Light-Photoreceptors and Proteins Related to Monilinia laxa Photoresponses

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Monilinia laxa Strains and Culture Conditions
2.2. Effect of Light on Monilinia laxa Colony Growth and Conidiation
2.3. Identification of Putative Photoreceptors
2.4. Light Transcriptomic Analyses
2.5. Statistical Analysis
3. Results
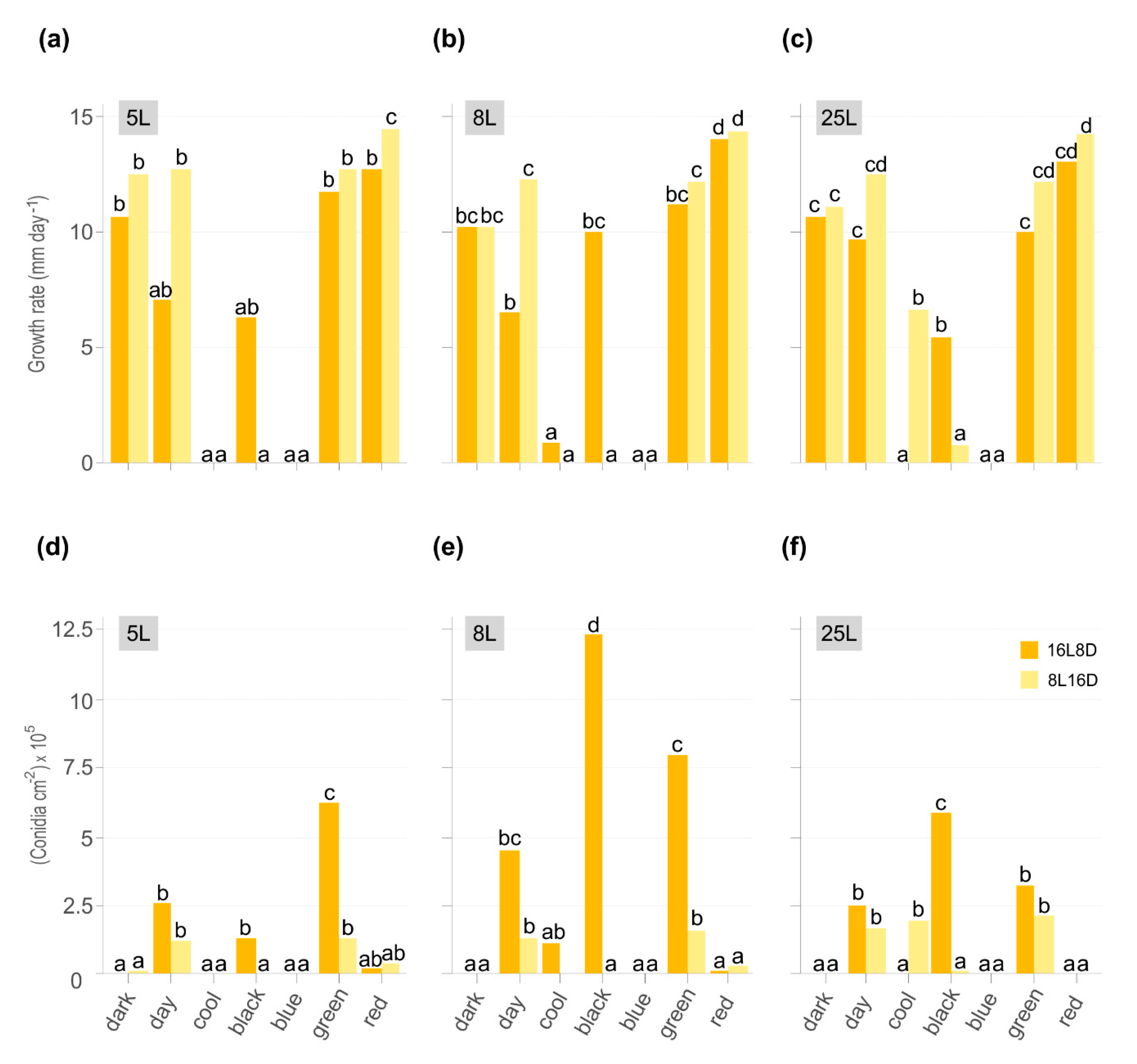
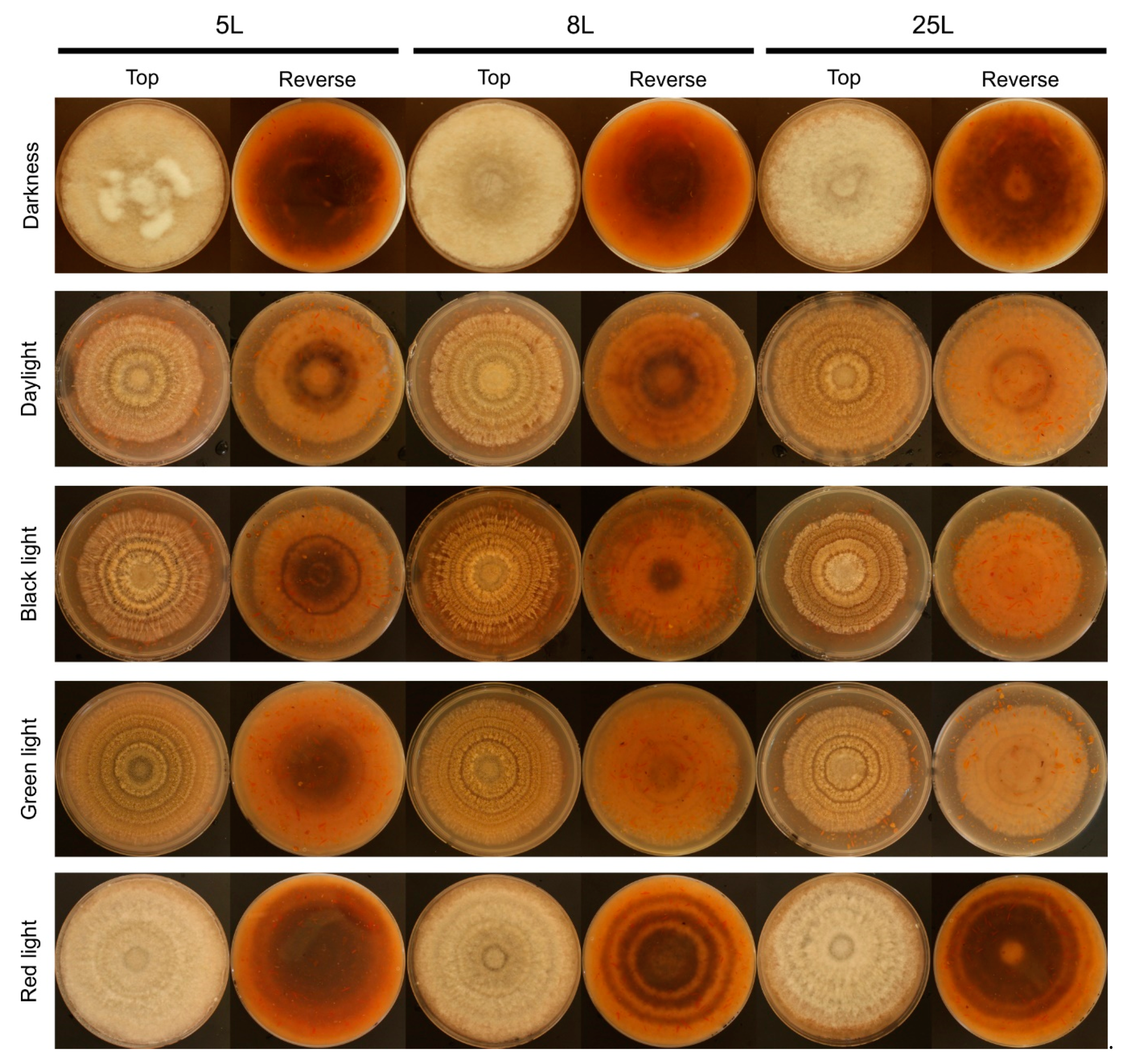
3.1. Effect of Light on Monilinia Laxa Colony Growth, Conidiation, and Morphology
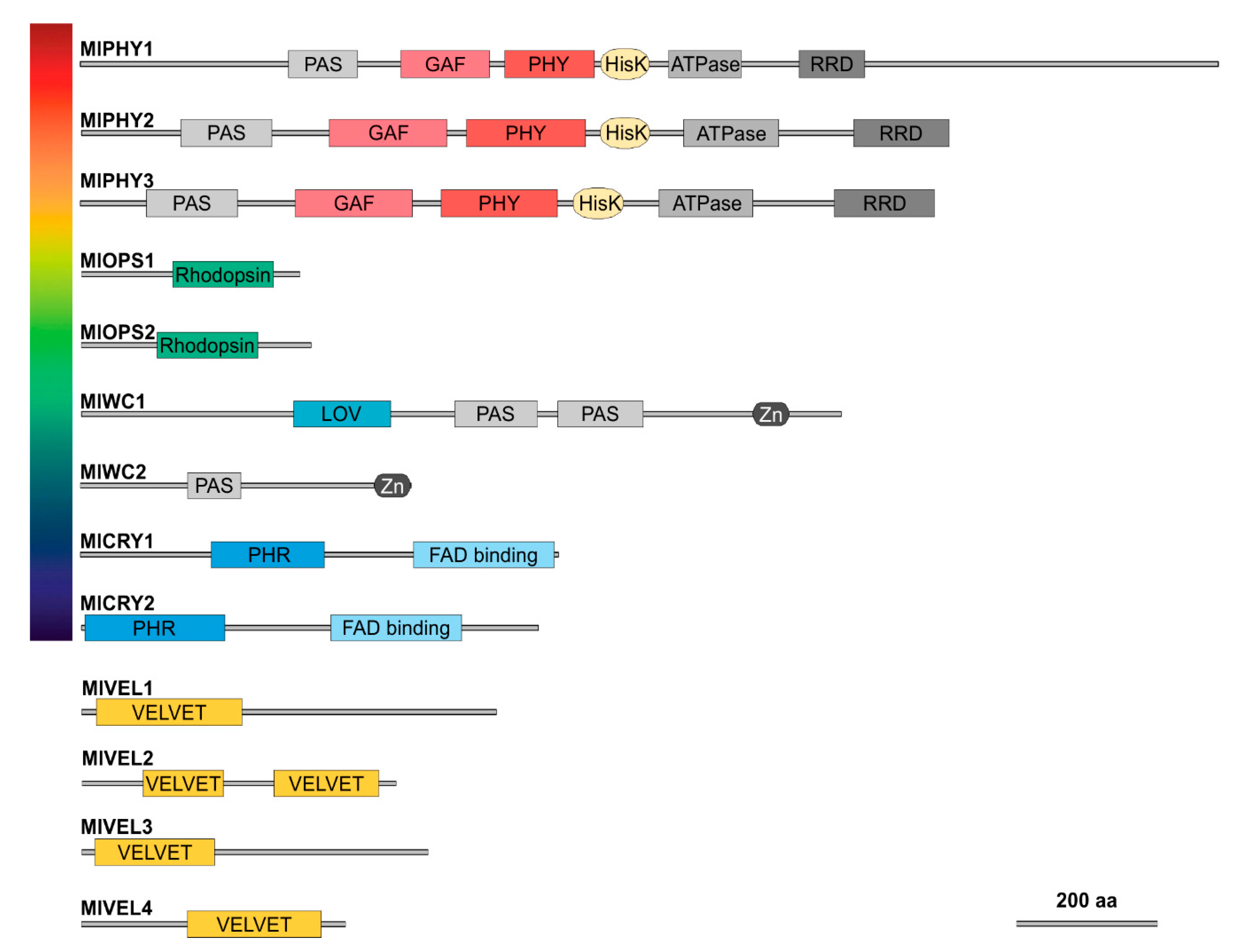
3.2. Identification of Putative Photoreceptors
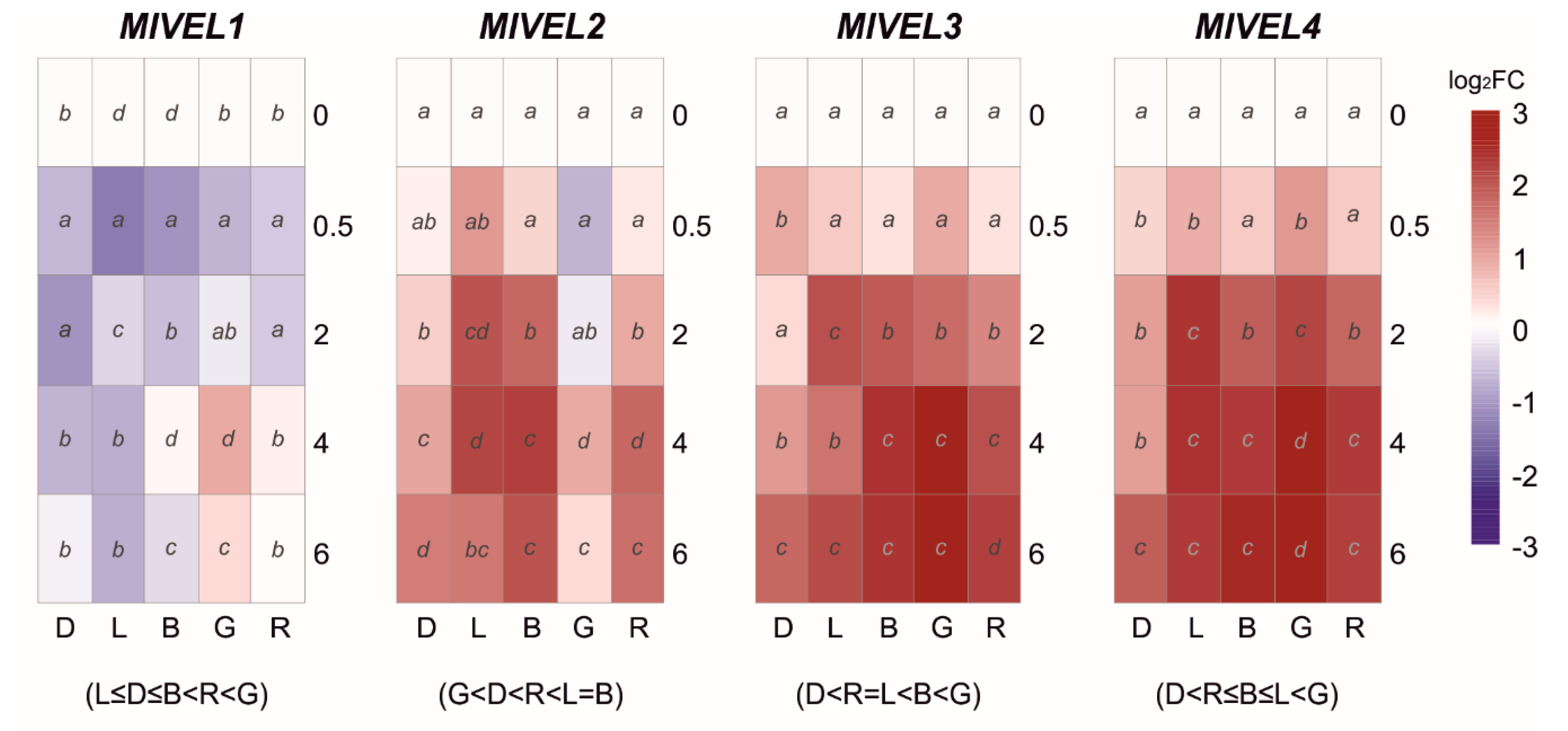
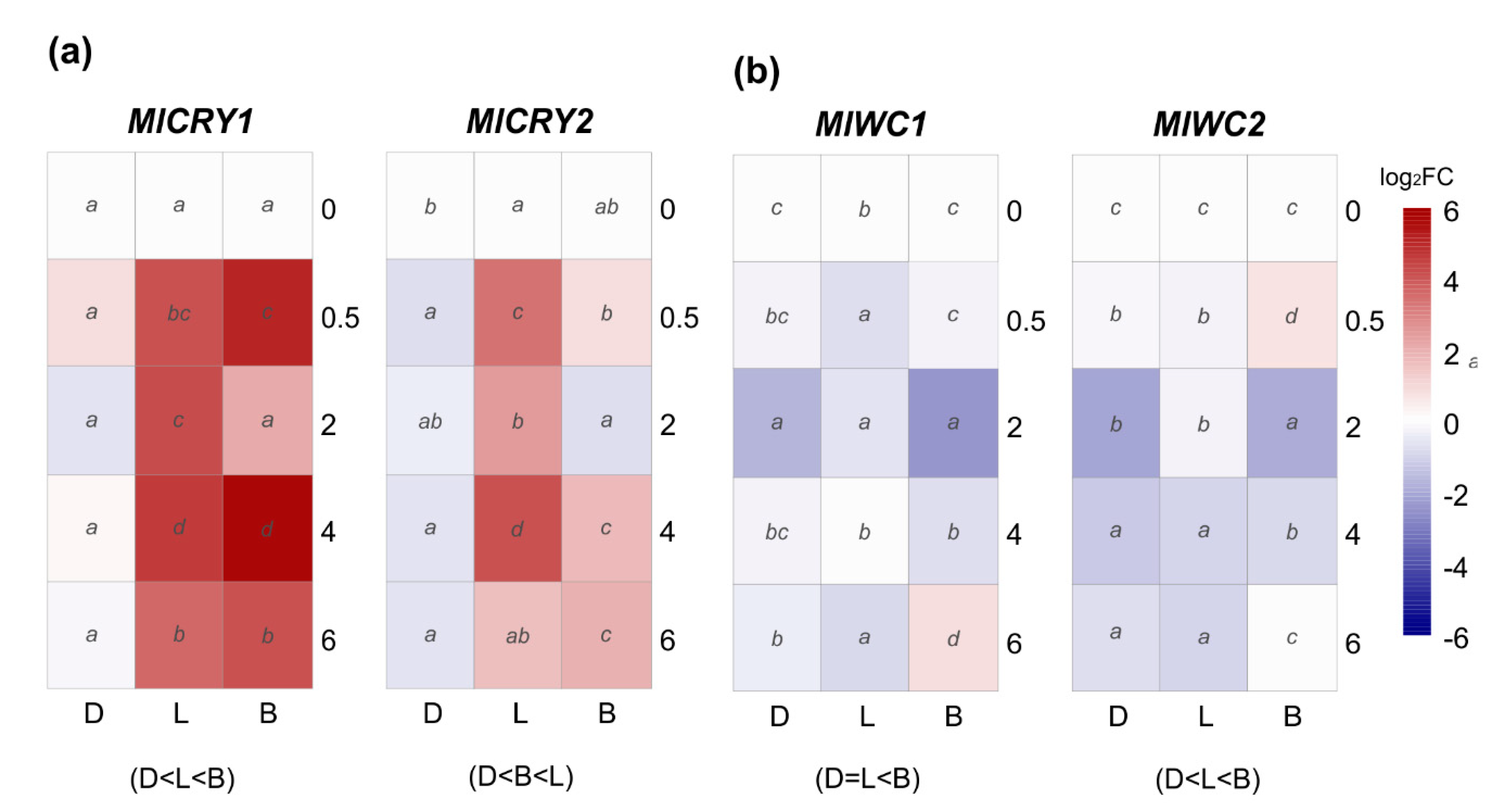
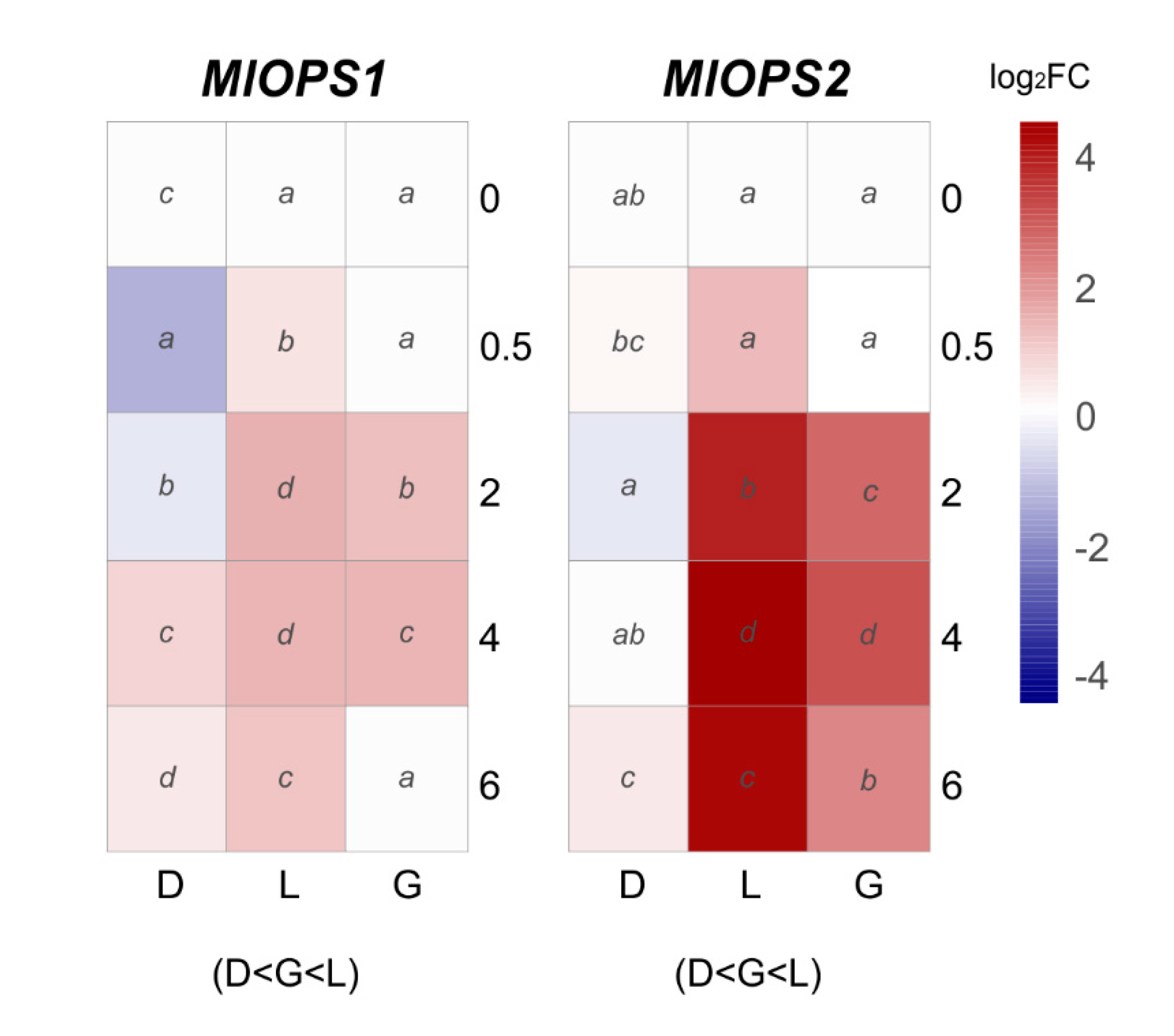
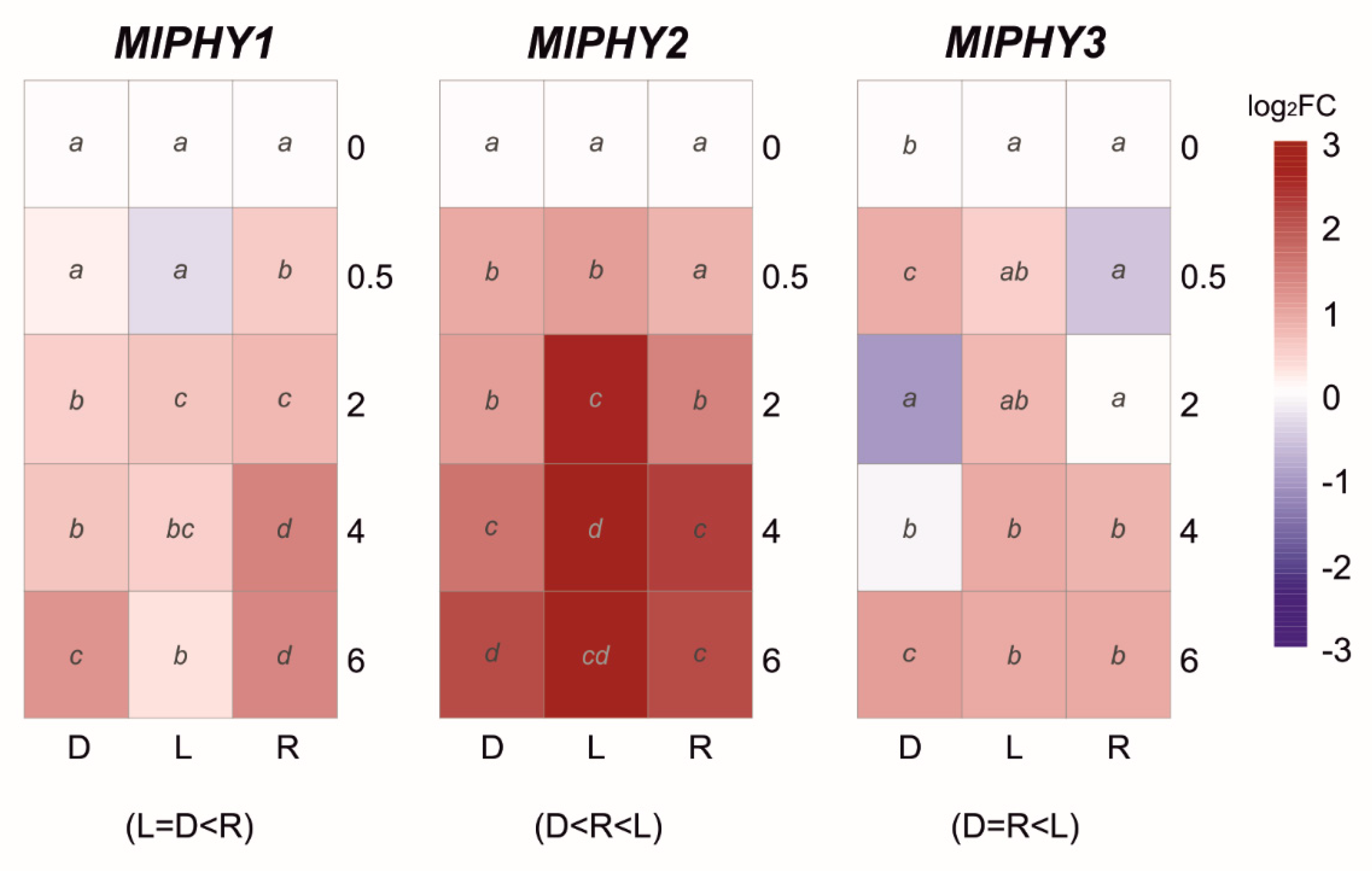
3.3. Effect of Light on the Transcriptional Pattern of Velvet Gene Family and Identified Photoreceptors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byrde, R.J.W.; Willetts, H.J. The Brown Rot Fungi of Fruit: Their Biology and Control; Pergamon Press: London, UK, 1977. [Google Scholar]
- Gell, I.; de Cal, A.; Torres, R.; Usall, J.; Melgarejo, P. Conidial density of Monilinia spp. on peach fruit surfaces in relation to the incidences of latent infections and brown rot. Eur. J. Plant Pathol. 2009, 123, 415–424. [Google Scholar] [CrossRef]
- Villarino, M.; Melgarejo, P.; Usall, J.; Segarra, J.; Lamarca, N.; de Cal, A. Secondary inoculum dynamics of Monilinia spp. and relationship to the incidence of postharvest brown rot in peaches and the weather conditions during the growing season. Eur. J. Plant Pathol. 2012, 133, 585–598. [Google Scholar] [CrossRef]
- Villarino, M.; Melgarejo, P.; Usall, J.; Segarra, J.; de Cal, A. Primary inoculum sources of Monilinia spp. in Spanish peach orchards and their relative importance in brown rot. Plant Dis. 2010, 94, 1048–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannon, F.; Gort, G.; van Leeuwen, G.; Holb, I.; Jeger, M. Diurnal patterns in dispersal of Monilinia fructigena conidia in an apple orchard in relation to weather factors. Agric. For. Meteorol. 2009, 149, 518–525. [Google Scholar] [CrossRef]
- Xu, X.-M.; Guerin, L.; Robinson, J.D. Effects of temperature and relative humidity on conidial germination and viability, colonization and sporulation of Monilinia fructigena. Plant Pathol. 2001, 50, 561–568. [Google Scholar] [CrossRef]
- Idnurm, A.; Crosson, S. The photobiology of microbial pathogenesis. PLoS Pathog. 2009, 5, 11–13. [Google Scholar] [CrossRef] [Green Version]
- Roden, L.C.; Ingle, R.A. Lights, rhythms, infection: The role of light and the circadian clock in determining the outcome of plant-pathogen interactions. Plant. Cell 2009, 21, 2546–2552. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.D.; Castillo, J.A. Influence of light on plant–phyllosphere interaction. Front. Plant. Sci. 2018, 9, 1482. [Google Scholar] [CrossRef] [Green Version]
- Canessa, P.; Schumacher, J.; Hevia, M.A.; Tudzynski, P.; Larrondo, L.F. Assessing the effects of light on differentiation and virulence of the plant pathogen Botrytis cinerea: Characterization of the white collar complex. PLoS ONE 2013, 8, e84223. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Traeger, S.; Porquier, A.; Dalmais, B.; Viaud, M.; Tudzynski, B. The VELVET complex in the gray mold fungus Botrytis cinerea: Impact of BcLAE1 on differentiation, secondary metabolism, and virulence. Mol. Plant Microbe Interact. 2015, 28, 659–674. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. The microbial opsin homolog sop1 is involved in Sclerotinia sclerotiorum development and environmental stress response. Front. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.K. Interaction of near-ultraviolet, blue, red, and far-red light in sporulation of Botrytis cinerea. Trans. Br. Mycol. Soc. 1975, 64, 215–222. [Google Scholar] [CrossRef]
- Zhu, P.; Zhang, C.; Xiao, H.; Wang, Y.; Toyoda, H.; Xu, L. Exploitable regulatory effects of light on growth and development of Botrytis cinerea. J. Plant Pathol. 2013, 95, 509–517. [Google Scholar] [CrossRef]
- De Cal, A.; Melgarejo, P. Effects of long-wave UV light on Monilinia growth and identification of species. Plant Dis. 1999, 83, 62–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Pires, S.; Garcia-Companys, M.; Espeso, E.A.; Melgarejo, P.; de Cal, A. Influence of light on the Monilinia laxa–stone fruit interaction. Plant Pathol. 2020. [Google Scholar] [CrossRef]
- Fischer, R.; Aguirre, J.; Herrera-Estrella, A.; Corrochano, L.M. The complexity of fungal vision. In The Fungal Kingdom; ASM Press: Washington, DC, USA, 2017; pp. 441–461. [Google Scholar]
- Fuller, K.K.; Loros, J.J.; Dunlap, J.C. Fungal photobiology: Visible light as a signal for stress, space and time. Curr. Genet. 2015, 61, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Romero, J.; Hedtke, M.; Kastner, C.; Müller, S.; Fischer, R. Fungi, hidden in soil or up in the air: Light makes a difference. Annu. Rev. Microbiol. 2010, 64, 585–610. [Google Scholar] [CrossRef] [Green Version]
- Corrochano, L.M. Light in the fungal world: From photoreception to gene transcription and beyond. Annu. Rev. Genet. 2019, 53, 149–170. [Google Scholar] [CrossRef]
- Yu, Z.; Fischer, R. Light sensing and responses in fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Braus, G.H.; Fischer, R.; Rodriguez-Romero, J. Spotlight on Aspergillus nidulans photosensory systems. Fungal Genet. Biol. 2010, 47, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Dunlap, J.C.; Loros, J.J. Neurospora illuminates fungal photoreception. Fungal Genet. Biol. 2010, 47, 922–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, A.; Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Seeing the world differently: Variability in the photosensory mechanisms of two model fungi. Environ. Microbiol. 2016, 18, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooney, J.L.; Yager, L.N. Light is required for conidiation in Aspergillus nidulans. Genes Dev. 1990, 4, 1473–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stinnett, S.M.; Espeso, E.A.; Cobeño, L.; Araújo-Bazán, L.; Calvo, A.M. Aspergillus nidulans VeA subcellular localization is dependent on the importin α carrier and on light. Mol. Microbiol. 2007, 63, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef]
- Etxebeste, O.; Otamendi, A.; Garzia, A.; Espeso, E.A.; Cortese, M.S. Rewiring of transcriptional networks as a major event leading to the diversity of asexual multicellularity in fungi. Crit. Rev. Microbiol. 2019, 45, 548–563. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Verma, S.; Corrochano, L.M. A glimpse into the basis of vision in the kingdom Mycota. Fungal Genet. Biol. 2010, 47, 881–892. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J. How light affects the life of Botrytis. Fungal Genet. Biol. 2017, 106, 26–41. [Google Scholar] [CrossRef]
- Angelini, R.M.d.M.; Abate, D.; Rotolo, C.; Gerin, D.; Pollastro, S.; Faretra, F. De novo assembly and comparative transcriptome analysis of Monilinia fructicola, Monilinia laxa and Monilinia fructigena, the causal agents of brown rot on stone fruits. BMC Genom. 2018, 19, 1–21. [Google Scholar] [CrossRef]
- Naranjo-Ortíz, M.A.; Rodríguez-Pires, S.; Torres, R.; de Cal, A.; Usall, J.; Gabaldón, T. Genome sequence of the brown rot fungal pathogen Monilinia laxa. Genome Announc. 2018, 6, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Landi, L.; Pollastro, S.; Rotolo, C.; Romanazzi, G.; Faretra, F.; Angelini, R.M.d.M. Draft genomic resources for the brown rot fungal pathogen Monilinia laxa. Mol. Plant Microbe Interact. 2020, 33, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Pires, S.; Melgarejo, P.; de Cal, A.; Espeso, E.A. Pectin as carbon source for Monilinia laxa, exoproteome and expression profiles of related genes. Mol. Plant Microbe Interact. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Biggs, A.R. Early and late-season susceptibility of peach fruits to Monilinia fructicola. Plant Dis. 1988, 72, 1070. [Google Scholar] [CrossRef]
- Casals, C.; Viñas, I.; Torres, R.; Griera, C.; Usall, J. Effect of temperature and water activity on in vitro germination of Monilinia spp. J. Appl. Microbiol. 2010, 108, 47–54. [Google Scholar] [CrossRef]
- Bernat, M.; Segarra, J.; Navas-Cortés, J.A.; Casals, C.; Torres, R.; Teixidó, N.; Usall, J. Influence of temperature and humidity on the survival of Monilinia fructicola conidia on stone fruits and inert surfaces. Ann. Appl. Biol. 2018, 173, 63–70. [Google Scholar] [CrossRef]
- Van Leeuwen, G.; van Kesteren, H.A. Delineation of the three brown rot fungi of fruit crops (Monilinia spp.) on the basis of quantitative characteristics. Can. J. Bot. 1998, 76, 2042–2050. [Google Scholar] [CrossRef]
- Schumacher, J.; Pradier, J.M.; Simon, A.; Traeger, S.; Moraga, J.; Collado, I.G.; Viaud, M.; Tudzynski, B. Natural variation in the VELVET gene bcvel1 affects virulence and light-dependent differentiation in Botrytis cinerea. PLoS ONE 2012, 7, e47840. [Google Scholar] [CrossRef] [Green Version]
- Tisch, D.; Schmoll, M. Light regulation of metabolic pathways in fungi. Appl. Microbiol. Biotechnol. 2010, 85, 1259–1277. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.K.; Epton, H.A.S. Effect of light on the growth and sporulation of Botrytis cinerea. Trans. Br. Mycol. Soc. 1973, 61, 145–157. [Google Scholar] [CrossRef]
- Hevia, M.A.; Canessa, P.; Müller-Esparza, H.; Larrondo, L.F. A circadian oscillator in the fungus Botrytis cinerea regulates virulence when infecting Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 112, 8744–8749. [Google Scholar] [CrossRef] [Green Version]
- Hevia, M.A.; Canessa, P.; Larrondo, L.F. Circadian clocks and the regulation of virulence in fungi: Getting up to speed. Semin. Cell Dev. Biol. 2016, 57, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Goodspeed, D.; Chehab, E.W.; Min-Venditti, A.; Braam, J.; Covington, M.F. Arabidopsis synchronizes jasmonate-mediated defense with insect circadian behavior. Proc. Natl. Acad. Sci. USA 2012, 109, 4674–4677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Heidrich, K.; Sanchez-Villarreal, A.; Parker, J.E.; Davis, S.J. TIME FOR COFFEE represses accumulation of the MYC2 transcription factor to provide time-of-day regulation of jasmonate signaling in Arabidopsis. Plant. Cell 2012, 24, 2470–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, J.; Li, N.; Li, J.; Trail, F.; Dunlap, J.C.; Townsend, J.P. Light sensing by opsins and fungal ecology: NOP-1 modulates entry into sexual reproduction in response to environmental cues. Mol. Ecol. 2018, 27, 216–232. [Google Scholar] [CrossRef]
- Tan, K.R.; Epton, H.A.S. Ultraviolet-absorbing compounds associated with sporulation in Botrytis cinerea. Trans. Br. Mycol. Soc. 1974, 63, 157–167. [Google Scholar] [CrossRef]
- Kumagai, T. Temperature and mycochrome system in near-UV light inducible and blue light reversible photoinduction of conidiation in Alternaria tomato. Photochem. Photobiol. 1989, 50, 793–798. [Google Scholar] [CrossRef]
- Betina, V. Photoinduced conidiation in Trichoderma viride. Folia Microbiol. 1995, 40, 219–224. [Google Scholar] [CrossRef]
- Igbalajobi, O.; Yu, Z.; Fischer, R. Red-and blue-light sensing in the plant pathogen Alternaria alternata depends on phytochrome and the white-collar protein LreA. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.Z.; Honda, Y.; Sonhaji, M. Phototropism of conidial germ tubes of Botrytis cinerea and its implication in plant infection processes. Plant Dis. 1998, 82, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Röhrig, J.; Kastner, C.; Fischer, R. Light inhibits spore germination through phytochrome in Aspergillus nidulans. Curr. Genet. 2013, 59, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Marquenie, D.; Lammertyn, J.; Geeraerd, A.H.; Soontjens, C.; van Impe, J.F.; Nicola, B.M.; Michiels, C.W. Inactivation of conidia of Botrytis cinerea and Monilinia fructigena using UV-C and heat treatment. Int. J. Food Microbiol. 2002, 74, 27–35. [Google Scholar] [CrossRef]
- Bayram, Ö.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Jin, W.B.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Leroch, M.; Schumacher, J.; Zimmer, D.; Könnel, A.; Klug, K.; Leisen, T.; Scheuring, D.; Sommer, F.; Mühlhaus, T.; et al. Investigations on VELVET regulatory mutants confirm the role of host tissue acidification and secretion of proteins in the pathogenesis of Botrytis cinerea. New Phytol. 2018, 219, 1062–1074. [Google Scholar] [CrossRef] [Green Version]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Duran, R.M.; Gregersen, S.; Smith, T.D.; Bhetariya, P.J.; Cary, J.W.; Harris-Coward, P.Y.; Mattison, C.P.; Grimm, C.; Calvo, A.M. The role of Aspergillus flavus veA in the production of extracellular proteins during growth on starch substrates. Appl. Microbiol. Biotechnol. 2014, 98, 5081–5094. [Google Scholar] [CrossRef]
- De Cal, A.; Sandín-España, P.; Martinez, F.; Egüen, B.; Chien-Ming, C.; Lee, M.H.; Melgarejo, P.; Prusky, D. Role of gluconic acid and pH modulation in virulence of Monilinia fructicola on peach fruit. Postharvest Biol. Technol. 2013, 86, 418–423. [Google Scholar] [CrossRef]
- Rodríguez-Pires, S.; Melgarejo, P.; de Cal, A.; Espeso, E.A. Proteomic studies to understand the mechanisms of peach tissue degradation by Monilinia laxa. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef]
- Ballario, P.; Vittorioso, P.; Magrelli, A.; Talora, C.; Cabibbo, A.; Macino, G. White collar-1, a central regulator of blue light responses in Neurospora, is a zinc finger protein. EMBO J. 1996, 15, 1650–1657. [Google Scholar] [CrossRef]
- Cohrs, K.C.; Schumacher, J. The two cryptochrome/photolyase family proteins fulfill distinct roles in DNA photorepair and regulation of conidiation in the gray mold fungus Botrytis cinerea. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrochano, L.M. Fungal photoreceptors: Sensory molecules for fungal development and behaviour. Photochem. Photobiol. Sci. 2007, 6, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Panzer, S.; Brych, A.; Batschauer, A.; Terpitz, U. Opsin 1 and opsin 2 of the corn smut fungus Ustilago maydis are green light-driven proton pumps. Front. Microbiol. 2019, 10, 735. [Google Scholar] [CrossRef] [PubMed]
- Bieszke, J.A.; Li, L.; Borkovich, K.A. The fungal opsin gene nop-1 is negatively-regulated by a component of the blue light sensing pathway and influences conidiation-specific gene expression in Neurospora crassa. Curr. Genet. 2007, 52, 149–157. [Google Scholar] [CrossRef]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Curr. Biol. 2005, 15, 1833–1838. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Noh, B.; Vierstra, R.D.; Loros, J.; Dunlap, J.C. Genetic and molecular analysis of phytochromes from the filamentous fungus Neurospora crassa. Eukaryot. Cell 2005, 4, 2140–2152. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene/Protein ID a | GenBank Accession | Putative Protein Function b | Predicted Protein Length c | B. cinerea B05.10 d | % Identity | % Coverage | M. laxa Mlax316 e | % Identity | % Coverage |
|---|---|---|---|---|---|---|---|---|---|---|
| Putative near-UV/blue light sensors | ||||||||||
| MlCRY1 | Monilinia__090370 | BK014380 | putative deoxyribodipyrimidine photo-lyase | 679 | Bcin05g08060 | 77.80 | 90.27 | EYC80_003544 | 99.68 | 100 |
| MlCRY2 | Monilinia__028340 | BK014381 | DASH family cryptochrome protein | 649 | Bcin09g01620 | 86.59 | 95.13 | EYC80_007030 | 99.82 | 91.84 |
| Blue light sensing | ||||||||||
| MlWC1 | Monilinia__016540 | BK014382 | putative white collar-1 protein | 1094 | Bcin02g07400 | 69.52 | 93.61 | EYC80_007313 | 99.50 | 100 * |
| MlWC2 | Monilinia__092390 | BK014383 | putative white collar-2 protein | 483 | Bcin05g05530 | 72.94 | 91.36 | EYC80_003689 | 100 | 100 |
| MlVVD1 | Monilinia__019850 | BK014384 | vivid PAS VVD protein | 237 | Bcin13g01270 | 83.10 | 96.98 | EYC80_009309 | 100 | 100 |
| MlLOV3 | Monilinia__014120 | BK014393 | similar to Bclov3 | 836 | Bcin10g03870 | 78.98 | 96.50 | EYC80_002606 | 99.88 | 100 |
| MlLOV4 | Monilinia__012050 | BK014394 | regulator of G protein | 588 | Bcin02g04390 | 79.93 | 98.65 | EYC80_003150 | 100 | 100 |
| Green light sensing | ||||||||||
| MlOPS1 | Monilinia__070650 | BK014385 | putative opsin-1 protein | 310 | Bcin02g02670 | 85.26 | 99.36 | EYC80_005142 | 100 | 100 |
| MlOPS2 | Monilinia__055890 | BK014386 | putative opsin-like protein | 327 | Bcin01g04540 | 79.35 | 96.45 | EYC80_010258 | 100 | 100 |
| Red light sensing | ||||||||||
| MlPHY1 | Monilinia__010410 | BK014390 | PHY1, histidine kinase-group VIII protein | 1616 | Bcin13g04690 | 79.18 | 98.71 | EYC80_009624 | 99.94 | 100 |
| MlPHY2 | Monilinia__052440 | BK014391 | PHY2, histidine kinase-group VIII protein | 1222 | Bcin01g09230 | 75.24 | 99.51 | EYC80_005619 | 96.97 | 100 |
| MlPHY3 | Monilinia__088010 | BK014392 | PHY3, histidine kinase-group VIII protein | 1196 | Bcin06g01290 | 79.24 | 98.84 | EYC80_000954 | 98.16 | 96.46 |
| Proteins implicated in photoresponses | ||||||||||
| MlVEL1 | Monilinia__005480 | BK014387 | velvet complex subunit 1 | 589 | Bcin15g03390 | 75.17 | 99.48 | EYC80_008048 | 99.66 | 95.77 |
| MlVEL2 | Monilinia__093380 | BK014388 | velvet complex subunit 2 | 446 | Bcin01g02730 | 89.02 | 93.79 | EYC80_005890 | 100 | 100 |
| MlVEL3 | Monilinia__072950 | BK014389 | velvet 3 | 492 | Bcin03g06410 | 78.64 | 98.00 | EYC80_000111 | 100 | 100 |
| MlVEL4 | Monilinia__100000 | MW349131 | velvet 4 | 375 | Bcin07g05880 | 92.70 | 95.41 | EYC80_003351 | 98.70 | 95.66 |
| Proteins implicated in the circadian clock | ||||||||||
| MlFRQ1 | Monilinia__081690 | BK014395 | putative frequency clock protein | 968 | Bcin02g08360 | 71.25 | 98.55 | EYC80_007367 | 97.93 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Pires, S.; Espeso, E.A.; Rasiukevičiūtė, N.; Melgarejo, P.; De Cal, A. Light-Photoreceptors and Proteins Related to Monilinia laxa Photoresponses. J. Fungi 2021, 7, 32. https://doi.org/10.3390/jof7010032
Rodríguez-Pires S, Espeso EA, Rasiukevičiūtė N, Melgarejo P, De Cal A. Light-Photoreceptors and Proteins Related to Monilinia laxa Photoresponses. Journal of Fungi. 2021; 7(1):32. https://doi.org/10.3390/jof7010032
Chicago/Turabian StyleRodríguez-Pires, Silvia, Eduardo A. Espeso, Neringa Rasiukevičiūtė, Paloma Melgarejo, and Antonieta De Cal. 2021. "Light-Photoreceptors and Proteins Related to Monilinia laxa Photoresponses" Journal of Fungi 7, no. 1: 32. https://doi.org/10.3390/jof7010032
APA StyleRodríguez-Pires, S., Espeso, E. A., Rasiukevičiūtė, N., Melgarejo, P., & De Cal, A. (2021). Light-Photoreceptors and Proteins Related to Monilinia laxa Photoresponses. Journal of Fungi, 7(1), 32. https://doi.org/10.3390/jof7010032