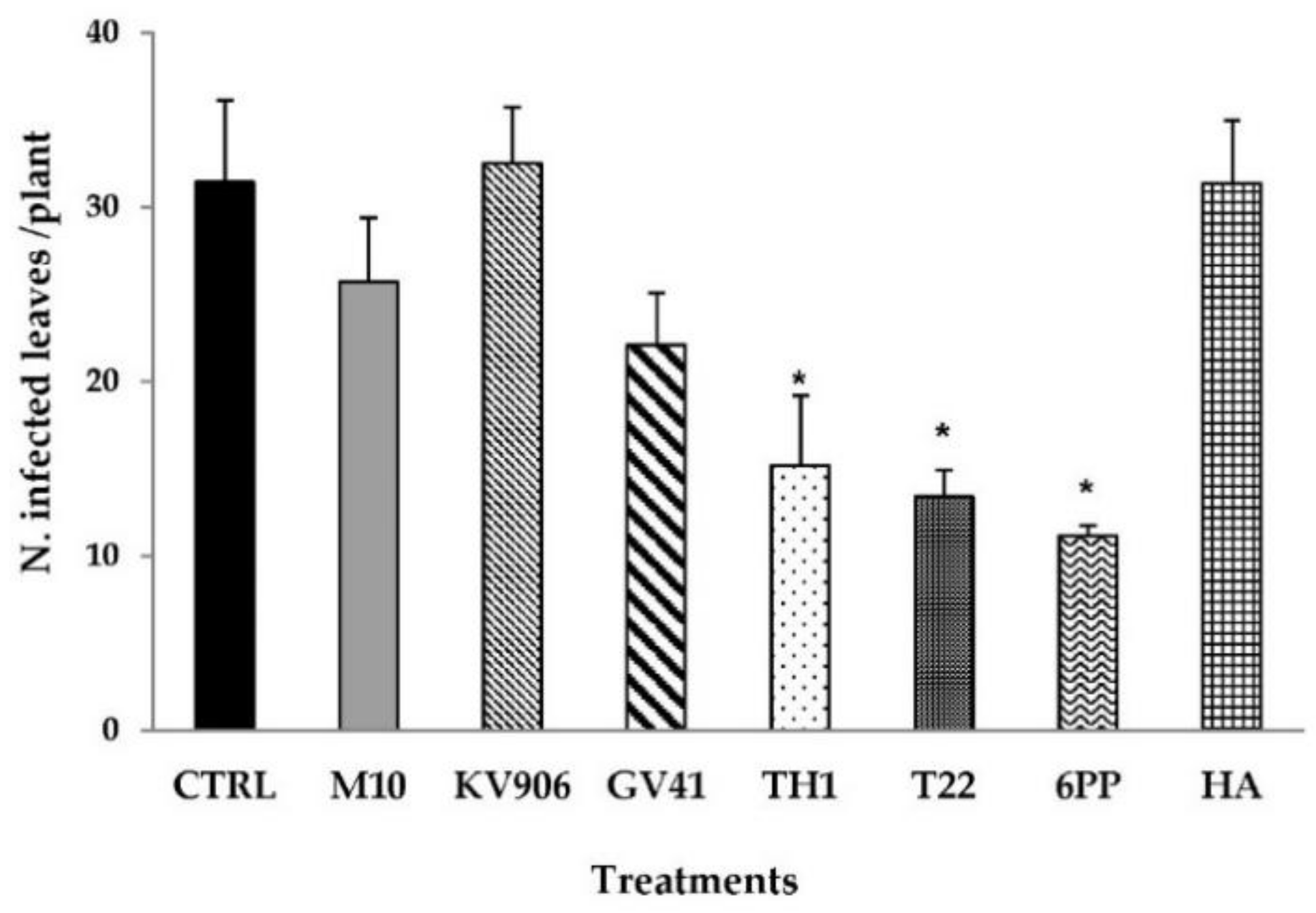
3.1. Effects on Disease Symptoms and Plant Growth
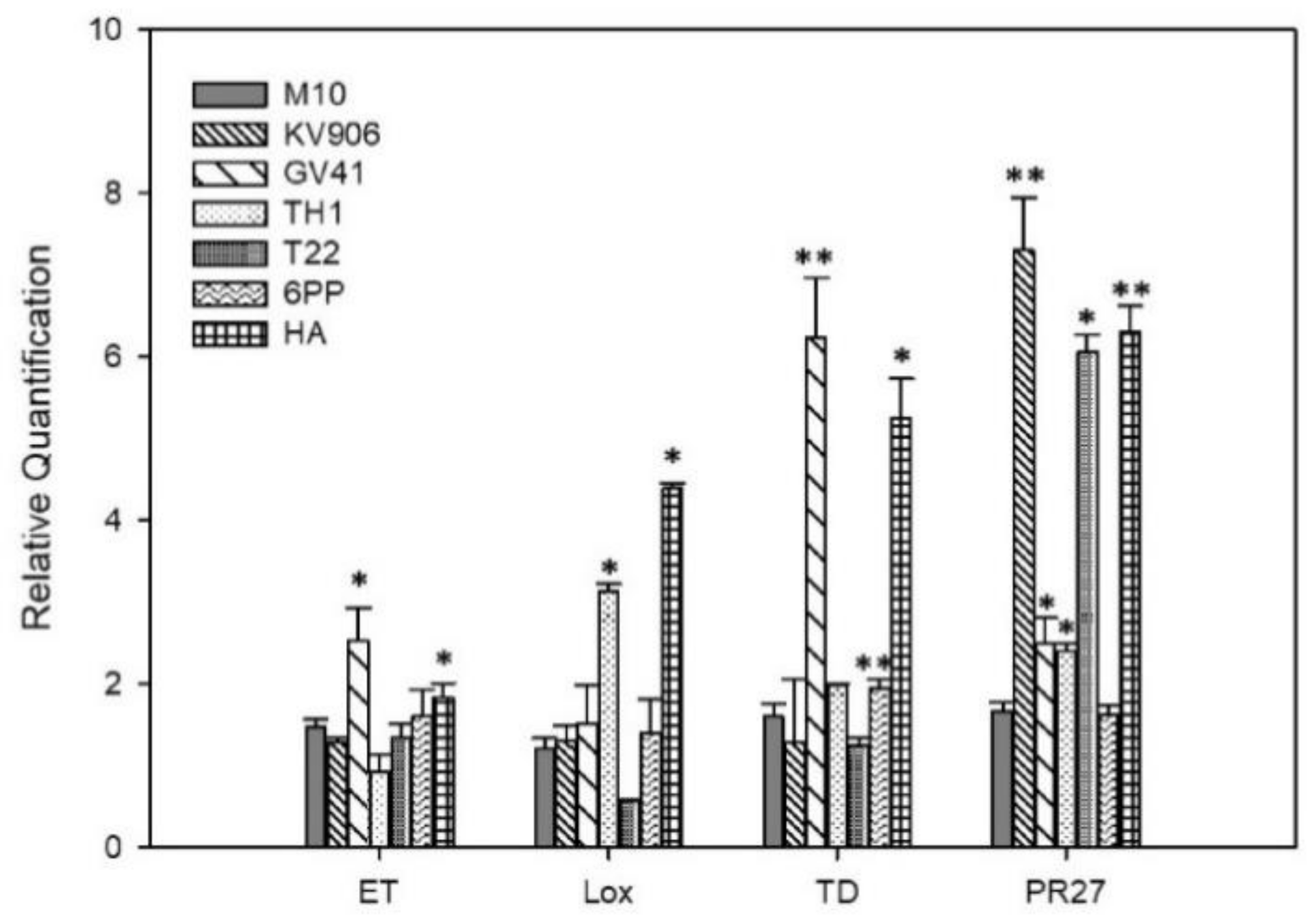
3.2. Induction of Plant Defense Responses
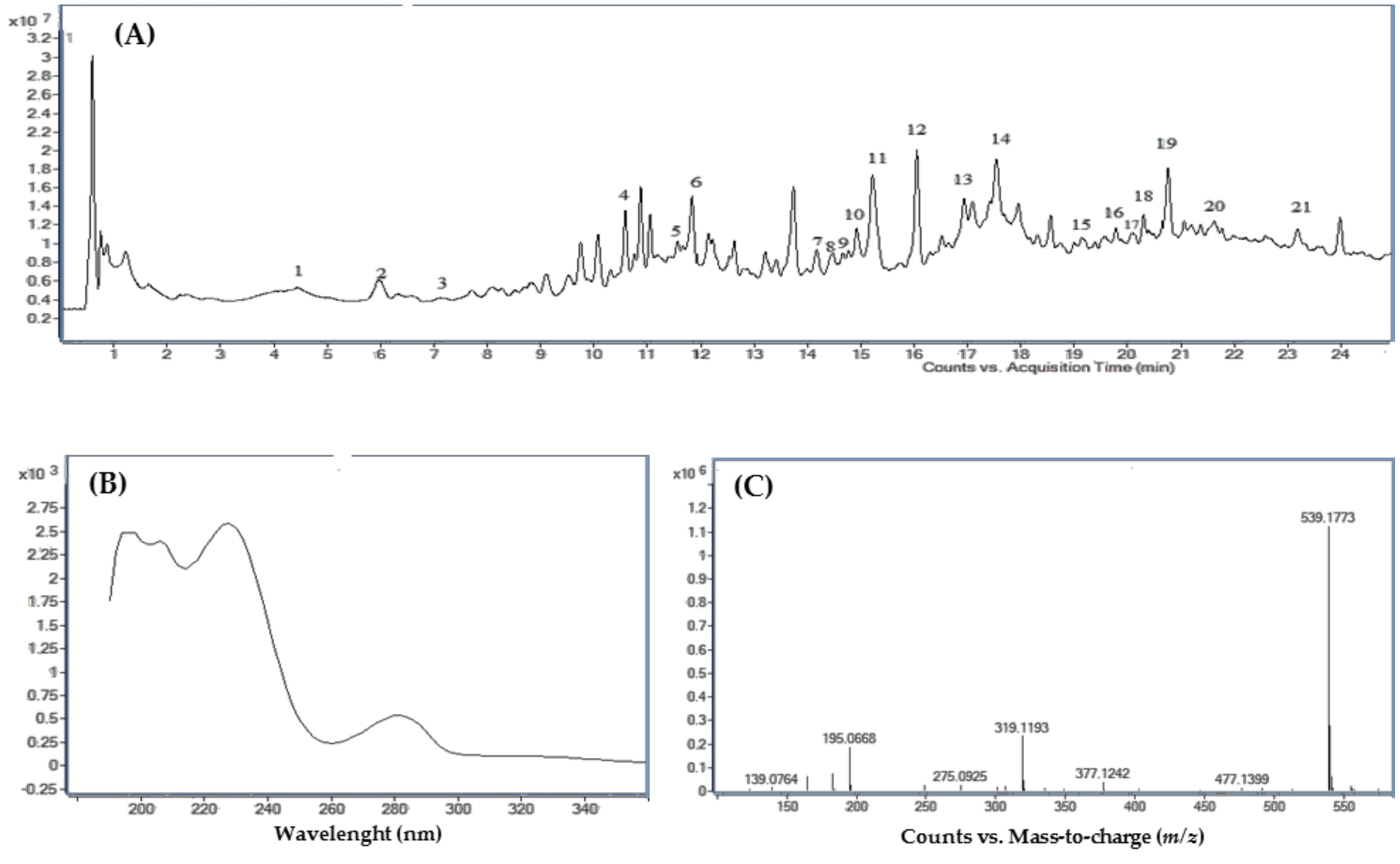
3.3. Characterization of Olive Leaf Metabolome
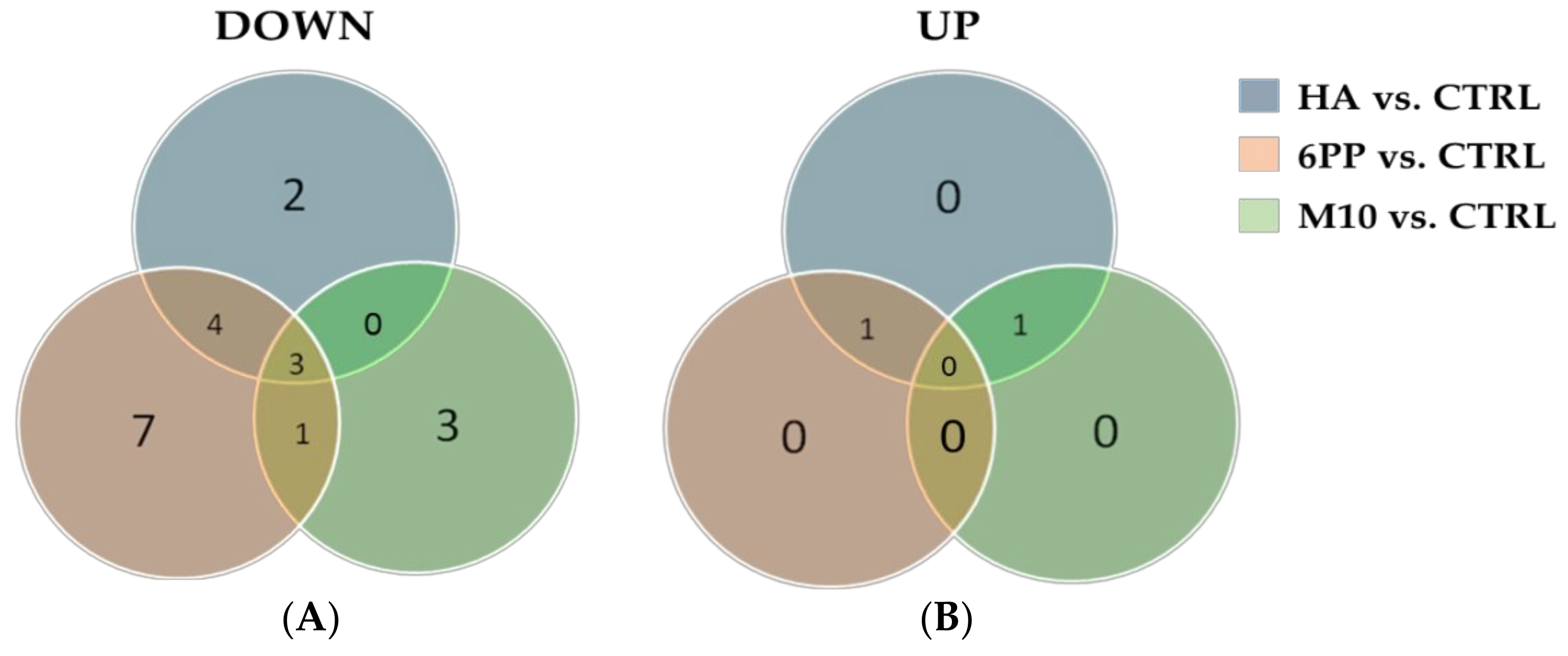
3.4. Untargeted Metabolomic Analysis of Leaf Extracts
3.5. Targeted Metabolomic Analysis of Leaf Extracts
3.6. Effect of Treatments on the Expression of Genes Involved in Oleuropein Biosynthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vossen, P. Olive Oil: History, Production, and Characteristics of the World’s Classic Oils. HortScience 2007, 42, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.-F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. Camb. Philos. Soc. 2012, 87, 885–899. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.P.; Ferreira, I.C.; Marcelino, F.; Valentão, P.; Andrade, P.B.; Seabra, R.; Estevinho, L.; Bento, A.; Pereira, J.A. Phenolic compounds and antimicrobial activity of olive (Olea europaea L. cv. Cobrançosa) leaves. Molecules 2007, 12, 1153–1162. [Google Scholar] [CrossRef]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Riccioloni, C.; Arcioni, S.; Vendramin, G.G.; Buonamici, A.; Porceddu, A.; Sarri, V.; Ojeda, M.A.; et al. A consensus list of microsatellites markers for olive genotyping. Mol. Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Keys, A. Mediterranean diet and public health: Personal reflections. Am. J. Clin. Nutr. 1995, 61, 1321–1323. [Google Scholar] [CrossRef] [Green Version]
- Romero, C.; Medina, E.; Mateo, M.A.; Brenes, M. New by-products rich in bioactive substances from the olive oil mill processing. J. Sci. Food Agric. 2018, 98, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Sahin, S.; Bilgin, M. Olive tree (Olea europaea L.) leaf as a waste by-product of table olive and olive oil industry: A review. J. Sci. Food Agric. 2018, 98, 1271–1279. [Google Scholar] [CrossRef]
- Talhaoui, N.; Gómez-Caravaca, A.M.; León, L.; de la Rosa, R.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Determination of phenolic compounds of ‘Sikitita’ olive leaves by HPLC-DAD-TOF-MS. Comparison with its parents ‘Arbequina’ and ‘Picual’ olive leaves. LWT-Food Sci. Technol. 2014, 58, 28–34. [Google Scholar] [CrossRef]
- Cavallo, P.; Vinale, F.; Sepe, I.; Galasso, G.; Fedele, F.L.; Sicari, A.; Zito, L.; Lorito, M. Reinforced olive pâté as a source of antioxidants with positive effects on young smokers. Medicina 2019, 55, 680. [Google Scholar] [CrossRef] [Green Version]
- Cavallo, P.; Dini, I.; Sepe, I.; Galasso, G.; Fedele, F.L.; Sicari, A.; Bolletti Censi, S.; Gaspari, A.; Ritieni, A.; Lorito, M.; et al. An innovative olive pâté with nutraceutical properties. Antioxidants 2020, 9, 581. [Google Scholar] [CrossRef]
- Brahmi, F.; Mechri, B.; Dabbou, S.; Dhibi, M.; Hammami, M. The efficacy of phenolics compounds with different polarities as antioxidants from olive leaves depending on seasonal variations. Ind. Crop. Prod. 2012, 38, 146–152. [Google Scholar] [CrossRef]
- Silva, S.; Gomes, L.; Leitao, F.; Coelho, A.V.; Boas, L.V. Phenolic compounds and antioxidant activity of Olea europaea L. fruits and leaves. Food Sci. Technol. Int. 2006, 12, 385–396. [Google Scholar] [CrossRef]
- Bilgin, M.; Şahin, S. Effects of geographical origin and extraction methods on total phenolic yield of olive tree (Olea europaea) leaves. J. Taiwan Inst. Chem. Eng. 2013, 44, 8–12. [Google Scholar] [CrossRef]
- Rafiee, Z.; Jafari, S.M.; Alami, M.; Khomeiri, M. Antioxidant properties of olive leaf extract and its application in sunflower oil. J. Food. Sci. Res. 2011, 21, 12–24. [Google Scholar]
- Japón-Luján, R.; Ruiz-Jiménez, J.; Luque de Castro, M.D. Discrimination and classification of olive tree varieties and cultivation zones by biophenol contents. J. Agric. Food Chem. 2006, 54, 9706–9712. [Google Scholar] [CrossRef]
- Taamalli, A.; Arráez-Román, D.; Ibañez, E.; Zarrouk, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Optimization of microwave-assisted extraction for the characterization of olive leaf phenolic compounds by using HPLC-ESI-TOF-MS/IT-MS(2). J. Agric. Food Chem. 2012, 60, 791–798. [Google Scholar] [CrossRef]
- Talhaoui, N.; Gómez-Caravaca, A.M.; Roldán, C.; León, L.; De la Rosa, R.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Chemometric analysis for the evaluation of phenolic patterns in olive leaves from six cultivars at different growth stages. J. Agric. Food Chem. 2015, 63, 1722–1729. [Google Scholar] [CrossRef]
- Talhaoui, N.; Gómez-Caravaca, A.M.; León, L.; de la Rosa, R.; Fernández-Gutiérrez, A.; Segura-Carretero, A. From olive fruits to olive oil: Phenolic compound transfer in six different olive cultivars grown under the same agronomical conditions. Int. J. Mol. Sci. 2016, 17, 337. [Google Scholar] [CrossRef] [Green Version]
- Olmo-García, L.; Bajoub, A.; Benlamaalam, S.; Hurtado-Fernández, E.; Bagur-González, M.G.; Chigr, M.; Mbarki, M.; Fernández-Gutiérrez, A.; Carrasco-Pancorbo, A. Establishing the Phenolic Composition of Olea europaea L. Leaves from cultivars grown in morocco as a crucial step towards their subsequent exploitation. Molecules 2018, 23, 2524. [Google Scholar] [CrossRef] [Green Version]
- Nicolì, F.; Negro, C.; Vergine, M.; Aprile, A.; Nutricati, E.; Sabella, E.; Miceli, A.; Luvisi, A.; De Bellis, L. Evaluation of phytochemical and antioxidant properties of 15 Italian Olea europaea L. cultivar leaves. Molecules 2019, 24, 1998. [Google Scholar] [CrossRef] [Green Version]
- Rahioui, B.; Aissam, S.; Messaouri, H.; Moukhli, A.; Khadari, B.; Ei, M.C. Role of phenolic metabolism in the defense of the olive-tree against leaf-spot disease caused by Spilocaea oleagina. Int. J. Agric. Biol. 2013, 15, 273–278. [Google Scholar]
- Rahioui, B.; El-Aabidine, A.Z.; Baissac, Y.; El-Boustani, E.; Khadari, B.; Jay Allemand, C.; El-Modafar, C. Phenolic compounds of olive-tree leaves and their relationship with the resistance to the leaf-spot disease caused by Spilocaea oleagina. Am. Eurasian J. Agric. Environ. Sci. 2009, 5, 204–214. [Google Scholar]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Roussos, P.A.; Paplomatas, E.J.; Tjamos, E.C. Phenolic responses of resistant and susceptible olive cultivars induced by defoliating and nondefoliating Verticillium dahlia pathotypes. Plant Dis. 2010, 94, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Petridis, A.; Therios, I.; Samouris, G.; Tananaki, C. Salinity-induced changes in phenolic compounds in leaves and roots of four olive cultivars (Olea europaea L.) and their relationship to antioxidant activity. Environ. Exp. Bot. 2012, 79, 37–43. [Google Scholar] [CrossRef]
- Cetinkaya, H.; Koc, M.; Kulak, M. Monitoring of mineral and polyphenol content in olive leaves under drought conditions: Application chemometric techniques. Ind. Crop. Prod. 2016, 88, 78–84. [Google Scholar] [CrossRef]
- Mechria, B.; Tekayaa, M.; Hammamia, M.; Chehabb, H. Root verbascoside and oleuropein are potential indicators of drought resistance in olive trees (Olea europaea L.). Plant Physiol. Biochem. 2019, 141, 407–414. [Google Scholar] [CrossRef]
- Báidez, A.G.; Gómez, P.; Del Río, J.A.; Ortuño, A. Dysfunctionality of the xylem in Olea europaea L. Plants associated with the infection process by Verticillium dahliae Kleb. Role of phenolic compounds in plant defense mechanism. J. Agric. Food Chem. 2007, 55, 3373–3377. [Google Scholar] [CrossRef]
- El Aabidine, A.Z.; Baissac, Y.; Moukhli, A.; Jay-Allemand, C.; Khadari, B.; El Modafar, C. Resistance of olive tree to Spilocaea oleagina is mediated by the synthesis of phenolic compounds. Int. J. Agric. Biol. 2010, 12, 61–67. [Google Scholar]
- Schrey, S.D.; Tarkka, M.T. Friends and foes: Streptomycetes as modulators of plant disease and symbiosis. Antonie Van Leeuwenhoek 2008, 94, 11–19. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Pradhan, J.K.; Singh, B.P.; Roy, M.; Imram, M.; Pathak, N.; Baisyal, B.M.; Rai, J.P.; et al. Bio-protective microbial agents from rhizosphere eco-systems trigger plant defense responses provide protection against sheath blight disease in rice (Oryza sativa L.). Microbiol. Res. 2016, 192, 300–312. [Google Scholar] [CrossRef]
- Asari, S.; Tarkowska, D.; Rolcik, J.; Novak, O.; Palmero, D.V.; Bejai, S.; Meijer, J. Analysis of plant growth-promoting properties of Bacillus amyloliquefaciens UCMB5113 using Arabidopsis thaliana as hostplant. Planta 2016, 245, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agricolture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: From ‘omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Marra, R.; Nicoletti, R.; Pagano, E.; Della Greca, M.; Salvatore, M.M.; Borrelli, F.; Lombardi, N.; Vinale, F.; Woo, S.L.; Andolfi, A. Inhibitory effect of trichodermanone C, a sorbicillinoid produced by Trichoderma citrinoviride associated to the green alga Cladophora sp., on nitrite production in LPS-stimulated macrophages. Nat. Prod. Res. 2019, 33, 3389–3397. [Google Scholar] [CrossRef]
- Vinale, F.; Flematti, G.; Sivasithamparam, K.; Lorito, M.; Marra, R.; Skelton, B.W.; Ghisalberti, E.L. Harzianic acid, an antifungal and plant growth promoting metabolite from Trichoderma harzianum. J. Nat. Prod. 2009, 72, 2032–2035. [Google Scholar] [CrossRef]
- Vinale, F.; Arjona Girona, I.; Nigro, M.; Mazzei, P.; Piccolo, A.; Ruocco, M.; Woo, S.L.; David Ruano Rosa, D.; López Herrera, C.; Lorito, M. Cerinolactone, a hydroxy-lactone derivative from Trichoderma Cerinum. J. Nat. Prod. 2011, 75, 103–106. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.L.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Res. 2012, 7, 1545–1550. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Strakowska, J.; Mazzei, P.; Piccolo, A.; Marra, R.; Lombardi, N.; Manganiello, G.; Pascale, A.; Woo, S.L.; Lorito, M. Cremenolide, a new antifungal, 10-member lactone from Trichoderma cremeum with plant growth promotion activity. Nat. Prod. Res. 2016, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Ruocco, M.; Manganiello, G.; Guerrieri, E.; Bernardo, U.; Mazzei, P.; Piccolo, A.; Sannino, F.; Caira, S.; Woo, S.L.; et al. Metabolites produced by Gnomoniopsis castanea associated with necrosis of chestnut galls. Chem. Biol. Technol. 2014, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Manganiello, G.; Sacco, A.; Ercolano, M.R.; Vinale, F.; Lanzuise, S.; Pascale, A.; Woo, S.L. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front. Microbiol. 2018, 9, 1966. [Google Scholar] [CrossRef]
- Carrero-Carrón, I.; Trapero-Casas, J.L.; Olivares-García, C.; Monte, E.; Hermosa, R.; Jiménez-Díaz, R.M. Trichoderma asperellum is effective for biocontrol of Verticillium wilt in olive caused by the defoliating pathotype of Verticillium dahliae. Crop. Prot. 2016, 88, 45–52. [Google Scholar] [CrossRef]
- Marra, R.; Lombardi, N.; d’Errico, G.; Troisi, J.; Scala, G.; Vinale, F.; Woo, S.L.; Bonanomi, G.; Lorito, M. Application of Trichoderma strains and metabolites enhances soybean productivity and nutrient content. J. Agric. Food Chem. 2019, 67, 1814–1822. [Google Scholar] [CrossRef]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef]
- Vinale, F.; Nicoletti, R.; Lacatena, F.; Marra, R.; Sacco, A.; Lombardi, N.; d’Errico, G.; Digilio, M.C.; Lorito, M.; Woo, S.L. Secondary metabolites from the endophytic fungus Talaromyces pinophilus. Nat. Prod. Res. 2017, 31, 1778–1785. [Google Scholar] [CrossRef] [Green Version]
- Loprieno, N.; Cenerini, I. Metodo per la diagnosi precoce dell’occhio di pavone dell’olivo Cycloconium oleaginum (Cast.). Phytopathology 1959, 34, 385–392. [Google Scholar] [CrossRef]
- Yamamoto, A.; Nakamura, T.; Adu-Gyamfi, J.J.; Saigusa, M. Relationship between chlorophyll content in leaves of sorghum and pigeonpea determined by extraction method and by chlorophyll meter (SPAD-502). J. Plant Nutr. 2002, 25, 2295–2301. [Google Scholar] [CrossRef]
- Corrado, G.; Alagna, F.; Rocco, M.; Renzone, G.; Varricchio, P.; Coppola, V.; Coppola, M.; Garonna, A.; Baldoni, L.; Scaloni, A.; et al. Molecular interactions between the olive and the fruit fly Bactrocera oleae. BMC Plant Biol. 2012, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tafuri, S.; Cocchia, N.; Carotenuto, D.; Vassetti, A.; Staropoli, A.; Mastellone, V.; Peretti, V.; Ciotola, F.; Albarella, S.; Del Prete, C.; et al. Chemical Analysis of Lepidium meyenii (Maca) and Its Effects on Redox Status and on Reproductive Biology in Stallions. Molecules 2019, 24, 1981. [Google Scholar] [CrossRef] [Green Version]
- Obanor, F.O.; Walter, M.; Jones, E.E.; Jaspers, M.V. Effect of temperature, relative humidity, leaf wetness and leaf age on Spilocaea oleagina conidium germination on olive leaves. Eur. J. Plant Pathol. 2008, 120, 211–222. [Google Scholar] [CrossRef]
- Otero, M.L.; Roca, M.; Zapata, R.; Ladux, J.L.; Ortiz, J.; Zanelli, M.; Matías, A.C.; Pérez, B.A. Effect of solarization, organic matter, and Trichoderma on the severity of verticillium wilt in olive trees (Olea europaea L.) and soil inoculum density. Acta Hortic. 2014, 1057, 121–126. [Google Scholar] [CrossRef]
- Amira, M.B.; Lopez, D.; Mohamed, A.T.; Khouaja, A.; Chaar, H.; Fumana, L.B.; Gousset-Dupont, A.; Bonhomme, L.; Label, P.; Goupil, P.; et al. Beneficial effect of Trichoderma harzianum strain Ths97 in biocontrolling Fusarium solani causal agent of root rot disease in olive trees. Biol. Control. 2017, 110, 70–78. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Prieto, P.; Rincón, A.M.; Gómez-Rodríguez, M.V.; Valderrama, R.; Barroso, J.B.; Mercado-Blanco, J. Fate of Trichoderma harzianum in the olive rhizosphere: Time course of the root colonization process and interaction with the fungal pathogen Verticillium Dahliae. BioControl 2016, 61, 269–282. [Google Scholar] [CrossRef]
- Porras, M.; Barrau, C.; Arroyo, F.T.; Santos, B.; Blanco, C.; Romero, F. Reduction of Phytophthora cactorum in strawberry fields by Trichoderma spp. and soil solarization. Plant Dis. 2007, 91, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Morsy, E.M.; Abdel-Kawi, K.A.; Khalil, M.N.A. Efficacy of Trichoderma viride and Bacillus subtilis as biocontrol agents against Fusarium solani on tomato plants. Egypt J. Phytopathol. 2009, 37, 47–57. [Google Scholar]
- Shanmugam, H.; Rathinam, R.; Chinnathambi, A.; Venkatesan, T. Antimicrobial and mutagenic properties of the root tubers of Gloriosa superba. Pak. J. Bot. 2009, 41, 293–299. [Google Scholar]
- John, R.P.; Tyagi, R.D.; Prevost, D.; Brar, S.K.; Pouleur, S.; Surampalli, R.Y. Mycoparasitic Trichoderma viride as a biocontrol agent against Fusarium oxysporumf sp. spadzuki and Pythium arrhenomanes and as a growth promoter of soybean. Crop. Prot. 2010, 29, 1452–1459. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Kathiresan, K. Thermostable cellulase produced by Trichoderma (Hypocrea estonica). J. Biotechnol. Sci. 2013, 1, 22–41. [Google Scholar]
- Doni, F.; Anizan, I.; Che Radziah, C.M.Z.; Salman, A.H.; Rodzihan, M.H.; Wan Mohtar, W.Y. Enhancement of rice seed germination and vigour by Trichodermaspp. Res. J. App. Sci. Eng. Technol. 2014, 7, 4547–4552. [Google Scholar] [CrossRef]
- Pascale, A.; Vinale, F.; Manganiello, G.; Nigro, M.; Lanzuise, S.; Ruocco, M.; Marra, R.; Lombardi, N.; Woo, S.L.; Lorito, M. Trichoderma and its secondary metabolites improve yield and quality of grapes. Crop. Prot. 2017, 92, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. Transcriptome and Metabolome Reprogramming in Tomato Plants by Trichoderma harzianum strain T22 Primes and Enhances Defense Responses Against Aphids. Front. Physiol. 2019, 10, 745. [Google Scholar] [CrossRef]
- Clarke, J.D.; Volko, S.M.; Ledford, H.; Ausubel, F.M.; Dong, X. Roles of salicylic acid, jasmonic acid, and ethylene in cpr-induced resistance in Arabidopsis. Plant Cell 2000, 12, 2175–2190. [Google Scholar] [CrossRef] [Green Version]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Walling, L.L. Adaptive defense responses to pathogens and insects. In Advances in Botanical Research; Loon, L.V., Ed.; Elsevier: London, UK, 2009; Volume 51, pp. 551–612. [Google Scholar]
- Grasso, F.; Coppola, M.; Carbone, F.; Baldoni, L.; Alagna, F.; Perrotta, G.; Pérez-Pulido, A.J.; Garonna, A.; Facella, P.; Daddiego, L.; et al. The transcriptional response to the olive fruit fly (Bactrocera oleae) reveals extended differences between tolerant and susceptible olive (Olea europaea L.) varieties. PLoS ONE 2017, 12, e0183050. [Google Scholar] [CrossRef] [Green Version]
- Heyman, J.; Canher, B.; Bisht, A.; Christiaens, F.; De Veylder, L. Emerging role of the plant ERF transcription factors in coordinating wound defense responses and repair. J. Cell Sci. 2017, 131, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Munné-Bosch, S. Ethylene response factors: A key regulatory hub in hormone and stress signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell 2002, 14, S153–S164. [Google Scholar] [CrossRef] [Green Version]
- Cha, Y.; Barman, D.N.; Kim, M.G.; Kim, W.Y. Stress defense mechanisms of NADPH-dependent thioredoxin reductases (NTRs) in plants. Plant Signal. Behav. 2015, 10, e017698. [Google Scholar] [CrossRef]
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Canini, A. Plant defense factors involved in Olea europaea resistance against Xylella fastidiosa infection. J. Plant Res. 2019, 132, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esti, M.; Cinquanta, L.; La Notte, E. Phenolic compounds in different olive varieties. J. Agric. Food Chem. 1998, 46, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Bruno, L.; Picardi, E.; Pacenza, M.; Chiappetta, A.; Muto, A.; Gagliardi, O.; Muzzalupo, I.; Pesole, G.; Bitonti, M.B. Changes in gene expression and metabolic profile of drupes of Oleaeuropaea L. cv Carolea in relation to maturation stage and cultivation area. BMC Plant Biol. 2019, 19, 428. [Google Scholar] [CrossRef]
- Pedan, V.; Popp, M.; Rohn, S.; Nyfeler, M.; Bongartz, A. Characterization of phenolic compounds and their contribution to sensory properties of olive oil. Molecules 2019, 24, 2041. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Soler-Rivas, C.; Espin, J.C.; Wichers, H.J. Oleuropein and related compounds. J. Sci. Food Agric. 2000, 80, 1013–1023. [Google Scholar] [CrossRef]
- Mayo-Prieto, S.; Marra, R.; Vinale, F.; Rodríguez-González, Á.; Woo, S.L.; Lorito, M.; Casquero, P.A. Effect of Trichoderma velutinum and Rhizoctonia solani on the Metabolome of Bean Plants (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2019, 20, 549. [Google Scholar] [CrossRef] [Green Version]
- Mazzei, P.; Vinale, F.; Woo, S.L.; Pascale, A.; Lorito, M.; Piccolo, A. Metabolomics by proton high-resolution magic-angle-spinning nuclear magnetic resonance of tomato plants treated with two secondary metabolites isolated from Trichoderma. J. Agric. Food Chem. 2016, 64, 3538–3545. [Google Scholar] [CrossRef] [Green Version]
- Gorrochategui, E.; Jaumot, J.; Lacorte, S.; Tauler, R. Data analysis strategies for targeted and untargeted LC-MS metabolomic studies: Overview and workflow. Trends Anal. Chem. 2016, 80, 425–442. [Google Scholar] [CrossRef]
- Amiot, M.J.; Fleuriet, A.; Macheix, J.J. Importance and evolution of phenolic compounds in olive during growth and maturation. J. Agric. Food Chem. 1986, 34, 823–826. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Freitas, H.; Santos, C.; Silva, A.M.S. The antioxidant system in Olea europaea to enhanced UV-B radiation also depends on flavonoids and secoiridoids. Phytochemistry 2020, 170, 112199. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. | RT (min) | Compound | Group * | Formula | Experimental Mass | Theoretical Mass | UV Max (nm) |
|---|---|---|---|---|---|---|---|
| 1 | 4.75 | Hydroxytyrosol-hexose | 3 | C14H20O8 | 316.1160 | 315.10 | 230; 280 |
| 2 | 6.33 | Oleoside methyl ester | 4 | C17H24O11 | 404.1321 | 404.36 | 235 |
| 3 | 7.45 | Secologanoside | 4 | C16H22O11 | 390.1151 | 390.34 | 234 |
| 4 | 10.90 | Oleuropeinaglycon | 1 | C16H26016 | 378.1569 | 378.38 | 235; 271 |
| 5 | 11.84 | Elenolic acid glucoside | 5 | C17H24011 | 404.1325 | 404.13 | 248; 274 |
| 6 | 11.90 | Luteolin rutinoside is. a | 2 | C27H30015 | 594.1589 | 594.52 | 248; 267 |
| 7 | 12.25 | Luteolin di-glucoside | 2 | C27H30016 | 610.1537 | 610.15 | 248; 267; 335 |
| 8 | 14.55 | Verbascoside | 3 | C29H36O15 | 624.2064 | 624.59 | 234; 329 |
| 9 | 14.60 | Rutin | 2 | C27H30016 | 610.1539 | 610.52 | 253 |
| 10 | 14.90 | Luteolin rutinoside is. b | 2 | C27H30015 | 594.1592 | 594.52 | 253; 347 |
| 11 | 15.18 | Chrysoeriol-7-O-glucoside | 2 | C22H22O11 | 462.1497 | 462.40 | 250; 347 |
| 12 | 15.81 | 2-Methoxyoleuropein | 1 | C26H34O14 | 570.1942 | 570.17 | 236; 280 |
| 13 | 16.90 | Apigeninrutinoside | 2 | C27H30014 | 578.1640 | 578.16 | 237; 266 |
| 14 | 17.40 | Apigenin glucoside | 2 | C21H20O10 | 432.1061 | 432.38 | 345 |
| 15 | 17.90 | Oleuropeindiglucoside | 1 | C31H24O18 | 702.2400 | 702.66 | 235; 277 |
| 16 | 19.10 | Oleuropein isomer a | 1 | C25H32O13 | 540.1840 | 540.18 | 240; 280 |
| 17 | 19.89 | Oleuroside | 1 | C25H32O13 | 540.1845 | 540.51 | 230; 280 |
| 18 | 20.10 | Oleuropein isomer b | 1 | C25H32O13 | 540.1848 | 540.18 | 235; 280 |
| 19 | 20.52 | Ligstroside | 1 | C25H32012 | 524.1900 | 524.52 | 230; 280 |
| 20 | 20.93 | Apigenin | 2 | C15H10O5 | 270.0530 | 270.24 | 268; 334 |
| 21 | 20.80 | Luteolin | 2 | C15H35O14 | 286.0488 | 286.05 | 255; 286 |
| 22 | 23.28 | Chrysoeriol | 2 | C16H12O6 | 300.0682 | 300.26 | 198 |
| Trichoderma | Treatment | UP vs. CTRL | DOWN vs. CTRL |
|---|---|---|---|
| Strain | GV41 | 3 | 1 |
| M10 | 1 | 7 | |
| T22 | 2 | 3 | |
| TH1 | 0 | 0 | |
| KV906 | 1 | 5 | |
| Metabolite | HA | 1 | 9 |
| 6PP | 2 | 15 |
| TREATMENT | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| RT (min) | Compound | Mass | CTRL | M10 | KV906 | GV41 | TH1 | T22 | 6PP | HA |
| 4.75 | Hydroxytyrosol-hexose | 316 | 0.947 (0.700) b,c | 0.448 (0.198) a | 0.391 (0.087) a | 0.515 (0.265) a,b | 1.203 (0.833) c | 0.350 (0.146) b,c | 0.241 (0.162) a | 0.341 (0.130) a |
| 7.45 | Secologanoside | 390 | 0.320 (0.020) b | 0.280 (0.040) a | 0.152 (0.024) a | 0.240 (0.072) a | 0.418 (0.022) b | 0.160 (0.022) a | 0.160 (0.040) a | 0.080 (0.064) a |
| 10.90 | Oleuropein glycon | 378 | 0.038 (0.001) a,c | 0.045 (0.002) b,c,d,e | 0.041 (0.002) a,d | 0.035 (0.001) a | 0.054 (0.010) e | 0.036 (0.001) a,b | 0.051 (0.004) d,e | 0.032 (0.001) a |
| 11.84 | Elenolic acid glucoside | 404 | 0.206 (0.058) a,b | 0.174 (0.003) a,b | 0.214 (0.002) a,b | 0.137 (0.003) a,b | 0.167 (0.004) a,b | 0.125 (0.002) a | 0.227 (0.095) b | 0.344 (0.048) c |
| 11.90 | Luteolin rutinoside is. a | 594 | 0.312 (0.160) a,b | 0.360 (0.032) c | 0.240 (0.016) a | 0.320 (0.016) a,b | 0.344 (0.032) b,c | 0.280 (0.005) a | 0.280 (0.024) a | 0.304 (0.040) a,b |
| 12.25 | Luteolin diglucoside | 610 | 0.384 (0.040) a,b | 0.600 (0.056) d | 0.376 (0.016) a | 0.440 (0.048) c | 0.464 (0.064) c,d | 0.392 (0.024) a,b | 0.416 (0.072) b,c | 0.360 (0.016) a |
| 14.60 | Rutin | 610 | 0.211 (1.192) c,d | 0.119 (0.040) b,c,d | 0.048 (0.036) a,b | 0.077 (0.038) b,c,d | 0.257 (0.102) d | 0.050 (0.035) a,b | 0.016 (0.020) a | 0.068 (0.040) a,c |
| 14.90 | Luteolin rutinosideis. b | 594 | 0.329 (0.026) c,d | 0.313 (0.016) c,d | 0.139 (0.011) a,b | 0.231 (0.021) b,c | 0.411 (0.037) d | 0.253 (0.018) b,c | 0.047 (0.003) a | 0.177 (0.014) a,b |
| 15.18 | Chrysoeriol-7-O-glucoside | 462 | 0.007 (0.001) a | 0.035 (0.002) a,b | 0.047 (0.004) b | 0.036 (0.003) a,b | 0.024 (0.002) a,b | 0.014 (0.001) a,b | 0.045 (0.003) b | 0.027 (0.002) a,b |
| 15.81 | 2-Methoyxyoleuropein | 570 | 0.400 (0.032) a,c | 0.160 (0.008) b,c | 0.152 (0.012) b,c | 0.500 (0.045) d | 0.280 (0.025) a,c | 0.400 (0.028) a,c | 0.080 (0.005) a,b | 0.064 (0.005) a |
| 16.90 | Apigenin rutinoside | 578 | 0.382 (0.040) a | 0.574 (0.040) b | 0.517 (0.034) a,b | 0.447 (0.037) a,b | 0.492 (0.038) a,b | 0.439 (0.020) a,b | 0.557 (0.060) b | 0.563 (0.044) b |
| 17.40 | Apigenin glucoside | 432 | 0.064 (0.004) a | 0.088 (0.006) c | 0.080 (0.007) a,b | 0.084 (0.006) b,c | 0.072 (0.004) a,b | 0.068 (0.005) a,b | 0.096 (0.012) c | 0.064 (0.006) a |
| 17.90 | Oleuropein diglucoside | 702 | 0.680 (0.320) c | 0.520 (0.280) b | 0.480 (0.056) a,b | 0.536 (0.048) b,c | 0.600 (0.320) b,c | 0.440 (0.048) a,b | 0.400 (0.060) a | 0.416 (0.056) a |
| 19.10 | Oleuropein isomer a | 540 | 8.428 (0.849) b,c | 2.106 (0.068) a | 2.192 (0.169) a,b | 1.608 (0.044) a | 11.694 (1.171) c | 0.933 (0.023) a | 0.480 (0.036) a | 0.646 (0.052) a |
| 20.10 | Oleuropein isomer b | 540 | 1.062 (0.085) b | 0.144 (0.007) a | 0.160 (0.013) a | 0.120 (0.011) a | 1.661 (0.150) b | 0.080 (0.006) a | 0.048 (0.003) a | 0.080 (0.006) a |
| 20.80 | Luteolin | 286 | 0.129 (0.049) c | 0.088 (0.031) a,c | 0.086 (0.023) a,b,c | 0.068 (0.004) a,b | 0.119 (0.042) c | 0.101 (0.021) b,c | 0.049 (0.002) a | 0.119 (0.003) c |
| 23.28 | Chrysoeriol | 300 | 0.053 (0.004) a,b,c | 0.036 (0.002) a | 0.084 (0.007) b,c,d | 0.040 (0.004) a,b | 0.096 (0.009) c,d | 0.048 (0.003) a,b | 0.038 (0.002) a,b | 0.104 (0.008) d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marra, R.; Coppola, M.; Pironti, A.; Grasso, F.; Lombardi, N.; d’Errico, G.; Sicari, A.; Bolletti Censi, S.; Woo, S.L.; Rao, R.; et al. The Application of Trichoderma Strains or Metabolites Alters the Olive Leaf Metabolome and the Expression of Defense-Related Genes. J. Fungi 2020, 6, 369. https://doi.org/10.3390/jof6040369
Marra R, Coppola M, Pironti A, Grasso F, Lombardi N, d’Errico G, Sicari A, Bolletti Censi S, Woo SL, Rao R, et al. The Application of Trichoderma Strains or Metabolites Alters the Olive Leaf Metabolome and the Expression of Defense-Related Genes. Journal of Fungi. 2020; 6(4):369. https://doi.org/10.3390/jof6040369
Chicago/Turabian StyleMarra, Roberta, Mariangela Coppola, Angela Pironti, Filomena Grasso, Nadia Lombardi, Giada d’Errico, Andrea Sicari, Sergio Bolletti Censi, Sheridan L. Woo, Rosa Rao, and et al. 2020. "The Application of Trichoderma Strains or Metabolites Alters the Olive Leaf Metabolome and the Expression of Defense-Related Genes" Journal of Fungi 6, no. 4: 369. https://doi.org/10.3390/jof6040369
APA StyleMarra, R., Coppola, M., Pironti, A., Grasso, F., Lombardi, N., d’Errico, G., Sicari, A., Bolletti Censi, S., Woo, S. L., Rao, R., & Vinale, F. (2020). The Application of Trichoderma Strains or Metabolites Alters the Olive Leaf Metabolome and the Expression of Defense-Related Genes. Journal of Fungi, 6(4), 369. https://doi.org/10.3390/jof6040369