
Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives


Abstract
:1. Introduction
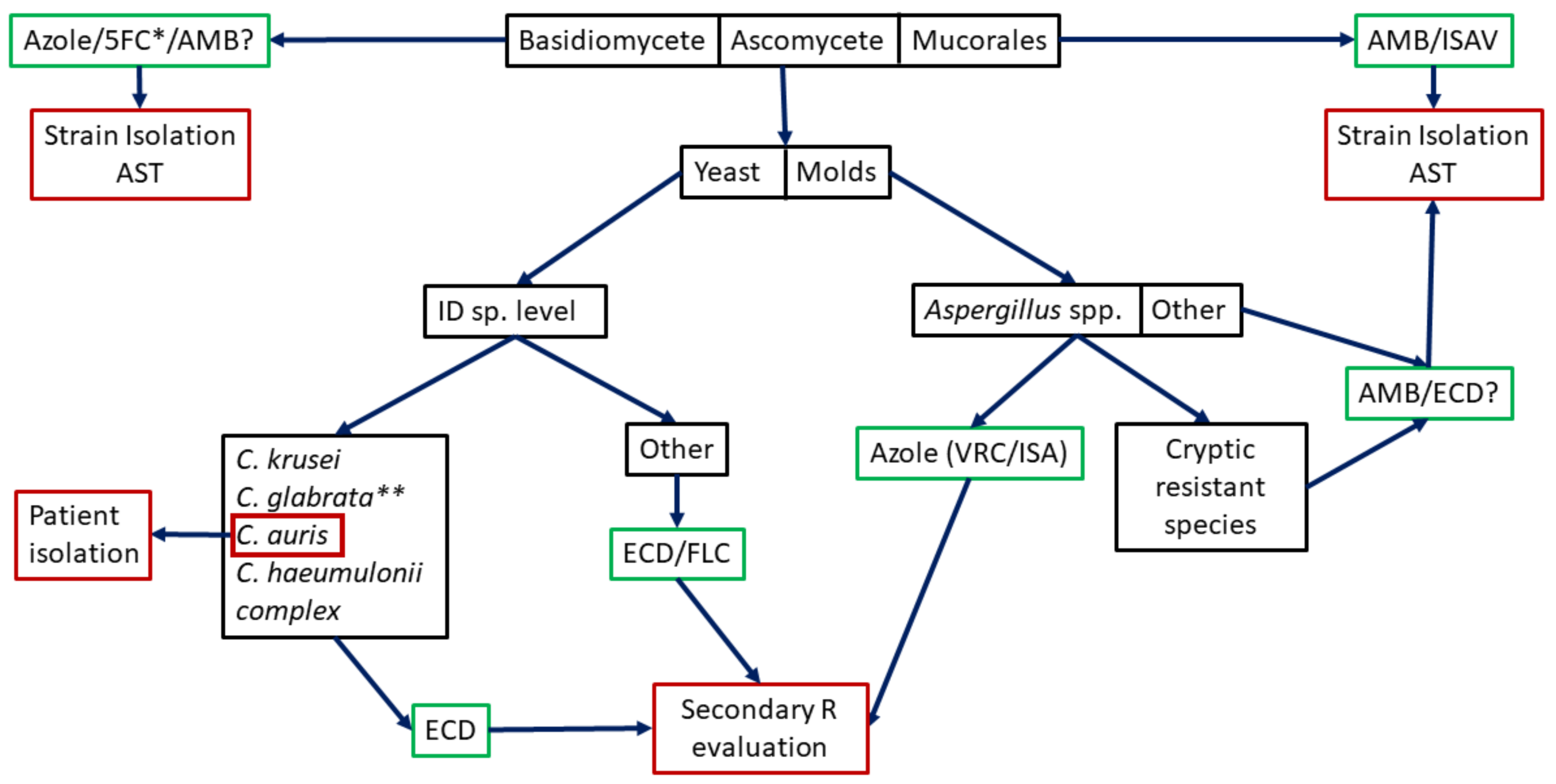
2. Intrinsic Resistance Detection
2.1. Intrinsic Resistance Detection by Commercially Available Molecular Taxonomy-Based Method
2.2. Intrinsic Resistance Detection by in-House Molecular-Based Method
2.3. Is Species Identification Enough as Surrogate Marker of Intrinsic Resistance or Should We Go Further?
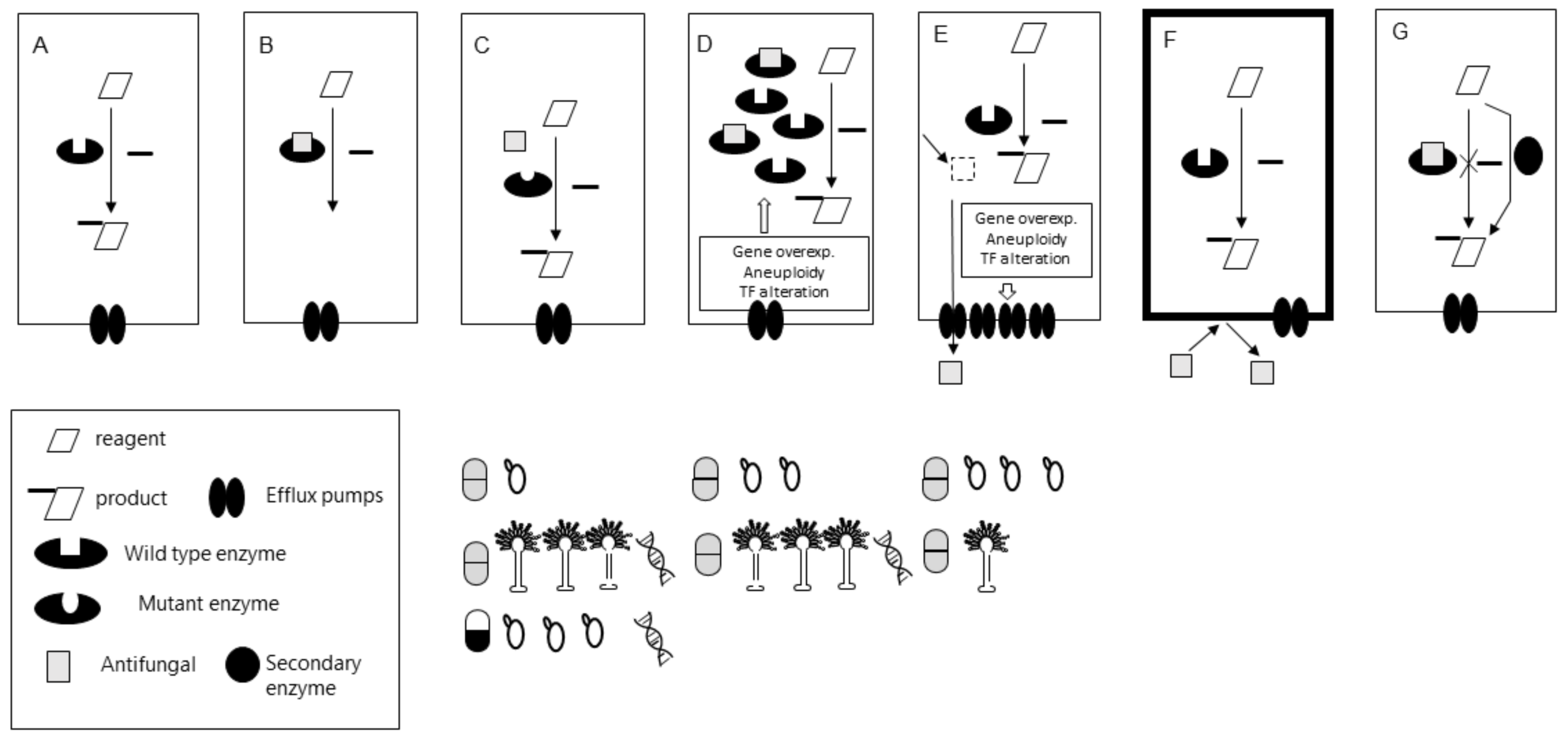
3. Secondary Resistance Detection
3.1. The Bottlenecks of Secondary Resistance Molecular Detection
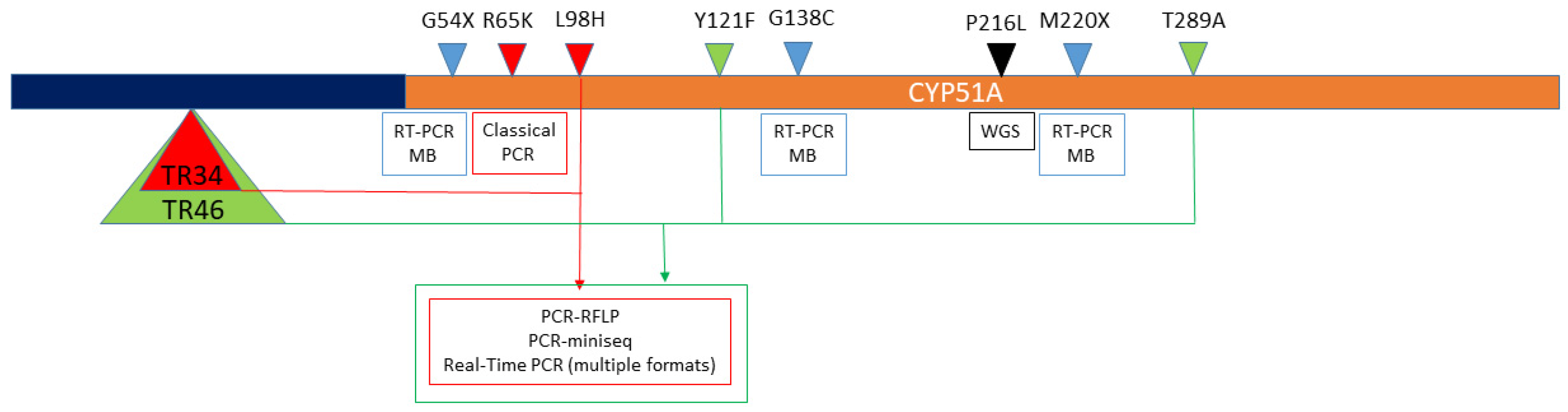
3.2. Available Molecular Tools. Which Secondary Mechanisms Are We Able to Detect?
3.2.1. Triazole Secondary Resistance in Aspergillus spp.
3.2.2. Azole Resistance in Candida spp.
3.2.3. Echinocandin Resistance in Candida spp.
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berkow, E.L.; Lockhart, S.R.; Ostrosky-Zeichner, L. Antifungal susceptibility testing: Current approaches. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef]
- Ganesan, A.; Wells, J.; Shaikh, F.; Peterson, P.; Bradley, W.; Carson, M.L.; Petfield, J.L.; Klassen-Fischer, M.; Akers, K.S.; Downing, K.; et al. Molecular Detection of Filamentous Fungi in Formalin-Fixed Paraffin-Embedded Specimens in Invasive Fungal Wound Infections Is Feasible with High Specificity. J. Clin. Microbiol. 2019, 58. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Chen, S.C.; Kauffman, C.A.; Steinbach, W.J.; Baddley, J.W.; Verweij, P.E.E.; Clancy, C.J.; Wingard, J.R.; Lockhart, S.R.; Groll, A.H.; et al. Revision and Update of the Consensus Definitions of Invasive Fungal Disease from the European Organization for Research and Treatment of Cancer and the Mycoses Study Group Education and Research Consortium. Clin. Infect. Dis. 2020, 71, 1367–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickes, B.L.; Wiederhold, N.P. Molecular diagnostics in medical mycology. Nat. Commun. 2018, 9, 5135. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Strandards Institute CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; CLSI Standard M38; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard-Third Edition M27-A3; CLSI (Clinical and Laboratory Standard Institute): Wayne, PA, USA, 2008. [Google Scholar]
- EUCAST. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Conidia Forming Moulds; EUCAST Definitive Document E.DEF 9.3.2.; EUCAST. 2020. Available online: https://www.aspergillus.org.uk/wp-content/uploads/2016/03/EUCAST_E_Def_9_3_Mould_testing_definitive_0.pdf (accessed on 9 March 2021).
- EUCAST. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts; EUCAST Document E.DEF 7.3.2. Clin. Microbiol. Infect. 2008, 14, 398–405. [Google Scholar] [CrossRef] [PubMed]
- White, T.C.; Marr, K.A.; Bowden, R.A. Clinical, Cellular, and Molecular Factors That Contribute to Antifungal Drug Resistance. Clin. Microbiol. Rev. 1998, 11, 382–402. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Lass-Flörl, C.; Garcia-Rubio, R.; Daneshnia, F.; Ilkit, M.; Boekhout, T.; Gabaldon, T.; Perlin, D.S. The quiet and underappreciated rise of drug-resistant invasive fungal pathogens. J. Fungi 2020, 6, 138. [Google Scholar] [CrossRef]
- Perlin, D.S. Echinocandin Resistance in Candida. Clin. Infect. Dis. 2015, 61, S612–S617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanglard, D.; Coste, A.; Ferrari, S. Antifungal drug resistance mechanisms in fungal pathogens from the perspective of transcriptional gene regulation. FEMS Yeast Res. 2009, 9, 1029–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardelli, F.; Macedo, D.; Dudiuk, C.; Cabeza, M.S.; Gamarra, S.; Garcia-Effron, G. Aspergillus fumigatus Intrinsic Fluconazole Resistance Is Due to the Naturally Occurring T301I Substitution in Cyp51Ap. Antimicrob. Agents Chemother. 2016, 60, 5420–5426. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.C.; Kelly, D.E.; Schunck, W.-H.; Shyadehi, A.Z.; Akhtar, M.; Lowe, D.J.; Baldwin, B.C.; Kelly, S.L. The Mutation T315A in Candida albicans Sterol 14α-Demethylase Causes Reduced Enzyme Activity and Fluconazole Resistance through Reduced Affinity. J. Biol. Chem. 1997, 272, 5682–5688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; M100Ed30; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- EUCAST. EUCAST Expert Rules Version 2 2020. Available online: https://www.eucast.org/expert_rules_and_intrinsic_resistance/ (accessed on 1 March 2020).
- Clinical and Laboratory Strandards Institute CLSI. Performance Standards for Antifungal Susceptibility Testing of Yeasts, 2nd ed.; M60; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- De Carolis, E.; Posteraro, B.; Lass-Flörl, C.; Vella, A.; Florio, A.R.; Torelli, R.; Girmenia, C.; Colozza, C.; Tortorano, A.M.; Sanguinetti, M.; et al. Species identification of Aspergillus, Fusarium and Mucorales with direct surface analysis by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Microbiol. Infect. 2012, 18, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iriart, X.; Lavergne, R.-A.; Fillaux, J.; Valentin, A.; Magnaval, J.-F.; Berry, A.; Cassaing, S. Routine Identification of Medical Fungi by the New Vitek MS Matrix-Assisted Laser Desorption Ionization–Time of Flight System with a New Time-Effective Strategy. J. Clin. Microbiol. 2012, 50, 2107–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bille, E.; Dauphin, B.; Leto, J.; Bougnoux, M.-E.; Beretti, J.-L.; Lotz, A.; Suarez, S.; Meyer, J.; Join-Lambert, O.; Descamps, P.; et al. MALDI-TOF MS Andromas strategy for the routine identification of bacteria, mycobacteria, yeasts, Aspergillus spp. and positive blood cultures. Clin. Microbiol. Infect. 2012, 18, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Wilkendorf, L.S.; Bowles, E.; Buil, J.B.; Van Der Lee, H.A.L.; Posteraro, B.; Sanguinetti, M.; Verweij, P.E. Update on Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry Identification of Filamentous Fungi. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Gamarra, S.; Chaves, M.; Cabeza, M.; Macedo, D.; Leonardelli, F.; Franco, D.; Boleas, M.; Garcia-Effron, G. Mucormycosis outbreak due to Rhizopus microsporus after arthroscopic anterior cruciate ligament reconstruction surgery evaluated by RAPD and MALDI-TOF Mass spectrometry. J. Med Mycol. 2018, 28, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.G.; Romero, C.; Dard, C.; Garnaud, C.; Cognet, O.; Girard, T.; Rasamoelina, T.; Cornet, M.; Maubon, D. Evaluation of ID Fungi Plates Medium for Identification of Molds by MALDI Biotyper. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Guarro, J.; Gené, J.; Stchigel, A.M. Developments in Fungal Taxonomy. Clin. Microbiol. Rev. 1999, 12, 454–500. [Google Scholar] [CrossRef] [Green Version]
- Gregory, T.R. DNA barcodes an adjunct to linnaean taxonomy. Nature 2005, 434, 1067. [Google Scholar] [CrossRef] [Green Version]
- Irinyi, L.; Lackner, M.; De Hoog, G.S.; Meyer, W. DNA barcoding of fungi causing infections in humans and animals. Fungal Biol. 2016, 120, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Schindel, D.E.; Miller, S.E. Benefits of DNA barcoding. Nature 2005, 435, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Will, K.W.; Rubinoff, D. Myth of the molecule: DNA barcodes for species cannot replace morphology for identification and classification. Cladistics 2004, 20, 47–55. [Google Scholar] [CrossRef]
- Marshall, E. Will DNA bar codes breathe life into classification? Science 2005, 307, 1037. [Google Scholar] [CrossRef] [PubMed]
- Ebach, M.C.; Holdrege, C. DNA barcoding is no substitute for taxonomy. Nat. Cell Biol. 2005, 434, 697. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.P.; Paulay, G. DNA Barcoding: Error Rates Based on Comprehensive Sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in Silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.L.; Leaw, S.N.; Chen, J.-H.; Chang, H.C.; Chang, T.C. Rapid Identification of Yeasts Commonly Found in Positive Blood Cultures by Amplification of the Internal Transcribed Spacer Regions 1 and 2. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 693–696. [Google Scholar] [CrossRef]
- Borman, A.M.; Petch, R.; Linton, C.J.; Palmer, M.D.; Bridge, P.D.; Johnson, E.M. Candida nivariensis, an Emerging Pathogenic Fungus with Multidrug Resistance to Antifungal Agents. J. Clin. Microbiol. 2008, 46, 933–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-López, S.; Dudiuk, C.; Vivot, W.; Szusz, W.; Córdoba, S.B.; Garcia-Effron, G. Phenotypic and Molecular Evaluation of Echinocandin Susceptibility of Candida glabrata, Candida bracarensis, and Candida nivariensis Strains Isolated during 30 Years in Argentina. Antimicrob. Agents Chemother. 2017, 61, e00170-17. [Google Scholar] [CrossRef] [Green Version]
- Cartier, N.; Chesnay, A.; N’Diaye, D.; Thorey, C.; Ferreira, M.; Haillot, O.; Bailly, É.; Desoubeaux, G. Candida nivariensis: Identification strategy in mycological laboratories. J. Med. Mycol. 2020, 30, 101042. [Google Scholar] [CrossRef]
- Dudiuk, C.; Morales-López, S.E.; Podesta, V.; Macedo, D.; Leonardelli, F.; Vitale, R.G.; Tosello, M.E.; Cabeza, M.S.; Biasoli, M.; Gamarra, S.; et al. Multiplex PCR designed to differentiate species within the Candida glabrata complex. Rev. Iberoam. Micol. 2017, 34, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.; Pawłowska, J.; Alastruey-Izquierdo, A.; Wrzosek, M.; Rodriguez-Tudela, J.; Dolatabadi, S.; Chakrabarti, A.; De Hoog, G. DNA barcoding in Mucorales: An inventory of biodiversity. Pers. Mol. Phylogeny Evol. Fungi 2013, 30, 11–47. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic Species Recognition and Species Concepts in Fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Tudela, J.L.; Diaz-Guerra, T.M.; Mellado, E.; Cano, V.; Tapia, C.; Perkins, A.; Gomez-Lopez, A.; Rodero, L.; Cuenca-Estrella, M. Susceptibility Patterns and Molecular Identification of Trichosporon Species. Antimicrob. Agents Chemother. 2005, 49, 4026–4034. [Google Scholar] [CrossRef] [Green Version]
- Roe, A.D.; Rice, A.V.; Bromilow, S.E.; Cooke, J.E.K.; Sperling, F.A.H. Multilocus species identification and fungal DNA barcoding: Insights from blue stain fungal symbionts of the mountain pine beetle. Mol. Ecol. Resour. 2010, 10, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [Green Version]
- Balajee, S.A.; Gribskov, J.L.; Hanley, E.; Nickle, D.; Marr, K.A. Aspergillus lentulus sp. nov., a new sibling species of A. fumigatus. Eukaryot. Cell 2005, 4, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Gautier, M.; Normand, A.-C.; Ranque, S. Previously unknown species of Aspergillus. Clin. Microbiol. Infect. 2016, 22, 662–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubka, V.; Kolarik, M. β-tubulin paralogue tubC is frequently misidentified as the benA gene in Aspergillus section Nigri taxonomy: Primer specificity testing and taxonomic consequences. Pers. Mol. Phylogeny Evol. Fungi 2012, 29, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Peterson, S.W. Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 2008, 100, 205–226. [Google Scholar] [CrossRef]
- Samson, R.; Visagie, C.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.; Perrone, G.; Seifert, K.; Susca, A.; Tanney, J.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Van Diepeningen, A.D.; Feng, P.; Ahmed, S.; Sudhadham, M.; Bunyaratavej, S.; De Hoog, G.S. Spectrum ofFusariuminfections in tropical dermatology evidenced by multilocus sequencing typing diagnostics. Mycoses 2014, 58, 48–57. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Nucleic Acid Based Tests. FDA. Available online: https://www.fda.gov/medical-devices/vitro-diagnostics/nucleic-acid-based-tests (accessed on 28 December 2020).
- Simor, A.E.; Porter, V.; Mubareka, S.; Chouinard, M.; Katz, K.; Vermeiren, C.; Fattouh, R.; Matukas, L.M.; Tadros, M.; Mazzulli, T.; et al. Rapid Identification of Candida Species from Positive Blood Cultures by Use of the Film Array Blood Culture Identification Panel. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacharioudakis, I.M.; Zervou, F.N.; Mylonakis, E. T2 Magnetic Resonance Assay: Overview of Available Data and Clinical Implications. J. Fungi 2018, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Biomerieux Enhanced BIOFIRE® Blood Culture Identification 2 (BCID2). Available online: https://www.rapidmicrobiology.com/news/enhanced-biofire-bcid2-panel-submitted-for-fda-clearance (accessed on 26 December 2020).
- Clancy, C.J.; Nguyen, M.H. Finding the “Missing 50%” of Invasive Candidiasis: How Nonculture Diagnostics Will Improve Understanding of Disease Spectrum and Transform Patient Care. Clin. Infect. Dis. 2013, 56, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Kasper, D.C.; Altiok, I.; Mechtler, T.P.; Böhm, J.; Straub, J.; Langgartner, M.; Pollak, A.; Herkner, K.R.; Berger, A. Molecular Detection of Late-Onset Neonatal Sepsis in Premature Infants Using Small Blood Volumes: Proof-of-Concept. Neonatology 2013, 103, 268–273. [Google Scholar] [CrossRef]
- Bomkamp, J.P.; Sulaiman, R.; Hartwell, J.L.; Desai, A.; Winn, V.C.; Wrin, J.; Kussin, M.L.; Hiles, J.J. Evaluation of a Rapid Fungal Detection Panel for Identification of Candidemia at an Academic Medical Center. J. Clin. Microbiol. 2019, 58. [Google Scholar] [CrossRef]
- Straub, J.; Paula, H.; Mayr, M.; Kasper, D.; Assadian, O.; Berger, A.; Rittenschober-Böhm, J. Diagnostic accuracy of the ROCHE Septifast PCR system for the rapid detection of blood pathogens in neonatal sepsis—A prospective clinical trial. PLoS ONE 2017, 12, e0187688. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- PathoNostics AsperGenius®. PathoNostics. Available online: https://www.pathonostics.com/product/aspergenius (accessed on 26 December 2020).
- Pelzer, B.W.; Seufert, R.; Koldehoff, M.; Liebregts, T.; Schmidt, D.; Buer, J.; Rath, P.-M.; Steinmann, J. Performance of the AsperGenius® PCR assay for detecting azole resistant Aspergillus fumigatus in BAL fluids from allogeneic HSCT recipients: A prospective cohort study from Essen, West Germany. Med. Mycol. 2019, 58, 268–271. [Google Scholar] [CrossRef]
- White, P.L.; Posso, R.B.; Barnes, R.A. Analytical and Clinical Evaluation of the PathoNostics AsperGenius Assay for Detection of Invasive Aspergillosis and Resistance to Azole Antifungal Drugs during Testing of Serum Samples. J. Clin. Microbiol. 2015, 53, 2115–2121. [Google Scholar] [CrossRef] [Green Version]
- Chong, G.M.; Van Der Beek, M.T.; Borne, P.A.V.D.; Boelens, J.; Steel, E.; Kampinga, G.A.; Span, L.F.R.; Lagrou, K.; Maertens, J.A.; Dingemans, G.J.H.; et al. PCR-based detection of Aspergillus fumigatus Cyp51A mutations on bronchoalveolar lavage: A multicentre validation of the AsperGenius assay ® in 201 patients with haematological disease suspected for invasive aspergillosis. J. Antimicrob. Chemother. 2016, 71, 3528–3535. [Google Scholar] [CrossRef] [Green Version]
- Chong, G.-L.M.; Van De Sande, W.W.J.; Dingemans, G.J.H.; Gaajetaan, G.R.; Vonk, A.G.; Hayette, M.-P.; Van Tegelen, D.W.E.; Simons, G.F.M.; Rijnders, B.J.A. Validation of a New Aspergillus Real-Time PCR Assay for Direct Detection of Aspergillus and Azole Resistance of Aspergillus fumigatus on Bronchoalveolar Lavage Fluid. J. Clin. Microbiol. 2015, 53, 868–874. [Google Scholar] [CrossRef] [Green Version]
- Ademtech MycoGENIE®. Real-Time PCR Kits—Ademtech. Available online: https://www.ademtech.com/molecular-diagnostic/mycology/real-time-pcr-kit/ (accessed on 26 December 2020).
- Chong, G.; Vonk, A.; Meis, J.; Dingemans, G.; Houbraken, J.; Hagen, F.; Gaajetaan, G.; Van Tegelen, D.; Simons, G.; Rijnders, B. Interspecies discrimination of A. fumigatus and siblings A. lentulus and A. felis of the Aspergillus section Fumigati using the AsperGenius® assay. Diagn. Microbiol. Infect. Dis. 2017, 87, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Morrell, M.; Fraser, V.J.; Kollef, M.H. Delaying the Empiric Treatment of Candida Bloodstream Infection until Positive Blood Culture Results Are Obtained: A Potential Risk Factor for Hospital Mortality. Antimicrob. Agents Chemother. 2005, 49, 3640–3645. [Google Scholar] [CrossRef] [Green Version]
- Ecker, D.J.; Sampath, R.; Massire, C.; Blyn, L.B.; Hall, T.A.; Eshoo, M.W.; Hofstadler, S.A. Ibis T5000: A universal biosensor approach for microbiology. Nat. Rev. Genet. 2008, 6, 553–558. [Google Scholar] [CrossRef]
- Wolk, D.M.; Kaleta, E.J.; Wysocki, V.H. PCR-electrospray ionization mass spectrometry: The potential to change infectious disease diagnostics in clinical and public health laboratories. J. Mol. Diagn. 2012, 14, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Simner, P.J.; Uhl, J.R.; Hall, L.; Weber, M.M.; Walchak, R.C.; Buckwalter, S.; Wengenack, N.L. Broad-Range Direct Detection and Identification of Fungi by Use of the PLEX-ID PCR-Electrospray Ionization Mass Spectrometry (ESI-MS) System. J. Clin. Microbiol. 2013, 51, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.H.; Ranken, R.; Sefers, S.E.; Lovari, R.; Quinn, C.D.; Meng, S.; Carolan, H.E.; Toleno, D.; Li, H.; Lee, J.N.; et al. Detection, Identification, and Distribution of Fungi in Bronchoalveolar Lavage Specimens by Use of Multilocus PCR Coupled with Electrospray Ionization/Mass Spectrometry. J. Clin. Microbiol. 2012, 51, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanio, A.; Garcia-Hermoso, D.; Mercier-Delarue, S.; Lanternier, F.; Gits-Muselli, M.; Menotti, J.; Denis, B.; Bergeron, A.; Legrand, M.; Lortholary, O.; et al. Molecular identification of Mucorales in human tissues: Contribution of PCR electrospray-ionization mass spectrometry. Clin. Microbiol. Infect. 2015, 21, 594.e1–594.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arastehfar, A.; Fang, W.; Pan, W.; Liao, W.; Yan, L.; Boekhout, T. Identification of nine cryptic species of Candida albicans, C. glabrata, and C. parapsilosis complexes using one-step multiplex PCR. BMC Infect. Dis. 2018, 18, 480. [Google Scholar] [CrossRef]
- Reyes-Montes, M.D.R.; Acosta-Altamirano, G.; Duarte-Escalante, E.; Salazar, E.G.; Martínez-Herrera, E.; Arenas, R.; González, G.; Frías-De-León, M.G. Usefulness of a multiplex PCR for the rapid identification of Candida glabrata species complex in Mexican clinical isolates. Rev. Inst. Med. Trop. São Paulo 2019, 61, e37. [Google Scholar] [CrossRef] [Green Version]
- Romeo, O.; Criseo, G. First molecular method for discriminating between Candida africana, Candida albicans, and Candida dubliniensis by using hwp1 gene. Diagn. Microbiol. Infect. Dis. 2008, 62, 230–233. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Canton, E.; Peman, J.; Dilger, A.; Romá, E.; Perlin, D.S. Assessment of Two New Molecular Methods for Identification of Candida parapsilosis Sensu Lato Species. J. Clin. Microbiol. 2011, 49, 3257–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavanti, A.; Davidson, A.D.; Gow, N.A.R.; Maiden, M.C.J.; Odds, F.C. Candida orthopsilosis and Candida metapsilosis spp. nov. to Replace Candida parapsilosis Groups II and III. J. Clin. Microbiol. 2005, 43, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Xu, J.; Shao, Y.; Gong, J.; Zhao, F.; He, L.; Shan, X. Rapid identification of the Candida glabrata species complex by high-resolution melting curve analysis. J. Clin. Lab. Anal. 2020, 34, e23226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theill, L.; Dudiuk, C.; Morano, S.; Gamarra, S.; Nardin, M.E.; Méndez, E.; Garcia-Effron, G. Prevalence and antifungal susceptibility of Candida albicans and its related species Candida dubliniensis and Candida africana isolated from vulvovaginal samples in a hospital of Argentina. Rev. Argent. Microbiol. 2016, 48, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Daneshnia, F.; Salehi, M.-R.; Zarrinfar, H.; Khodavaisy, S.; Haas, P.-J.; Roudbary, M.; Najafzadeh, M.-J.; Zomorodian, K.; Charsizadeh, A.; et al. Molecular characterization and antifungal susceptibility testing of Candida nivariensis from blood samples—An Iranian multicentre study and a review of the literature. J. Med Microbiol. 2019, 68, 770–777. [Google Scholar] [CrossRef]
- Morales-López, S.E.; Taverna, C.G.; Bosco-Borgeat, M.E.; Maldonado, I.; Vivot, W.; Szusz, W.; Garcia-Effron, G.; Córdoba, S.B. Candida glabrata species complex prevalence and antifungal susceptibility testing in a culture collection: First description of Candida nivariensis in Argentina. Mycopathologia 2016, 181, 871–878. [Google Scholar] [CrossRef]
- Theill, L.; Dudiuk, C.; Morales-Lopez, S.; Berrio, I.; Rodríguez, J.Y.; Marin, A.; Gamarra, S.; Garcia-Effron, G. Single-tube classical PCR for Candida auris and Candida haemulonii identification. Rev. Iberoam. Micol. 2018, 35, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Kordalewska, M.; Zhao, Y.; Lockhart, S.R.; Chowdhary, A.; Berrio, I.; Perlin, D.S. Rapid and Accurate Molecular Identification of the Emerging Multidrug-Resistant Pathogen Candida auris. J. Clin. Microbiol. 2017, 55, 2445–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgacopoulos, O.; Nunnally, N.S.; Le, N.; Lysen, C.; Welsh, R.M.; Kordalewska, M.; Perlin, D.S.; Berkow, E.L.; Sexton, D.J. Performance Evaluation of Culture-Independent SYBR Green Candida auris Quantitative PCR Diagnostics on Anterior Nares Surveillance Swabs. J. Clin. Microbiol. 2020, 58, 58. [Google Scholar] [CrossRef]
- Leach, L.; Zhu, Y.; Chaturvedi, S. Development and Validation of a Real-Time PCR Assay for Rapid Detection of Candida auris from Surveillance Samples. J. Clin. Microbiol. 2017, 56. [Google Scholar] [CrossRef] [Green Version]
- Sexton, D.J.; Kordalewska, M.; Bentz, M.L.; Welsh, R.M.; Perlin, D.S.; Litvintseva, A.P. Direct Detection of Emergent Fungal Pathogen Candida auris in Clinical Skin Swabs by SYBR Green-Based Quantitative PCR Assay. J. Clin. Microbiol. 2018, 56, e01337-18. [Google Scholar] [CrossRef] [Green Version]
- Kordalewska, M.; Lee, A.; Zhao, Y.; Perlin, D.S. Detection of Candida auris Antifungal Drug Resistance Markers Directly from Clinical Skin Swabs. Antimicrob. Agents Chemother. 2019, 63, 63. [Google Scholar] [CrossRef] [PubMed]
- Millon, L.; Scherer, E.; Rocchi, S.; Bellanger, A.-P. Molecular Strategies to Diagnose Mucormycosis. J. Fungi 2019, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Dannaoui, E. Molecular tools for identification of Zygomycetes and the diagnosis of zygomycosis. Clin. Microbiol. Infect. 2009, 15, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.P.; Bialek, R.; Milner, D.A.; Petschnigg, E.M.; Baden, L.R.; Marty, F.M. Molecular Methods to Improve Diagnosis and Identification of Mucormycosis: Fig. 1. J. Clin. Microbiol. 2011, 49, 2151–2153. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, P.; Bretagne, S.; Gantier, J.-C.; Garcia-Hermoso, D.; Lortholary, O.; Dromer, F.; Dannaoui, E. Molecular Identification of Zygomycetes from Culture and Experimentally Infected Tissues. J. Clin. Microbiol. 2006, 44, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Dannaoui, E.; Schwarz, P.; Slany, M.; Loeffler, J.; Jorde, A.T.; Cuenca-Estrella, M.; Hauser, P.M.; Shrief, R.; Huerre, M.; Freiberger, T.; et al. Molecular Detection and Identification of Zygomycetes Species from Paraffin-Embedded Tissues in a Murine Model of Disseminated Zygomycosis: A Collaborative European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Fungal Infection Study Group (EFISG) Evaluation. J. Clin. Microbiol. 2010, 48, 2043–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudiol, C.; Bodro, M.; Simonetti, A.; Tubau, F.; González-Barca, E.; Cisnal, M.; Domingo-Domenech, E.; Jiménez, L.; Carratalà, J. Changing aetiology, clinical features, antimicrobial resistance, and outcomes of bloodstream infection in neutropenic cancer patients. Clin. Microbiol. Infect. 2013, 19, 474–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, A.; Chen, S.; Sorrell, T.; Carter, D.; Malik, R.; Martin, P.; Halliday, C. Development and Clinical Application of a Panfungal PCR Assay to Detect and Identify Fungal DNA in Tissue Specimens. J. Clin. Microbiol. 2006, 45, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Bialek, R.; Konrad, F.; Kern, J.; Aepinus, C.; Cecenas, L.; Gonzalez, G.M.; Just-Nübling, G.; Willinger, B.; Presterl, E.; Lass-Flörl, C.; et al. PCR based identification and discrimination of agents of mucormycosis and aspergillosis in paraffin wax embedded tissue. J. Clin. Pathol. 2005, 58, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Machouart, M.-C.; Larche, J.; Burton, K.; Collomb, J.; Maurer, P.; Cintrat, A.; Biava, M.F.; Greciano, S.; Kuijpers, A.F.A.; Contet-Audonneau, N.; et al. Genetic Identification of the Main Opportunistic Mucorales by PCR-Restriction Fragment Length Polymorphism. J. Clin. Microbiol. 2006, 44, 805–810. [Google Scholar] [CrossRef] [Green Version]
- Baldin, C.; Soliman, S.S.M.; Jeon, H.H.; Alkhazraji, S.; Gebremariam, T.; Gu, Y.; Bruno, V.M.; Cornely, O.A.; Leather, H.L.; Sugrue, M.W.; et al. PCR-Based Approach Targeting Mucorales-Specific Gene Family for Diagnosis of Mucormycosis. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [Green Version]
- Scherer, E.; Iriart, X.; Bellanger, A.P.; Dupont, D.; Guitard, J.; Gabriel, F.; Cassaing, S.; Charpentier, E.; Guenounou, S.; Cornet, M.; et al. Quantitative PCR (qPCR) Detection of Mucorales DNA in Bronchoalveolar Lavage Fluid to Diagnose Pulmonary Mucormycosis. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Martínez, L.; Buitrago, M.J.; Castelli, M.V.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Development of a single tube multiplex real-time PCR to detect the most clinically relevant Mucormycetes species. Clin. Microbiol. Infect. 2013, 19, E1–E7. [Google Scholar] [CrossRef] [Green Version]
- Pujol, C.; Pfaller, M.A.; Soll, D.R. Flucytosine Resistance Is Restricted to a Single Genetic Clade of Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A.; Chowdhary, A.; Gonzalez, G.M.; Guinea, J.; Hagen, F.; Meis, J.F.; Thompson, G.R.; Turnidge, J. Multicenter Study of Isavuconazole MIC Distributions and Epidemiological Cutoff Values for the Cryptococcus neoformans-Cryptococcus gattii Species Complex Using the CLSI M27-A3 Broth Microdilution Method. Antimicrob. Agents Chemother. 2014, 59, 666–668. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A.; Aller, A.I.; Canton, E.; Castañón-Olivares, L.R.; Chowdhary, A.; Cordoba, S.; Cuenca-Estrella, M.; Fothergill, A.; Fuller, J.; Govender, N.; et al. Cryptococcus neoformans-Cryptococcus gattii Species Complex: An International Study of Wild-Type Susceptibility Endpoint Distributions and Epidemiological Cutoff Values for Fluconazole, Itraconazole, Posaconazole, and Voriconazole. Antimicrob. Agents Chemother. 2012, 56, 5898–5906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009–17) in India: Role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Nenoff, P.; Verma, S.B.; Ebert, A.; Süß, A.; Fischer, E.; Auerswald, E.; Dessoi, S.; Hofmann, W.; Schmidt, S.; Neubert, K.; et al. Spread of Terbinafine-Resistant Trichophyton mentagrophytes Type VIII (India) in Germany–“The Tip of the Iceberg?”. J. Fungi 2020, 6, 207. [Google Scholar] [CrossRef] [PubMed]
- Kanafani, Z.A.; Perfect, J.R. Resistance to Antifungal Agents: Mechanisms and Clinical Impact. Clin. Infect. Dis. 2008, 46, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinel-Ingroff, A.; Arendrup, M.; Cantón, E.; Cordoba, S.; Dannaoui, E.; García-Rodríguez, J.; Gonzalez, G.M.; Govender, N.P.; Martin-Mazuelos, E.; Lackner, M.; et al. Multicenter Study of Method-Dependent Epidemiological Cutoff Values for Detection of Resistance in Candida spp. and Aspergillus spp. to Amphotericin B and Echinocandins for the Etest Agar Diffusion Method. Antimicrob. Agents Chemother. 2016, 61. [Google Scholar] [CrossRef] [Green Version]
- Dick, J.D.; Merz, W.G.; Saral, R. Incidence of polyene-resistant yeasts recovered from clinical specimens. Antimicrob. Agents Chemother. 1980, 18, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Sokol-Anderson, M.L.; Brajtburg, J.; Medoff, G. Amphotericin B-Induced Oxidative Damage and Killing of Candida albicans. J. Infect. Dis. 1986, 154, 76–83. [Google Scholar] [CrossRef]
- Healey, K.R.; Perlin, D.S. Fungal Resistance to Echinocandins and the MDR Phenomenon in Candida glabrata. J. Fungi 2018, 4, 105. [Google Scholar]
- Bienvenu, A.L.; Leboucher, G.; Picot, S. Comparison of fks gene mutations and minimum inhibitory concentrations for the detection of Candida glabrata resistance to micafungin: A systematic review and meta-analysis. Mycoses 2019, 62, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Kwa, A.L.; Cheng, S.; Du, C.; Clancy, C.J. The Presence of anFKSMutation Rather than MIC Is an Independent Risk Factor for Failure of Echinocandin Therapy among Patients with Invasive Candidiasis Due to Candida glabrata. Antimicrob. Agents Chemother. 2012, 56, 4862–4869. [Google Scholar] [CrossRef] [Green Version]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole Resistance in Aspergillus spp.: A Worldwide Problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zoll, J.; Engel, T.; Heuvel, J.V.D.; Verweij, P.E.; Debets, A.J.M. The Medical Triazole Voriconazole Can Select for Tandem Repeat Variations in Azole-Resistant Aspergillus Fumigatus Harboring TR34/L98H via Asexual Reproduction. J. Fungi 2020, 6, 277. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.G.; Meis, J.F. Azole Resistance in Aspergillus fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole Resistance in Aspergillus Species: An Emerging Problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Sionov, E.; Chang, Y.C.; Garraffo, H.M.; Dolan, M.A.; Ghannoum, M.A.; Kwon-Chung, K.J. Identification of a Cryptococcus neoformans Cytochrome P450 Lanosterol 14α-Demethylase (Erg11) Residue Critical for Differential Susceptibility between Fluconazole/Voriconazole and Itraconazole/Posaconazole. Antimicrob. Agents Chemother. 2011, 56, 1162–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Chang, Y.C.; Kwon-Chung, K.J. Azole Heteroresistance in Cryptococcus neoformans: Emergence of Resistant Clones with Chromosomal Disomy in the Mouse Brain during Fluconazole Treatment. Antimicrob. Agents Chemother. 2013, 57, 5127–5130. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.; Sionov, E.; Lamichhane, A.K.; Kwon-Chung, K.J.; Chang, Y.C. Roles of Three Cryptococcus neoformans and Cryptococcus gattii Efflux Pump-Coding Genes in Response to Drug Treatment. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lofgren, L.A.; Uehling, J.K.; Branco, S.; Bruns, T.D.; Martin, F.; Kennedy, P.G. Genome-based estimates of fungal rDNA copy number variation across phylogenetic scales and ecological lifestyles. Mol. Ecol. 2019, 28, 721–730. [Google Scholar] [CrossRef]
- Mellado, E.; Garcia-Effron, G.; Alcázar-Fuoli, L.; Melchers, W.J.G.; Verweij, P.E.; Cuenca-Estrella, M.; Rodríguez-Tudela, J.L. A New Aspergillus fumigatus Resistance Mechanism Conferring In Vitro Cross-Resistance to Azole Antifungals Involves a Combination of cyp51A Alterations. Antimicrob. Agents Chemother. 2007, 51, 1897–1904. [Google Scholar] [CrossRef] [Green Version]
- Mellado, E.; Garcia-Effron, G.; Alcazar-Fuoli, L.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Substitutions at Methionine 220 in the 14α-Sterol Demethylase (Cyp51A) of Aspergillus fumigatus Are Responsible for Resistance In Vitro to Azole Antifungal Drugs. Antimicrob. Agents Chemother. 2004, 48, 2747–2750. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Guerra, T.M.; Mellado, E.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. A Point Mutation in the 14α-Sterol Demethylase Gene cyp51A Contributes to Itraconazole Resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2003, 47, 1120–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Effron, G.; Lee, S.; Park, S.; Cleary, J.D.; Perlin, D.S. Effect of Candida glabrata FKS1 and FKS2 Mutations on Echinocandin Sensitivity and Kinetics of 1,3-β-d-Glucan Synthase: Implication for the Existing Susceptibility Breakpoint. Antimicrob. Agents Chemother. 2009, 53, 3690–3699. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Effron, G.; Park, S.; Perlin, D.S. Correlating Echinocandin MIC and Kinetic Inhibition of fks1 Mutant Glucan Synthases for Candida albicans: Implications for Interpretive Breakpoints. Antimicrob. Agents Chemother. 2008, 53, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marichal, P.; Koymans, L.; Willemsens, S.; Bellens, D.; Verhasselt, P.; Luyten, W.; Borgers, M.; Ramaekers, F.C.S.; Odds, F.C.; Bossche, H.V. Contribution of mutations in the cytochrome P450 14α-demethylase (Erg11p, Cyp51p) to azole resistance in Candida albicans. Microbiology 1999, 145, 2701–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Effron, G.; Dilger, A.; Alcazar-Fuoli, L.; Park, S.; Mellado, E.; Perlin, D.S. Rapid Detection of Triazole Antifungal Resistance in Aspergillus fumigatus. J. Clin. Microbiol. 2008, 46, 1200–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Linden, J.W.M.; Snelders, E.; Arends, J.P.; Daenen, S.M.; Melchers, W.J.G.; Verweij, P.E. Rapid Diagnosis of Azole-Resistant Aspergillosis by Direct PCR Using Tissue Specimens. J. Clin. Microbiol. 2010, 48, 1478–1480. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Park, S.; Lass-Florl, C.; Fraczek, M.G.; Kirwan, M.; Gore, R.; Smith, J.; Bueid, A.; Moore, C.B.; Bowyer, P.; et al. High-frequency Triazole Resistance Found in Nonculturable Aspergillus fumigatus from Lungs of Patients with Chronic Fungal Disease. Clin. Infect. Dis. 2011, 52, 1123–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaassen, C.H.W.; De Valk, H.A.; Curfs-Breuker, I.M.; Meis, J.F. Novel mixed-format real-time PCR assay to detect mutations conferring resistance to triazoles in Aspergillus fumigatus and prevalence of multi-triazole resistance among clinical isolates in the Netherlands. J. Antimicrob. Chemother. 2010, 65, 901–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Kathuria, S.; Randhawa, H.S.; Gaur, S.N.; Klaassen, C.H.; Meis, J.F. Isolation of multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR/L98H mutations in the cyp51A gene in India. J. Antimicrob. Chemother. 2011, 67, 362–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiess, B.; Seifarth, W.; Merker, N.; Howard, S.J.; Reinwald, M.; Dietz, A.; Hofmann, W.-K.; Buchheidt, D. Development of Novel PCR Assays to Detect Azole Resistance-Mediating Mutations of theAspergillus fumigatus cyp51AGene in Primary Clinical Samples from Neutropenic Patients. Antimicrob. Agents Chemother. 2012, 56, 3905–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiess, B.; Postina, P.; Reinwald, M.; Cornely, O.A.; Hamprecht, A.; Hoenigl, M.; Lass-Flörl, C.; Rath, P.-M.; Steinmann, J.; Miethke, T.; et al. Incidence of Cyp51 A Key Mutations in Aspergillus fumigatus—A Study on Primary Clinical Samples of Immunocompromised Patients in the Period of 1995–2013. PLoS ONE 2014, 9, e103113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Khan, Z.; Hagen, F.; Meis, J.F. Simple, Low-Cost Molecular Assays for TR34/L98H Mutations in the cyp51A Gene for Rapid Detection of Triazole-Resistant Aspergillus fumigatus Isolates. J. Clin. Microbiol. 2014, 52, 2223–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudakova, A.; Spiess, B.; Tangwattanachuleeporn, M.; Sasse, C.; Buchheidt, D.; Weig, M.; Groß, U.; Bader, O. Molecular Tools for the Detection and Deduction of Azole Antifungal Drug Resistance Phenotypes in Aspergillus Species. Clin. Microbiol. Rev. 2017, 30, 1065–1091. [Google Scholar] [CrossRef] [Green Version]
- Araújo, R.; Gungor, O.; Amorim, A. Single-tube PCR coupled with mini-sequencing assay for the detection of cyp51A and cyp51B polymorphisms in Aspergillus fumigatus. Futur. Microbiol. 2015, 10, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, F.; Hashemi, S.J.; Zoll, J.; Melchers, W.J.G.; Rafati, H.; Dehghan, P.; Rezaie, S.; Tolooe, A.; Tamadon, Y.; Van Der Lee, H.A.; et al. Quantitative Analysis of Single-Nucleotide Polymorphism for Rapid Detection of TR34/L98H and TR46/Y121F/T289A-Positive Aspergillus fumigatus Isolates Obtained from Patients in Iran from 2010 to 2014. Antimicrob. Agents Chemother. 2015, 60, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraczek, M.G.; Bromley, M.; Buied, A.; Moore, C.B.; Rajendran, R.; Rautemaa, R.; Ramage, G.; Denning, D.W.; Bowyer, P. The cdr1B efflux transporter is associated with non-cyp51a-mediated itraconazole resistance in Aspergillus fumigatus. J. Antimicrob. Chemother. 2013, 68, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.-S.; Rodriguez-Manzano, J.; Malpartida-Cardenas, K.; Sewell, T.; Bader, O.; Armstrong-James, D.; Fisher, M.C.; Georgiou, P. Rapid and Sensitive Detection of Azole-Resistant Aspergillus fumigatus by Tandem Repeat Loop-Mediated Isothermal Amplification. J. Mol. Diagn. 2019, 21, 286–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macedo, D.; Devoto, T.B.; Pola, S.; Finquelievich, J.L.; Cuestas, M.L.; García-Effron, G. A Novel Combination of CYP51A Mutations Confers Pan-Azole Resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2020, 64, 64. [Google Scholar] [CrossRef]
- Hagiwara, D.; Takahashi, H.; Watanabe, A.; Takahashi-Nakaguchi, A.; Kawamoto, S.; Kamei, K.; Gonoi, T. Whole-Genome Comparison of Aspergillus fumigatus Strains Serially Isolated from Patients with Aspergillosis. J. Clin. Microbiol. 2014, 52, 4202–4209. [Google Scholar] [CrossRef] [Green Version]
- Montesinos, I.; Argudín, M.A.; Hites, M.; Ahajjam, F.; Dodémont, M.; Dagyaran, C.; Bakkali, M.; Etienne, I.; Jacobs, F.; Knoop, C.; et al. Culture-Based Methods and Molecular Tools for Azole-Resistant Aspergillus fumigatus Detection in a Belgian University Hospital. J. Clin. Microbiol. 2017, 55, 2391–2399. [Google Scholar] [CrossRef] [Green Version]
- Dannaoui, E.; Gabriel, F.; Gaboyard, M.; Lagardere, G.; Audebert, L.; Quesne, G.; Godichaud, S.; Verweij, P.E.; Accoceberry, I.; Bougnoux, M.-E. Molecular Diagnosis of Invasive Aspergillosis and Detection of Azole Resistance by a Newly Commercialized PCR Kit. J. Clin. Microbiol. 2017, 55, 3210–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Nagasaki, Y.; Kordalewska, M.; Press, E.G.; Shields, R.K.; Nguyen, M.H.; Clancy, C.J.; Perlin, D.S. Rapid Detection ofFKS-Associated Echinocandin Resistance in Candida glabrata. Antimicrob. Agents Chemother. 2016, 60, 6573–6577. [Google Scholar] [CrossRef] [Green Version]
- Dudiuk, C.; Gamarra, S.; Leonardeli, F.; Jimenez-Ortigosa, C.; Vitale, R.G.; Afeltra, J.; Perlin, D.S.; Garcia-Effron, G. Set of Classical PCRs for Detection of Mutations in Candida glabrata FKS Genes Linked with Echinocandin Resistance. J. Clin. Microbiol. 2014, 52, 2609–2614. [Google Scholar] [CrossRef] [Green Version]
- Biswas, C.; Chen, S.C.-A.; Halliday, C.; Martínez, E.; Rockett, R.J.; Wang, Q.; Timms, V.J.; Dhakal, R.; Sadsad, R.; Kennedy, K.J.; et al. Whole Genome Sequencing of Candida glabrata for Detection of Markers of Antifungal Drug Resistance. J. Vis. Exp. 2017, 56714, e56714. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.D.; Bolden, C.B.; Kuykendall, R.J.; Lockhart, S.R. Development of a Luminex-Based Multiplex Assay for Detection of Mutations Conferring Resistance to Echinocandins in Candida glabrata. J. Clin. Microbiol. 2013, 52, 790–795. [Google Scholar] [CrossRef] [Green Version]
- Dudiuk, C.; Gamarra, S.; Jiménez-Ortigosa, C.; Leonardelli, F.; Macedo, D.; Perlin, D.S.; Garcia-Effron, G. Quick Detection of FKS1Mutations Responsible for Clinical Echinocandin Resistance in Candida albicans. J. Clin. Microbiol. 2015, 53, 2037–2041. [Google Scholar] [CrossRef] [Green Version]
- Balashov, S.V.; Park, S.; Perlin, D.S. Assessing Resistance to the Echinocandin Antifungal Drug Caspofungin in Candida albicans by Profiling Mutations in FKS1. Antimicrob. Agents Chemother. 2006, 50, 2058–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnaud, C.; Botterel, F.; Sertour, N.; Bougnoux, M.-E.; Dannaoui, E.; Larrat, S.; Hennequin, C.; Guinea, J.; Cornet, M.; Maubon, D. Next-generation sequencing offers new insights into the resistance of Candida spp. to echinocandins and azoles. J. Antimicrob. Chemother. 2015, 70, 2556–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, M.; Deshpande, L.M.; Davis, A.P.; Rhomberg, P.R.; Pfaller, M.A. Monitoring Antifungal Resistance in a Global Collection of Invasive Yeasts and Molds: Application of CLSI Epidemiological Cutoff Values and Whole-Genome Sequencing Analysis for Detection of Azole Resistance in Candida albicans. Antimicrob. Agents Chemother. 2017, 61, e00906-17. [Google Scholar] [CrossRef] [Green Version]
- Healey, K.R.; Kordalewska, M.; Ortigosa, C.J.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 Mutations Identified in Isolates of Candida auris Directly Contribute to Reduced Azole Susceptibility. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Lee, A.; Jiménez-Ortigosa, C.; Kordalewska, M.; Perlin, D.S.; Zhao, Y. Rapid Detection of ERG11-Associated Azole Resistance and FKS-Associated Echinocandin Resistance in Candida auris. Antimicrob. Agents Chemother. 2018, 63, e01811-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morio, F.; Loge, C.; Besse, B.; Hennequin, C.; Le Pape, P. Screening for amino acid substitutions in the Candida albicans Erg11 protein of azole-susceptible and azole-resistant clinical isolates: New substitutions and a review of the literature. Diagn. Microbiol. Infect. Dis. 2010, 66, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.S.; Rolley, N.; Kelly, D.E.; Kelly, S.L. Identification and Characterization of Four Azole-Resistant erg3 Mutants of Candida albicans. Antimicrob. Agents Chemother. 2010, 54, 4527–4533. [Google Scholar] [CrossRef] [Green Version]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and Isochromosome Formation in Drug-Resistant Candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, C.; Bhattacharya, S.; Wakabayashi, H.; Bellaousov, S.; Kravets, A.; Welle, S.L.; Myers, J.; Hayes, J.J.; Bulger, M.; Rustchenko, E. Transcriptional Regulation on Aneuploid Chromosomes in Divers Candida albicans Mutants. Sci. Rep. 2018, 8, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coste, A.; Turner, V.; Ischer, F.; Morschhäuser, J.; Forche, A.; Selmecki, A.; Berman, J.; Bille, J.; Sanglard, D. A Mutation in Tac1p, a Transcription Factor Regulating CDR1 and CDR2, Is Coupled with Loss of Heterozygosity at Chromosome 5 to Mediate Antifungal Resistance in Candida albicans. Genetics 2006, 172, 2139–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Kravets, A.; Bethlendy, G.; Welle, S.; Rustchenko, E. Chromosome 5 Monosomy of Candida albicans Controls Susceptibility to Various Toxic Agents, Including Major Antifungals. Antimicrob. Agents Chemother. 2013, 57, 5026–5036. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S. Antifungal drug resistance: Do molecular methods provide a way forward? Curr. Opin. Infect. Dis. 2009, 22, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rex, J.H.; Pfaller, M.A. Has antifungal susceptibility testing come of age? Clin. Infect. Dis. 2002, 35, 982–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Format | Target and Technique Characteristics | Samples | Detected Mechanism | Points to Consider | Reference/Publication Year |
|---|---|---|---|---|---|
| Real-time PCR with molecular beacons | Two panels of multiplex PCRs. The first detects ITC and cross azole resistance. The second detects PSC resistance and confirms the coexistence of TR and L98H. | Isolates strains | G54X (ITC-R), G54W (ITC/PSC-R), M220X (ITC-R), G138X/C * (Cross-R), TR34-L98H (Cross-R) | 60 strains (52 clinical and 8 lab mutants) harboring G54X, M220X, G138C, TR34-L98H, TR34 alone, and L98H alone. | [126] 2008 |
| Real-time PCR using Taq-man probes | CYP51A ORF and promoter | Formalin-fixed and paraffin-embedded tissue | TR34-L98H | Only one patient | [127]/2010 |
| Nested 2 step PCR. Firstly classical PCR in 2 tubes (outer). Second PCR in real-time format with molecular beacons | CYP51A ORF (first classical PCR) and promoter plus a partial ORF amplification (second classical PCR). Molecular beacons bind to secondly amplified targets. | Sputum and BAL | TR34-L98H G54, G138, and M220. | DNA extraction using the MycXtra fungal DNA extraction kit (Myconostica Ltd.). 22 samples 5 proven and 17 probable aspergilloses. | [128]/2011 |
| Real-time PCR–FRET probes with melting curves analysis | CYP51A ORF and promoter. | Isolated strains (clinical) | TR34-L98H G54, G138, and M220. | 215 [129] and 103 [130] A. fumigatus sensu stricto included. TR34-L98H (n = 4) was the only detected mechanism. There were no G54, G138, and M220 mutations (only wild type confirmation) | [129,130]/2010 and 2012 |
| Classical PCR and nested PCR followed by sequencing. | 3 individual PCRs able to amplify CYP51A promoter and fractions of its ORF | Isolated strains [131] and clinical samples (BAL and tissue) [132] | TR34-L98H and M220 | Developed using strains and clinical samples [131] and tested in a clinical setting [132]. | [131,132] 2012/2014 |
| PCR-RFLP (amplification followed by AluI digestion) | A promoter and a CYP51A ORF fragment (289 bp) [133]. Later, a bigger fragment was used [134]. | Isolated strains (clinical and environmental) | TR34-L98H [133] and TR34-L98H and TR46-Y121F-T289A [134] | Good correlation with MIC but false negative (isolates harboring other mechanisms) | [128,130] 2014 and 2017 |
| Single tube PCR followed by minisequencing | A multiplex classical PCR followed by purification and detection of 21 SNPs at CYP51A and CYP51B by single-base extension reaction (SNaPshotTM) using a Sanger-based sequencer. | Isolated strains | TR34, G54, L98, G138, M220, S297, G448, and 12 CYP51A polymorphisms. Two CYP51B polymorphisms were also included. | 79 clinical and 21 environmental isolates. No resistant mutants but several with CYP51A and CYP51B polymorphism. | [135] 2015 |
| Real-time PCR with locked nucleotide probes | Partial CYP51A ORF amplification. Detection of both wild type and mutant (L98H and Y121F) alleles. | Isolated strains | TR34-L98H and TR46-Y121F-T289A. It only detects the mutation in the ORF, not the promoter alteration | Detection of 6 L98H mutants/166. No Y121F mutants were detected. | [136] 2016 |
| Quantitative real-time PCR with sybrgreen | Different efflux pump genes. | 10 clinical strains with high azole MICs. | Detection of overexpression of efflux pumps | 80% of the strains showed > 5 –fold increase of cdr1B gene expression. | [137]/2013 |
| Loop-mediated isothermal amplification (LAMP) | TR34 promoter alteration | Clinical strains | Detection of TR34 alone. | Rapid (<25 min) High sensitivity (10 genomic copies). | [138]/2019 |
| Classical PCR using astringent conditions | TR34-R65K-L98H | Clinical strains | Detection of R65K mutation and TR34. | Low-cost detection of a triazole-cross resistance (TR34) and pan azole resistance (R65K) | [139]/2020 |
| Whole-genome sequencing | Complete genome | Isolates from sequential clinical samples from two patients (one Aspergilloma and one invasive aspergillosis) | P216L | Complex to perform in a clinical setting. Potential to uncover any mechanism after analysis | [140]/2014 |
| AsperGenius multiplex real-time PCR assay. | Two panels. The first is a taxonomy panel (based on 28S rDNA). The second is A. fumigatus sensu stricto resistance detection (melting curve analysis). | Isolated strains (n = 131) [63] were used for validation. Clinical samples. BAL (n = 22) [63] (n = 201) [62] (n = 124) [61] (n = 100) [141] were used for clinical evaluation (most from proven and probable invasive aspergillosis). | Detects intrinsic resistant species (A. terreus and cryptic species of the Fumigatii section) and the following secondary resistance markers separately: TR34, L98H, Y121F, and T289A. | Some cross-reactivity (false-positive results) were obtained when R. oryzae (R. arrhizus) and P. chrysogenum DNA is present at high concentrations. During validation, high sensitivity and specificity were proved (both >80%) [62]. TR34-L98H was the most prevalent mechanism [61,62,141] | [61,62,63,141]/2017–2015-2016–2016 |
| MycoGENIE multiplex real-time PCR assay. | Identification of A. fumigatus sensu stricto (based on rDNA sequence) and TR34-L98H detection | Clinical samples | Possible false positive when aspergillosis is caused by non-Aspergillus fumigatus species. | [142]/2017 |
| Organism | Format | Target and Technique Characteristics | Samples | Detected Mechanism | Points to Consider | Reference/Publication Year |
|---|---|---|---|---|---|---|
| Candida glabrata | Asymmetric real-time PCR coupled with molecular beacons with melting curve analysis | FKS1 and FKS2 hot spot 1 regions. | Isolated strains | Echinocandin resistance. 4 amino acid substitutions at Fks1p (F625, S629, D632, and I634) and 2 at Fks2p (F659 and S663). Melting curve analysis can differentiate different nucleotide substitutions in the same position (8 at FKS1 and 7 at FKS2). | Tested with a blinded panel of 188 strains. 100% concordance with sequencing. | [143]/2016 |
| Classical PCR with astringent conditions | FKS1 and FKS2 hot spot 1 regions. | Isolated strains | Echinocandin resistance. 3 amino acid substitutions at Fks1p (F625, S6229, and D632) and 2 at Fks2p (F659 and S663). | Tested with a blinded panel of 50 strains. Not able to detect F659del mutants [144]. It was tested later and showed a 99.25% concordance with MIC values. One strain was misclassified as resistant due to a silent mutation [145]. | [36,144]/2014 and 2017 | |
| High-throughput microsphere-based assay using the Luminex MagPix technology | FKS1 hot spot 1 and hot spot 2 regions. | Strain collection | Echinocandin resistance. It potentially can detect all the FKS mutations. | Screen a collection of 1032 strains. | [146]/2014 | |
| Candida albicans | Classical PCR with astringent | FKS1 hot spot 1 and hot spot 2 regions. | Isolated strains | Echinocandin resistance. It detects 8 different substitutions at 5 Fks1p residues. Four at hot spot 1 (F641, S645, D648, P649) and one at hot spot 2 (R1361) | 96% sensitivity. It can detect all the homozygous mutants included. Heterozygous mutants give false susceptibility due to method-inherent limitations of the classical PCR. | [147]/2015 |
| Candida albicans | Allele-specific real-time PCR molecular-beacon | FKS1 hot spot 1 | Laboratory mutants generated by CSF pressure. | Echinocandin resistance. It detects 4 substitutions at the residue S645. | It was the first published method. It gave the proof of concept that it is possible to detect FKS mutations. Currently outdated. | [148]/2006 |
| Candida albicans, Candida glabrata and Candida parapsilosis | NGS | 6 genes linked with antifungal resistance (ERG11, ERG3, TAC1, CgPDR1, FKS1, and FKS2) | Isolated strains. For validation, resistant strains with known mechanisms. Then, clinical resistant strains. | Azole and echinocandin resistance. New mechanisms were uncovered including one gain-of-function and one loss-of-function CgPDR1 mutations responsible for azole resistance and hypersusceptibility, respectively. | It demonstrates that a mixed population (mutated and WT) would be isolated from a patient during caspofungin treatment. It gave the proof of concept that it is possible to use NGS for extensive assessment of mutations responsible for antifungal resistance | [149]/2015 |
| Candida albicans and Candida glabrata | WGS and Sanger sequencing | Sanger sequencing of FKS hot spot regions and WGS for azole resistance markers | Isolated strains | FKS, ERG11, ERG3, UPC2, MDR1, MRR1, TAC1, CDR1, and CDR2. | Strains showing echinocandin and/or azole high MIC values were studied. Used as a research tool and not as a diagnostic tool. | [150]/2017 |
| Candida auris | WGS | Isolated strains | ERG11 mutations | Worldwide strains were divided into 4 clades and each clade showed differential FLC susceptibility (Clade II lower MIC- no ERG11 mutations) | [102]/2017 | |
| Candida auris | Asymmetric real-time PCR coupled with molecular beacons with melting curve analysis | One FKS1 and two ERG11 mutations. | Isolated strains | Echinocandin and FLC resistance. It can detect the main mechanisms of azole resistance (Y132F and K143R in Erg11p) and echinocandin resistance (S639F in Fks1p) in this species. | Some strains belonging to clade IV (South America) would be identified as false FLC susceptible since the most prevalent mechanism of FLC resistance is the substitution I466M [151]. | [152]/2019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Effron, G. Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives. J. Fungi 2021, 7, 197. https://doi.org/10.3390/jof7030197
Garcia-Effron G. Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives. Journal of Fungi. 2021; 7(3):197. https://doi.org/10.3390/jof7030197
Chicago/Turabian StyleGarcia-Effron, Guillermo. 2021. "Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives" Journal of Fungi 7, no. 3: 197. https://doi.org/10.3390/jof7030197
APA StyleGarcia-Effron, G. (2021). Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives. Journal of Fungi, 7(3), 197. https://doi.org/10.3390/jof7030197