Candida parapsilosis Colony Morphotype Forecasts Biofilm Formation of Clinical Isolates



 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Routine Diagnostic and Strain Maintenance
2.2. Biofilm Quantitation
2.3. Antifungal Drug Susceptibility Testing
2.4. Morphotype Development and Agar Invasion
2.5. Statistical Analyses
3. Results
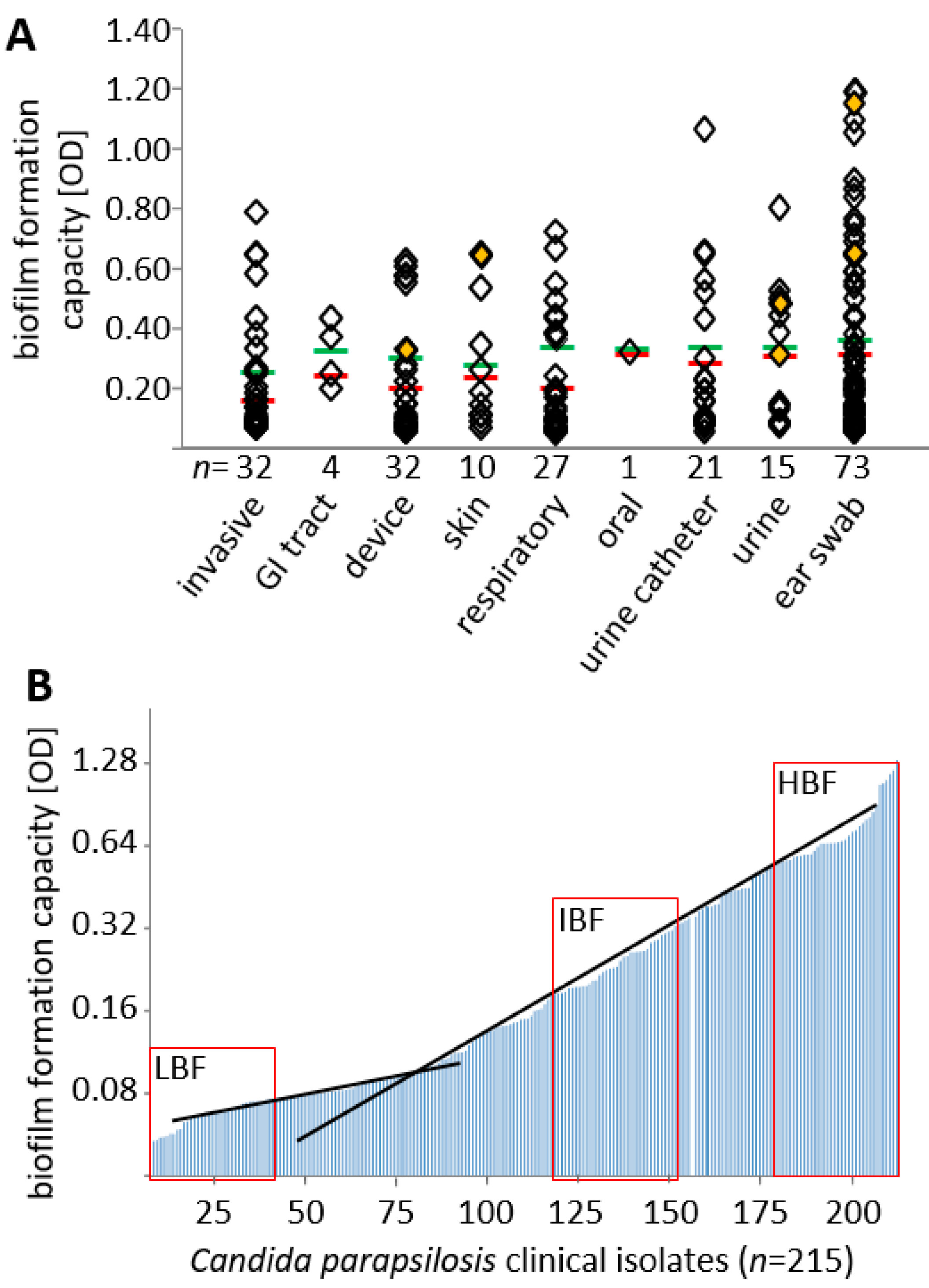
3.1. Biofilm Formation Capacity Is Independent of Body Site of Isolation
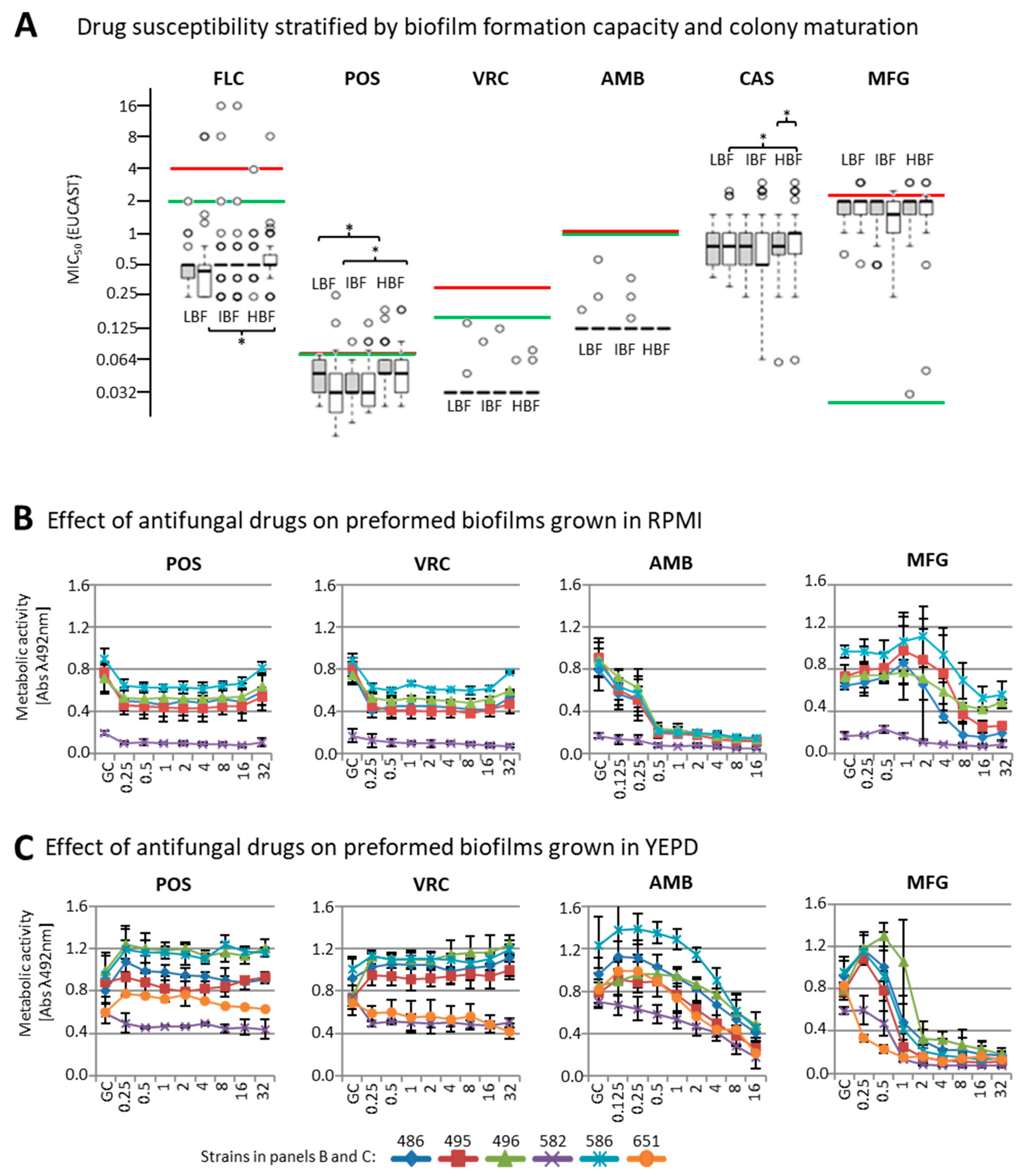
3.2. Effect of Biofilm Formation on Antifungal Drug Susceptibility
3.3. Susceptibility of Biofilms to Antifungal Drugs
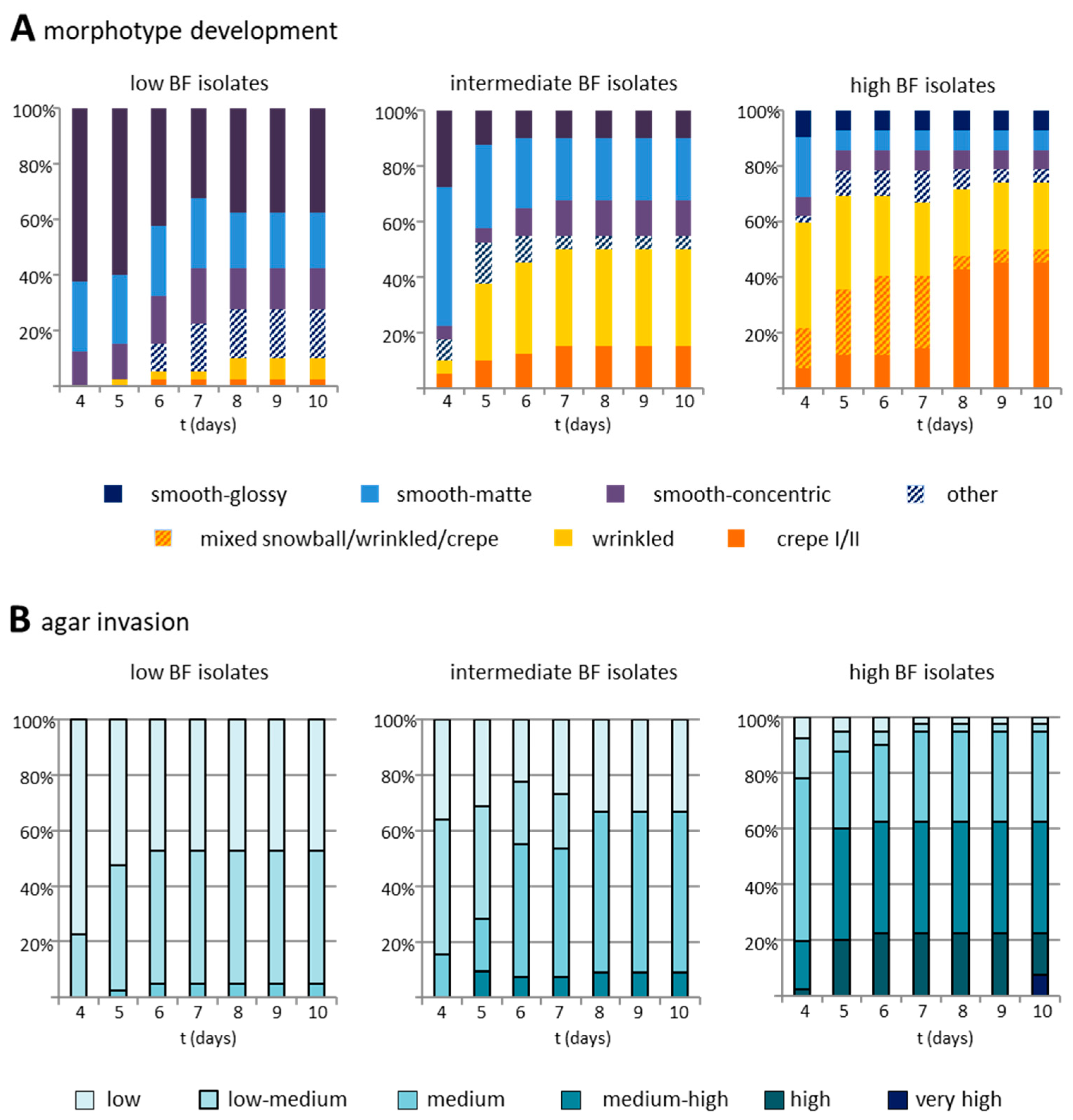
3.4. Biofilm Formation Capacity on Polystyrol Correlates with Colony Morphotype and Agar Invasiveness
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weems, J.J., Jr. Candida parapsilosis: Epidemiology, pathogenicity, clinical manifestations, and antimicrobial susceptibility. Clin. Infect. Dis. 1992, 14, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Krcmery, V.; Barnes, A.J. Non-albicans Candida spp. causing fungaemia: Pathogenicity and antifungal resistance. J. Hosp. Infect. 2002, 50, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J. Role of sentinel surveillance of candidemia: Trends in species distribution and antifungal susceptibility. J. Clin. Microbiol. 2002, 40, 3551–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pammi, M.; Holland, L.; Butler, G.; Gacser, A.; Bliss, J.M. Candida parapsilosis is a significant neonatal pathogen: A systematic review and meta-analysis. Pediatr. Infect. Dis. J. 2013, 32, e206–e216. [Google Scholar] [CrossRef] [Green Version]
- Trofa, D.; Gacser, A.; Nosanchuk, J.D. Candida parapsilosis, an emerging fungal pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, E.; Hammarstrom, H.; Ataollahy, N.; Kondori, N. Species distribution and antifungal drug susceptibilities of yeasts isolated from the blood samples of patients with candidemia. Sci. Rep. 2019, 9, 3838. [Google Scholar] [CrossRef] [Green Version]
- Toth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D.; Turner, S.A.; Butler, G.; Vagvolgyi, C.; Gacser, A. Candida parapsilosis: From Genes to the Bedside. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [Green Version]
- Lupetti, A.; Tavanti, A.; Davini, P.; Ghelardi, E.; Corsini, V.; Merusi, I.; Boldrini, A.; Campa, M.; Senesi, S. Horizontal transmission of Candida parapsilosis candidemia in a neonatal intensive care unit. J. Clin. Microbiol. 2002, 40, 2363–2369. [Google Scholar] [CrossRef] [Green Version]
- Gacser, A.; Trofa, D.; Schafer, W.; Nosanchuk, J.D. Targeted gene deletion in Candida parapsilosis demonstrates the role of secreted lipase in virulence. J. Clin. Investig. 2007, 117, 3049–3058. [Google Scholar] [CrossRef] [Green Version]
- Linden, J.R.; De Paepe, M.E.; Laforce-Nesbitt, S.S.; Bliss, J.M. Galectin-3 plays an important role in protection against disseminated candidiasis. Med. Mycol. 2013, 51, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, D.M.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Comparison of biofilms formed by Candida albicans and Candida parapsilosis on bioprosthetic surfaces. Infect. Immun. 2002, 70, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neji, S.; Hadrich, I.; Trabelsi, H.; Abbes, S.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Ayadi, A. Virulence factors, antifungal susceptibility and molecular mechanisms of azole resistance among Candida parapsilosis complex isolates recovered from clinical specimens. J. Biomed. Sci. 2017, 24, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattif, A.A.; Mukherjee, P.K.; Chandra, J.; Swindell, K.; Lockhart, S.R.; Diekema, D.J.; Pfaller, M.A.; Ghannoum, M.A. Characterization of biofilms formed by Candida parapsilosis, C. metapsilosis, and C. orthopsilosis. Int. J. Med. Microbiol. 2010, 300, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Pannanusorn, S.; Fernandez, V.; Romling, U. Prevalence of biofilm formation in clinical isolates of Candida species causing bloodstream infection. Mycoses 2013, 56, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Kozik, A.; Karkowska-Kuleta, J.; Zajac, D.; Bochenska, O.; Kedracka-Krok, S.; Jankowska, U.; Rapala-Kozik, M. Fibronectin-, vitronectin- and laminin-binding proteins at the cell walls of Candida parapsilosis and Candida tropicalis pathogenic yeasts. BMC Microbiol. 2015, 15, 197. [Google Scholar] [CrossRef] [Green Version]
- Butler, G.; Rasmussen, M.D.; Lin, M.F.; Santos, M.A.; Sakthikumar, S.; Munro, C.A.; Rheinbay, E.; Grabherr, M.; Forche, A.; Reedy, J.L.; et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature 2009, 459, 657–662. [Google Scholar] [CrossRef] [Green Version]
- Pryszcz, L.P.; Nemeth, T.; Gacser, A.; Gabaldon, T. Unexpected genomic variability in clinical and environmental strains of the pathogenic yeast Candida parapsilosis. Genome Biol. Evol. 2013, 5, 2382–2392. [Google Scholar] [CrossRef] [Green Version]
- Neale, M.N.; Glass, K.A.; Longley, S.J.; Kim, D.J.; Laforce-Nesbitt, S.S.; Wortzel, J.D.; Shaw, S.K.; Bliss, J.M. Role of the Inducible Adhesin CpAls7 in Binding of Candida parapsilosis to the Extracellular Matrix under Fluid Shear. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, T.; Ding, C.; Guida, A.; d′Enfert, C.; Higgins, D.G.; Butler, G. Correlation between biofilm formation and the hypoxic response in Candida parapsilosis. Eukaryot. Cell 2009, 8, 550–559. [Google Scholar] [CrossRef] [Green Version]
- Katragkou, A.; Chatzimoschou, A.; Simitsopoulou, M.; Dalakiouridou, M.; Diza-Mataftsi, E.; Tsantali, C.; Roilides, E. Differential activities of newer antifungal agents against Candida albicans and Candida parapsilosis biofilms. Antimicrob. Agents Chemother. 2008, 52, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soldini, S.; Posteraro, B.; Vella, A.; De Carolis, E.; Borghi, E.; Falleni, M.; Losito, A.R.; Maiuro, G.; Trecarichi, E.M.; Sanguinetti, M.; et al. Microbiologic and clinical characteristics of biofilm-forming Candida parapsilosis isolates associated with fungaemia and their impact on mortality. Clin. Microbiol. Infect. 2018, 24, 771–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soll, D.R. The role of phenotypic switching in the basic biology and pathogenesis of Candida albicans. J. Oral Microbiol. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Molero, E.; Willis, J.; Dudakova, A.; Gacser, A.; Weig, M.; Groß, U.; Gabaldón, T.; Bader, O. Phenotypic variability in a coinfection with three independent C. parapsilosis lineages. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Laffey, S.F.; Butler, G. Phenotype switching affects biofilm formation by Candida parapsilosis. Microbiology 2005, 151, 1073–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lott, T.J.; Kuykendall, R.J.; Welbel, S.F.; Pramanik, A.; Lasker, B.A. Genomic heterogeneity in the yeast Candida parapsilosis. Curr. Genet. 1993, 23, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Enger, L.; Joly, S.; Pujol, C.; Simonson, P.; Pfaller, M.; Soll, D.R. Cloning and characterization of a complex DNA fingerprinting probe for Candida parapsilosis. J. Clin. Microbiol. 2001, 39, 658–669. [Google Scholar] [CrossRef] [Green Version]
- Nosek, J.; Holesova, Z.; Kosa, P.; Gacser, A.; Tomaska, L. Biology and genetics of the pathogenic yeast Candida parapsilosis. Curr. Genet. 2009, 55, 497–509. [Google Scholar] [CrossRef]
- Bernhard, M.; Weig, M.; Zautner, A.E.; Gross, U.; Bader, O. Yeast on-target lysis (YOTL), a procedure for making auxiliary mass spectrum data sets for clinical routine identification of yeasts. J. Clin. Microbiol. 2014, 52, 4163–4167. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Molero, E.; de Boer, A.D.; Dekker, H.L.; Moreno-Martínez, A.; Kraneveld, E.A.; Ichsan Chauhan, N.; Weig, M.; de Soet, J.J.; de Koster, C.G.; Bader, O.; et al. Proteomic analysis of hyperadhesive Candida glabrata clinical isolates reveals a core wall proteome and differential incorporation of adhesins. FEMS Yeast Res. 2015, 15, fov098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EUCAST. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_7_3_1_Yeast_testing__definitive.pdf (accessed on 16 April 2019).
- Grossman, N.T.; Pham, C.D.; Cleveland, A.A.; Lockhart, S.R. Molecular mechanisms of fluconazole resistance in Candida parapsilosis isolates from a U.S. surveillance system. Antimicrob. Agents Chemother. 2015, 59, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Vande Walle, K.; Wickes, B.L.; Lopez-Ribot, J.L. Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arastehfar, A.; Daneshnia, F.; Hilmioglu-Polat, S.; Fang, W.; Yasar, M.; Polat, F.; Yesim Metin, D.; Rigole, P.; Coenye, T.; Ilkit, M.; et al. First report of candidemia clonal outbreak caused by emerging fluconazole-resistant Candida parapsilosis isolates harboring Y132F and/or Y132F+K143R in Turkey. Antimicrob. Agents Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.; Fuchs, B.B.; Pinhati, H.M.; Siqueira, R.A.; Hagen, F.; Meis, J.F.; Mylonakis, E.; Colombo, A.L. Candida parapsilosis Resistance to Fluconazole: Molecular Mechanisms and In Vivo Impact in Infected Galleria mellonella Larvae. Antimicrob. Agents Chemother. 2015, 59, 6581–6587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magobo, R.E.; Lockhart, S.R.; Govender, N.P. Fluconazole-resistant Candida parapsilosis strains with a Y132F substitution in the ERG11 gene causing invasive infections in a neonatal unit, South Africa. Mycoses 2020, 63, 471–477. [Google Scholar] [CrossRef]
- Van Asbeck, E.C.; Clemons, K.V.; Stevens, D.A. Candida parapsilosis: A review of its epidemiology, pathogenesis, clinical aspects, typing and antimicrobial susceptibility. Crit. Rev. Microbiol. 2009, 35, 283–309. [Google Scholar] [CrossRef]
- Imbert, C.; Rammaert, B. What Could Be the Role of Antifungal Lock-Solutions? From Bench to Bedside. Pathogens 2018, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Borges, K.R.A.; Pimentel, I.V.; Lucena, L.; Silva, M.; Monteiro, S.G.; Monteiro, C.A.; Nascimento, M.; Bezerra, G.F.B. Adhesion and biofilm formation of Candida parapsilosis isolated from vaginal secretions to copper intrauterine devices. Rev. Inst. Med. Trop. Sao Paulo 2018, 60, e59. [Google Scholar] [CrossRef]
- Tan, Y.; Leonhard, M.; Ma, S.; Schneider-Stickler, B. Influence of culture conditions for clinically isolated non-albicans Candida biofilm formation. J. Microbiol. Methods 2016, 130, 123–128. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomaz, D.Y.; de Almeida, J.N., Jr.; Lima, G.M.E.; Nunes, M.O.; Camargo, C.H.; Grenfell, R.C.; Benard, G.; Del Negro, G.M.B. An Azole-Resistant Candida parapsilosis Outbreak: Clonal Persistence in the Intensive Care Unit of a Brazilian Teaching Hospital. Front. Microbiol. 2018, 9, 2997. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.S.; Bizerra, F.C.; Freymuller, E.; Arthington-Skaggs, B.A.; Colombo, A.L. Biofilm production and evaluation of antifungal susceptibility amongst clinical Candida spp. isolates, including strains of the Candida parapsilosis complex. Med. Mycol. 2011, 49, 253–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinel-Ingroff, A.; Barchiesi, F.; Cuenca-Estrella, M.; Pfaller, M.A.; Rinaldi, M.; Rodriguez-Tudela, J.L.; Verweij, P.E. International and multicenter comparison of EUCAST and CLSI M27-A2 broth microdilution methods for testing susceptibilities of Candida spp. to fluconazole, itraconazole, posaconazole, and voriconazole. J. Clin. Microbiol. 2005, 43, 3884–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modiri, M.; Hashemi, S.J.; Ghazvin, I.R.; Khodavaisy, S.; Ahmadi, A.; Ghaffari, M.; Rezaie, S. Antifungal susceptibility pattern and biofilm-related genes expression in planktonic and biofilm cells of Candida parapsilosis species complex. Curr. Med. Mycol. 2019, 5, 35–42. [Google Scholar] [CrossRef]
- Khodavaisy, S.; Badali, H.; Meis, J.F.; Modiri, M.; Mahmoudi, S.; Abtahi, H.; Salehi, M.; Dehghan Manshadi, S.A.; Aala, F.; Agha Kuchak Afshari, S.; et al. Comparative in vitro activities of seven antifungal drugs against clinical isolates of Candida parapsilosis complex. J. Mycol. Med. 2020, 30, 100968. [Google Scholar] [CrossRef]
- Rodriguez-Tudela, J.L.; Gomez-Lopez, A.; Arendrup, M.C.; Garcia-Effron, G.; Perlin, D.S.; Lass-Florl, C.; Cuenca-Estrella, M. Comparison of caspofungin MICs by means of EUCAST method EDef 7.1 using two different concentrations of glucose. Antimicrob. Agents Chemother. 2010, 54, 3056–3057. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.; Leite-Andrade, M.C.; Brandao, I.S.; Alves, A.; Buonafina, M.D.S.; Nunes, M.; Araujo-Neto, L.N.; Freitas, M.A.; Brayner, F.A.; Alves, L.C.; et al. Anti-biofilm effect by the combined action of fluconazole and acetylsalicylic acid against species of Candida parapsilosis complex. Infect. Genet. Evol. 2020, 84, 104378. [Google Scholar] [CrossRef]
- Pannanusorn, S.; Ramirez-Zavala, B.; Lunsdorf, H.; Agerberth, B.; Morschhauser, J.; Romling, U. Characterization of biofilm formation and the role of BCR1 in clinical isolates of Candida parapsilosis. Eukaryot. Cell 2014, 13, 438–451. [Google Scholar] [CrossRef] [Green Version]
- De Bernardis, F.; Mondello, F.; San Millan, R.; Ponton, J.; Cassone, A. Biotyping and virulence properties of skin isolates of Candida parapsilosis. J. Clin. Microbiol. 1999, 37, 3481–3486. [Google Scholar] [CrossRef] [Green Version]
- Rupp, S.; Summers, E.; Lo, H.J.; Madhani, H.; Fink, G. MAP kinase and cAMP filamentation signaling pathways converge on the unusually large promoter of the yeast FLO11 gene. EMBO J. 1999, 18, 1257–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, T.; Toth, Z.; Toth, R.; Vagvolgyi, C.; Gacser, A. Iron Metabolism, Pseudohypha Production, and Biofilm Formation through a Multicopper Oxidase in the Human-Pathogenic Fungus Candida parapsilosis. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Cassone, A.; De Bernardis, F.; Pontieri, E.; Carruba, G.; Girmenia, C.; Martino, P.; Fernandez-Rodriguez, M.; Quindos, G.; Ponton, J. Biotype diversity of Candida parapsilosis and its relationship to the clinical source and experimental pathogenicity. J. Infect. Dis. 1995, 171, 967–975. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphology a | n | Agar Invasion a | |||||
|---|---|---|---|---|---|---|---|
| Low | Low-Medium | Medium | Medium-High | High | Very High | ||
| smooth | 59 | 34% | 27% | 5% | 32% | 2% | 0% |
| wrinkled | 22 | 5% | 9% | 23% | 50% | 9% | 5% |
| mixed/infrequent | 16 | 0% | 6% | 6% | 63% | 19% | 6% |
| crepe I and II | 25 | 4% | 4% | 0% | 36% | 28% | 28% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Molero, E.; De-la-Pinta, I.; Fernández-Pereira, J.; Groß, U.; Weig, M.; Quindós, G.; de Groot, P.W.J.; Bader, O. Candida parapsilosis Colony Morphotype Forecasts Biofilm Formation of Clinical Isolates. J. Fungi 2021, 7, 33. https://doi.org/10.3390/jof7010033
Gómez-Molero E, De-la-Pinta I, Fernández-Pereira J, Groß U, Weig M, Quindós G, de Groot PWJ, Bader O. Candida parapsilosis Colony Morphotype Forecasts Biofilm Formation of Clinical Isolates. Journal of Fungi. 2021; 7(1):33. https://doi.org/10.3390/jof7010033
Chicago/Turabian StyleGómez-Molero, Emilia, Iker De-la-Pinta, Jordan Fernández-Pereira, Uwe Groß, Michael Weig, Guillermo Quindós, Piet W. J. de Groot, and Oliver Bader. 2021. "Candida parapsilosis Colony Morphotype Forecasts Biofilm Formation of Clinical Isolates" Journal of Fungi 7, no. 1: 33. https://doi.org/10.3390/jof7010033
APA StyleGómez-Molero, E., De-la-Pinta, I., Fernández-Pereira, J., Groß, U., Weig, M., Quindós, G., de Groot, P. W. J., & Bader, O. (2021). Candida parapsilosis Colony Morphotype Forecasts Biofilm Formation of Clinical Isolates. Journal of Fungi, 7(1), 33. https://doi.org/10.3390/jof7010033