
Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
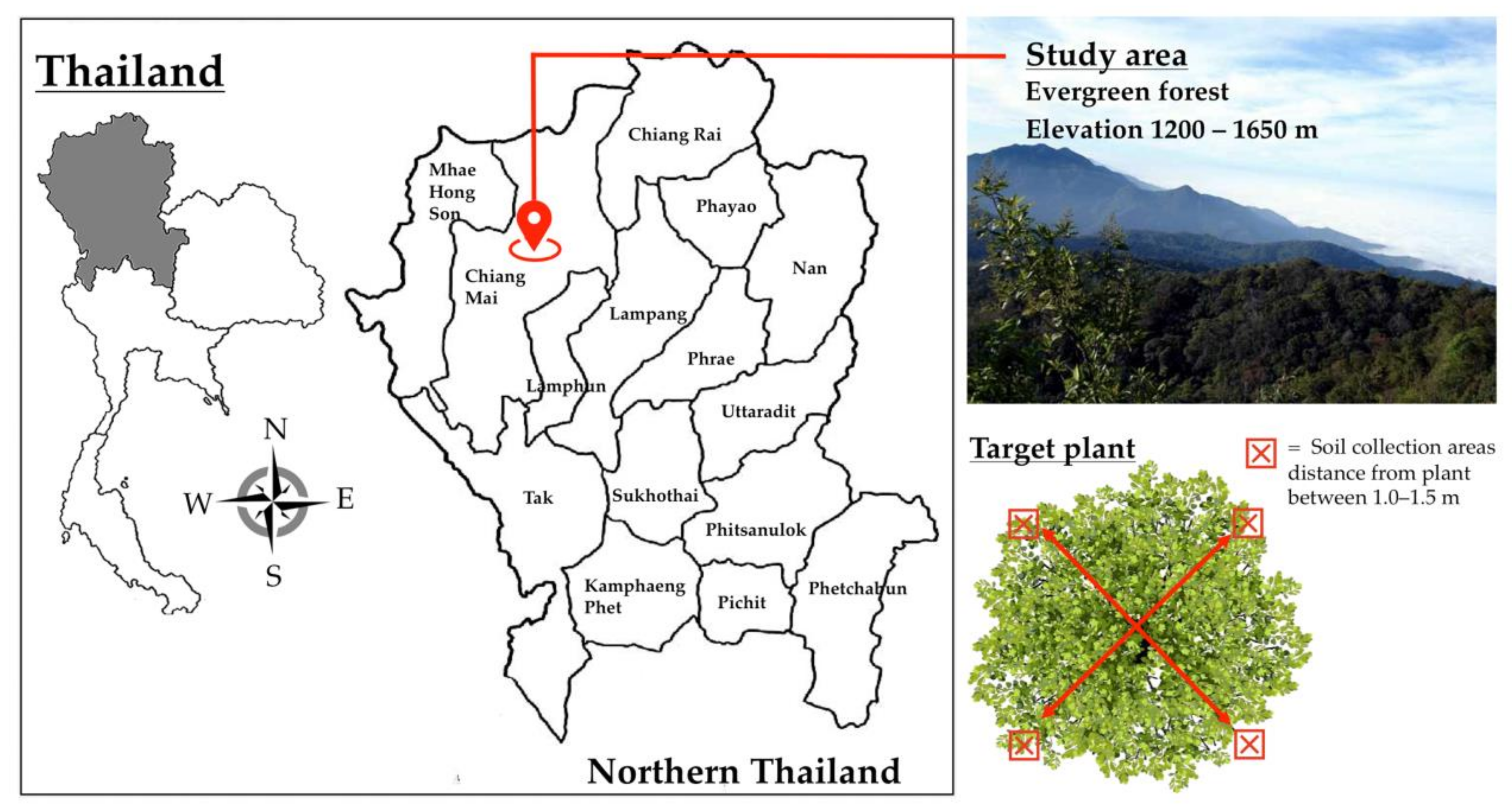
2.2. Soil Sampling and eDNA Extraction
2.3. PCR Amplification, Amplicon Processing, and Illumina Sequencing
2.4. PCR Sequence Analysis and Taxonomical Assignment
2.5. Truffle Hunting and Identification
3. Results
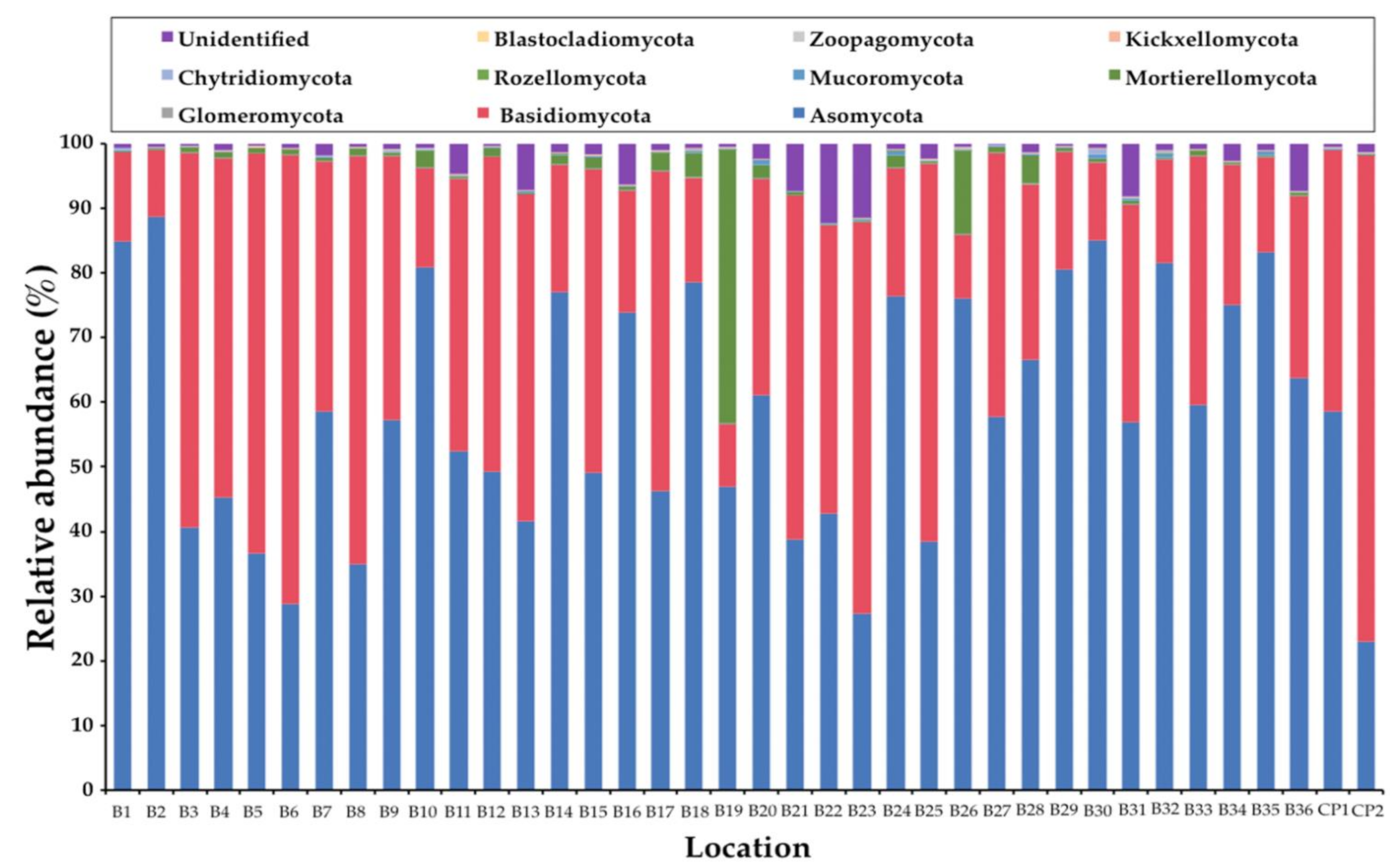
3.1. General Soil Fungal Composition
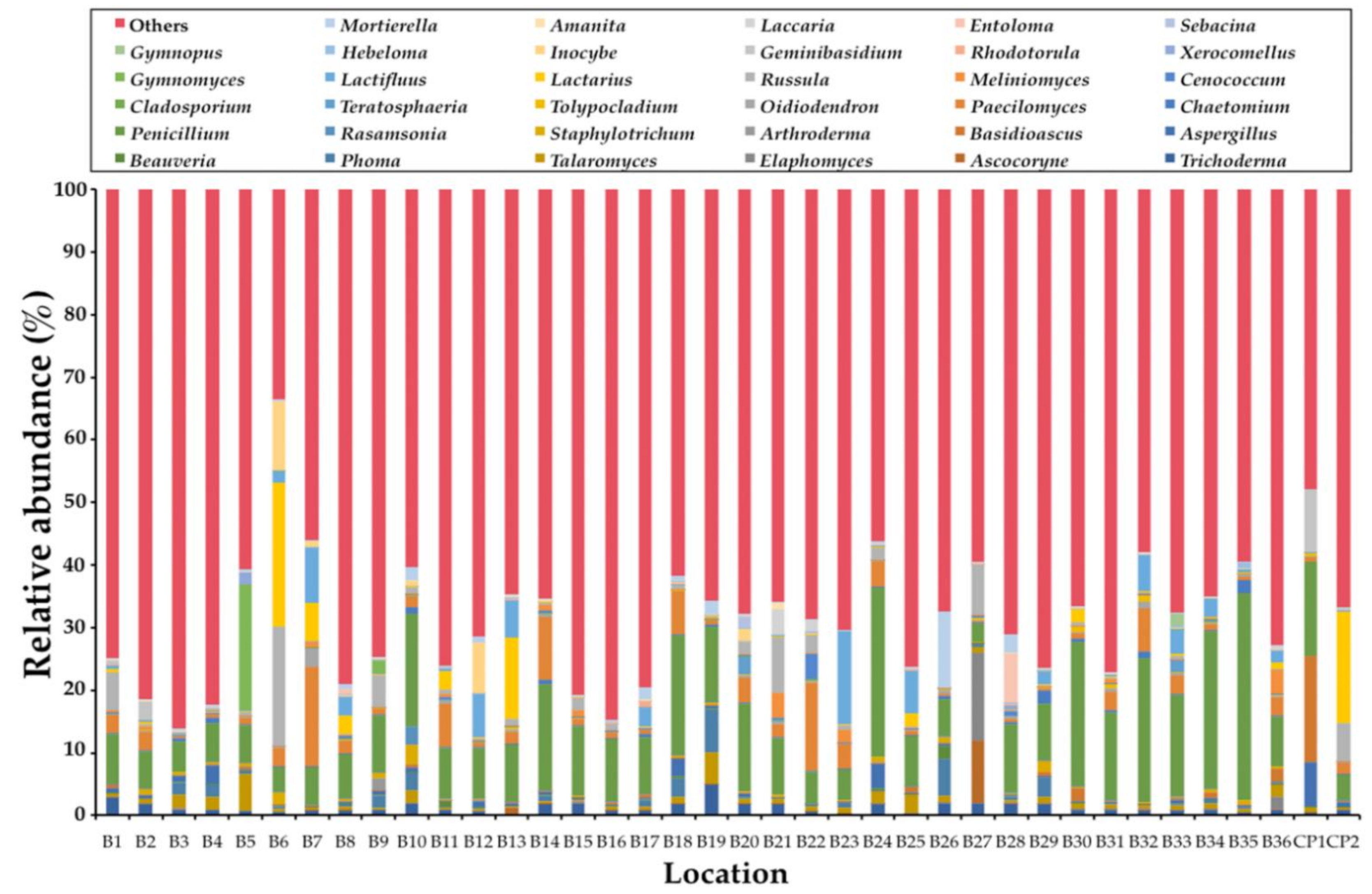
3.2. OTUs of Truffles (Tuber Species) in Soil Samples
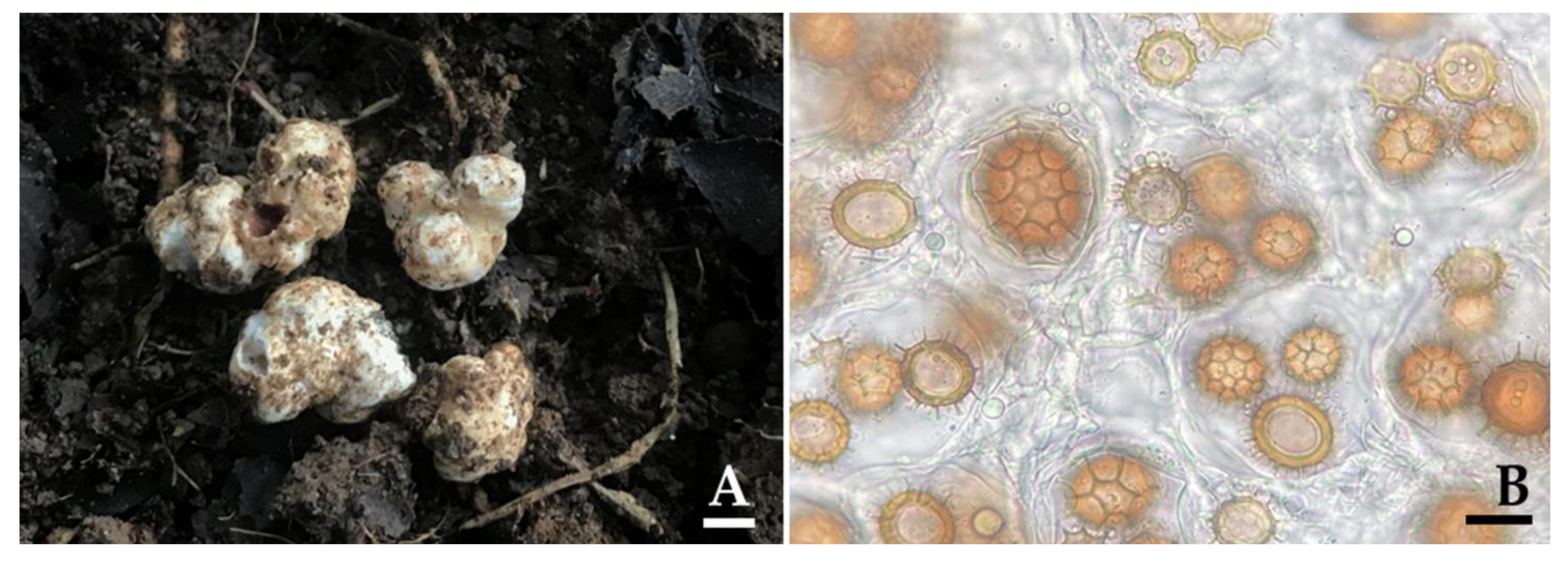
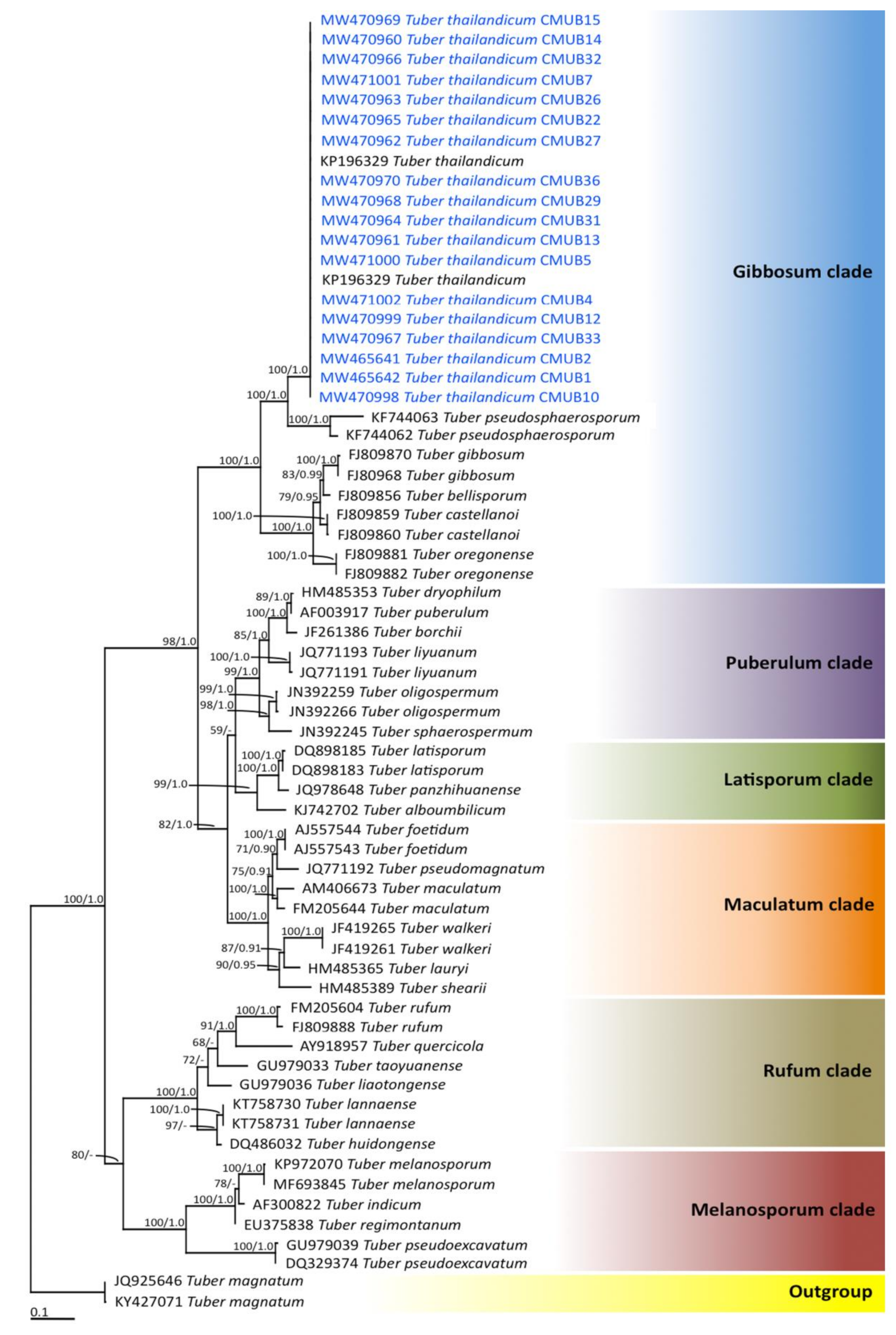
3.3. Truffle Hunting and Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Hall, I.R.; Brown, G.T.; Zambonelli, A. Taming the Truffle, the History, Lore and Science of the Ultimate Mushroom; Timber Press: Portland, OR, USA, 2007. [Google Scholar]
- Bonito, G.; Grygansyi, A.P.; Trappe, J.M.; Vilgalys, R. A global metaanalysis of Tuber ITS rDNA sequences: Species diversity, host associations and long-distance dispersal. Mol. Ecol. 2010, 19, 4994–5008. [Google Scholar] [CrossRef] [PubMed]
- Zambonelli, A.; Iotti, M.; Murat, C. True Truffle (Tuber spp.) in the World, 1st ed.; Springer International Publishing AG: Basel, Switzerland, 2016. [Google Scholar]
- Splivallo, R.; Valdez, N.; Kirchhoff, N.; Ona, M.C.; Schmidt, J.P.; Feussner, I.; Karlovsky, P. Intraspecific genotypic variability determines concentrations of key truffle volatiles. New Phytol. 2012, 194, 823–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Splivallo, R.; Ottonello, S.; Mello, A.; Karlovsky, P. Truffle volatiles: From chemical ecology to aroma biosynthesis. New Phyt. 2011, 189, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Trappe, J.M.; Molina, R.; Luoma, D.L.; Cázares, E.; Pilz, D.; Smith, J.E.; Castellano, M.A.; Miller, S.L.; Trappe, M.J. Diversity, ecology, and conservation of truffle fungi in forests of the pacific northwest; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Stobbe, U.; Egli, S.; Tegel, W.; Peter, M.; Sproll, L.; Büntgen, U. Potential and limitation of Burgundy truffle cultivation. Appl. Microbiol. Biotechnol. 2013, 97, 5215–5224. [Google Scholar] [CrossRef]
- Hall, I.R.; Yun, W.; Amicucci, A. Cultivation of edible ectomycorrhizal mushrooms. Trends Biotechnol. 2013, 21, 433–438. [Google Scholar] [CrossRef]
- Bonito, G.; Trappe, J.M.; Rawlimson, P.; Vilgalys, R. Improved resolution of major clades whitish Tuber and taxonomy of species within the Tuber gibbosum complex. Mycologia 2010, 102, 1042–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.E.; Bonito, G.; Sharma, J.; Long, J.; Davis-Long, B.; Brenneman, T. Pecan Truffles (Tuber lyonii) What We Know and What We Need to Know. Georgia Pecan Magazine 2012, 52–58. [Google Scholar]
- Mortimer, P.E.; Karunarathna, S.C.; Li, Q.; Gui, H.; Yang, X.; Yang, X.; He, J.; Ye, L.; Guo, J.; Li, H.; et al. Prized edible Asian mushrooms: Ecology, conservation and sustainability. Fungal Divers. 2012, 56, 31–47. [Google Scholar] [CrossRef]
- Chen, Y.L.; Gong, M.Q. Truffles Tuber spp.: Biodiversity and geological distribution. Edible Fungi of China 2000, 19, 25–26. [Google Scholar]
- Chen, J.; Liu, P.G.; Wang, Y. Tuber umbilicatum, a new species from China, with a key to the spinose-reticulate spored Tuber species. Mycotaxon 2005, 94, 1–6. [Google Scholar]
- Deng, X.J.; Liu, P.G.; Liu, C.Y.; Wang, Y. A new white truffle species, Tuber panzhihuanense from China. Mycol. Prog. 2013, 12, 557–561. [Google Scholar] [CrossRef]
- Fan, L.; Cao, J.Z. Two new species of white truffle from China. Mycotaxon 2012, 121, 297–304. [Google Scholar] [CrossRef]
- García-Montero, L.G.; Díaz, P.; Massimo, G.D.; García-Abril, A. A review of research on Chinese Tuber species. Mycol. Prog. 2010, 9, 315–335. [Google Scholar] [CrossRef]
- Kinoshima, A.; Sasaki, H.; Nara, K. Two new truffle species, Tuber japonicum and Tuber flavidosporum spp. nov. found from Japan. Mycoscience 2016, 57, 366–373. [Google Scholar] [CrossRef]
- Fan, L.; Liu, X.; Cao, J. Tuber turmericum sp. nov., a Chinese truffle species based on morphological and molecular data. Mycol. Prog. 2015, 14, 111. [Google Scholar] [CrossRef]
- Kinoshita, A.; Sasaki, H.; Nara, K. Phylogeny and diversity of Japanese truffles (Tuber spp.) inferred from sequences of four nuclear loci. Mycologia 2011, 103, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity–up to 96% of fungi in northern Thailand are novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Lumyong, S. A new whitish truffle, Tuber thailandicum from northern Thailand and its ectomycorrhizal association. Mycol. Prog. 2015, 14, 83. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Vadthanarat, S.; Raspé, O.; Lumyong, S. Morphology and molecular evidence support a new truffle. Tuber lannaense, from Thailand. Mycol. Prog. 2016, 15, 827–834. [Google Scholar]
- Suwannarach, N.; Kumla, J.; Lumyong, S. Tuber magnatum in Thailand, a first report from Asia. Mycotaxon 2017, 132, 635–642. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of a problematic alien invasive species-the American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Nagler, M.; Insam, H.; Pietramellara, G.; Ascher-Jenull, J. Extracellular DNA in natural environments: Features, relevance and applications. Appl. Microbiol. Biotechnol. 2018, 102, 6343–6356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—an emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Hajibabaei, M.; Rieseberg, L.H. Environmental DNA. Mol. Ecol. 2012, 21, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Epp, L.S.; Boessenkool, S.; Bellemain, E.P.; Haile, J.; Esposito, A.; Riaz, T.; Erseus, C.; Gusarov, V.I.; Edwards, M.E.; Johnsen, A. New environmental metabarcodes for analysing soil DNA: Potential for studying past and present ecosystems. Mol. Ecol. 2012, 21, 1821–1833. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Soliman, T.; Yang, S.Y.; Yamazaki, T.; Jenke-Kodama, H. Profiling soil microbial communities with next-generation sequencing: The influence of DNA kit selection and technician technical expertise. PeerJ. 2017, 5, e4178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deiner, K.; Walser, J.; Machler, E.; Altermatt, F. Choice of capture and extraction methods affect detection of freshwater biodiversity from environmental DNA. Biol. Conserv. 2015, 183, 53–63. [Google Scholar] [CrossRef]
- Uroz, S.; Oger, P.; Tisserand, E.; Cebron, A.; Turpault, M.P.; Buee, M.; de Boer, W.; Leveau, J.H.; Frey-Klett, P. Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities. Sci. Rep. 2016, 6, 27756. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, M.; Peng, M.; Sui, X.; Li, W.; Sun, G. Variations in soil functional fungal community structure associated with pure and mixed plantations in typical temperate forests of China. Front. Microbiol. 2019, 10, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedersoo, L.; Bahram, M.; Cajthaml, T.; Polme, S.; Hiiesualu, I.; Anslan, S.; Harend, H.; Buegger, F.; Pritsch, K.; Koricheva, J.; et al. Tree diversity and species identity effects on soil fungi, protists and animals are context dependent. ISME J. 2016, 10, 346–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landinez-Torres, A.; Panelli, S.; Picco, A.M.; Comandatore, F.; Tosi, S.; Capelli, E. A meta-barcoding analysis of soil mycobiota of the upper Andean Colombian agro-environment. Sci. Rep. 2019, 9, 10085. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers--a user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iotti, M.; Amicucci, A.; Bonito, G.; Bonuso, E.; Stocchi, V.; Zambonelli, A. Selection of a set of specific primers for the identification of Tuber rufum: A truffle species with high genetic variability. FEMS Microbiol. Lett. 2007, 277, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Sejalon-Delmas, N.; Roux, C.; Martins, M.; Kulifaj, M.; Becard, G.; Dargent. Molecular tools for the identification of Tuber melanosporum in agroindustry. J. Agric. Food Chem. 2000, 48, 2608–2613. [Google Scholar] [CrossRef] [PubMed]
- Gryndler, M.; Hršelová, H.; Soukupová, L.; Streiblová, E.; Valda, S.; Borovička, J.; Gryndlerová, H.; Gažo, J.; Miko, M. Detection of summer truffle (Tuber aestivum Vittad.) in ectomycorrhizae and in soil using specific primers. FEMS Microbiol. Lett. 2011, 318, 84–91. [Google Scholar] [CrossRef]
- Herder, J.; Valentin, A.; Bellemain, E.; Dejean, T.; van Delft, J.; Thomsen, P.F.; Taberlet, P. Environmental DNA a Review of the Possible Applications for the Detection of (Invasive) Species; Stichting RAVON: Nijmegen, The Netherlands, 2014. [Google Scholar]
- Horisawa, S.; Sakuma, Y.; Doi, S. Qualitative and quantitative PCR methods using species-specific primer for detection and identification of wood rot fungi. J. Wood Sci. 2009, 55, 133–138. [Google Scholar] [CrossRef]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Mbareche, H.; Veillette, M.; Bilodeau, G.; Duchaine, C. Comparison of the performance of ITS1 and ITS2 as barcodes in amplicon-based sequencing of bioaerosols. PeerJ 2020, 8, e8523. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.C.; Liu, C.; Huang, L.; Bengtsson-Palme, J.; Chen, H.; Zhang, J.H.; Cai, D.; Li, J.Q. ITS1: A DNA barcode better than ITS2 in eukaryotes? Mol. Ecol. Resour. 2015, 15, 573–586. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Kristiansson, E.; Ryberg, M.; Hallenberg, N.; Larsson, K.-H. Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol. Bioinform. 2008, 4, EBO–S653. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods. 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.H.; Zheng, L.Y.; Liu, C.Y.; Wang, Y.; Li, L.; Zhao, Y.C.; Zhang, X.L.; Yang, M.; Xiong, H.K.; Qing, Y.; et al.; et al. Two new truffles species, Tuber alboumbilicum and Tuber pseudobrumale from China. Mycol. Prog. 2014, 13, 1157–1163. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and modelchoice across a large model space. Sys. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or bootstrap? A simulation study comparing the performance of Bayesian Markov Chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonito, G.; Smith, M.E.; Nowak, M.; Healy, R.A.; Guevara, G.; Cázares, E.; Kinoshita, A.; Nouhra, E.R.; Domínguez, L.S.; Tedersoo, L.; et al. Historical biogeography and diversification of truffles in the Tuberaceae and their newly identified southern hemisphere sister lineage. PLoS ONE 2013, 8, e52765. [Google Scholar] [CrossRef]
- Polemis, E.; Konstantinidis, G.; Fryssouli, V.; Slavova, M.; Tsampazis, T.; Nakkas, V.; Assyov, B.; Kaounas, V.; Zervakis, G.I. Tuber pulchrosporum sp. nov., a black truffle of the Aestivum clade (Tuberaceae, Pezizales) from the Balkan peninsula. MycoKeys. 2019, 20, 35–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wubet, T.; Christ, S.; Schöning, I.; Boch, S.; Gawlich, M.; Schnabel, B.; Fischer, M.; Buscot, F. Differences in soil fungal communities between European beech (Fagus sylvatica L.) dominated forests are related to soil and understory vegetation. PLoS ONE 2012, 7, e47500. [Google Scholar] [CrossRef] [PubMed]
- Coince, A.; Caël, O.; Bach, C.; Lengellé, J.; Cruaud, C.; Gavory, F. Below-ground fine-scale distribution and soil versus fine root detection of fungal and soil oomycete communities in a French beech forest. Fungal Ecol. 2013, 6, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Osona, T. Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, T.; Guo, T.; Yao, Y.; Wang, R.; Chai, B. Seasonal microbial community characteristic and its driving factors in a copper tailings dam in the Chinese Loess Plateau. Front. Microbiol. 2020, 11, 1574. [Google Scholar] [CrossRef] [PubMed]
- Nagati, M.; Roy, M.; Manzi, S.; Richard, F.; Desrocjers, A.; Gardes, M.; Bergeron, Y. Impact of local forest composition on soil fungal communities in a mixed boreal forest. Plant Soil 2018, 432, 345–357. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun 2019, 10, 2369. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, N.; Schneider, D.; Sahner, J.; Ballauff, J.; Edy, N.; Barus, H.; Irawan, B.; Wilarso, S.; Qaim, M.; Daniel, R.; et al. Intensive tropical land use massively shifts soil fungal communities. Sci. Rep. 2019, 9, 3403. [Google Scholar] [CrossRef] [Green Version]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Terhonen, E.; Kovalchuk, A.; Tuovila, H.; Chen, H.; Oghenekaro, A.O.; Heinonsala, J.; Kohler, A.; Kasanen, R.; Vasander, H.; et al. Dominant tree species and soil type affect the fungal community structure in a boreal peatland forest. Appl. Environ. Microbiol. 2016, 82, 2632–2643. [Google Scholar] [CrossRef] [Green Version]
- Männistö, M.; Vuosku, J.; Stark, S.; Saravesi, K.; Suokas, M.; Markkola, A.; Martz, F.; Rautio, P. Bacterial and fungal communities in boreal forest soil are insensitive to changes in snow cover conditions. FEMS Microbiol. Ecol. 2018, 94, 1–14. [Google Scholar]
- Shi, L.; Dossa, G.G.O.; Paudel, E.; Zang, H.; Xu, J.; Harrison, R.D. Changes in fungal communities across a forest disturbance gradient. Appl. Environ. Microbiol. 2019, 85, e00080-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuske, S.J.; Anslan, S.; Tedersoo, L.; Congdon, B.C.; Abell, S.E. Ectomycorrhizal fungal communities are dominated by mammalian dispersed truffle-like taxa in north-east Australian woodlands. Mycrorrhiza 2019, 29, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuong, T.M.D.; Zeng, J.Y.; Man, X.L. Soil fungal and bacterial communities in southern boreal forests of the Greater Khingan Mountains and their relationship with soil properties. Sci. Rep. 2020, 10, 22025. [Google Scholar] [CrossRef]
- Hambleton, S.; Sigler, L. Meliniomyces, a new anamorph genus for root-associated fungi with phylogenetic affinities to Rhizoscyphus ericae (≡ Hymenoscyphus ericae), Leotiomycetes. Stud. Mycol. 2005, 53, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Villarreal-Ruiz, L.; Neri-Luna, C.; Anderson, I.C.; Alexander, I.J. In vitro interactions between ectomycorrhizal fungi and ericaceous plants. Symbiosis 2012, 56, 67–75. [Google Scholar] [CrossRef]
- Gómez-Hernández, M.; Williams-Linera, G.; Guevara, R.; Lodge, D.J. Patterns of macromycete community assemblage along an elevation gradient: Options for fungal gradient and metacommunity analyse. Biodivers. Conserv. 2012, 21, 2247. [Google Scholar] [CrossRef]
- Cuadros-Orellana, S.; Leite, L.R.; Smith, A.; Medeiros, J.D.; Badotti, F.; Fonseca, P.L.C.; Vaz, A.B.; Oliveira, G.; Goes-Neta, G. Assessment of fungal diversity in the environment using Metagenomics: A decade in review. Fungal Genom. Biol. 2013, 3, 2. [Google Scholar]
- Wozniak, M.; Gradziel, J.; Galazka, A.; Frac, M. Metagaenomic analysis of bacteria and fungi community composition associated with Paulownia elongate × Paulownia fortune. BioResources 2019, 14, 8511–8529. [Google Scholar] [CrossRef]
- Bai, L.; Sun, H.; Zhang, X.; Cai, B. Next-generation sequencing of root fungal communities in continuous cropping soybean. Chil. J. Agric. Res. 2018, 78, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Sommermann, L.; Geistlinger, J.; Wibberg, D.; Deubel, A.; Zwanzig, J.; Babin, D.; Schluter, A.; Schellenbern, I. Fungal community profiles in agricultural soils of a long-term field trial under different tillage, fertilization and crop rotation conditions analyzed by high-throughput ITS-amplicon sequencing. PLoS One 2018, 13, e0195345. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Maunoury, L.; Deveau, A.; Moreno, M.; Todesco, F.; Belmondo, S.; Murat, C.; Courty, P.E.; Jąkalski, M.; Selosse, M.A. Two ectomycorrhizal truffles, Tuber melanosporum and T. aestivum, endophytically colonise roots of non-ectomycorrhizal plants in natural environments. New Phytol. 2020, 225, 2542–2556. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Ye, L.; Huang, Y.; Kang, Z.; Zhang, B. Colonization by Tuber melanosporum and Tuber indicum affects the growth of Pinus armandii and phoD alkaline phosphatase encoding bacterial community in the rhizosphere. Microbiol. Res. 2020, 239, 126520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ye, L.; Kang, Z.; Zou, J.; Zhang, X.; Li, X. Mycorrhization of Quercus acutissima with Chinese black truffle significantly altered the host physiology and root-associated microbiomes. Peer J. 2019, 7, e6421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Li, X.; Li, Q.; Wu, H.; Xiong, C.; Geng, Q.; Sun, H.; Sun, Q. Soil microbial communities of three major Chinese truffles in southwest China. Can. J. Microbiol. 2016, 62, 970–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaco, P.; Toumi, M.; Sferra, G.; Toth, E.; Naclerio, G.; Bucci, A. The bacterial communities of Tuber aestivum: Preliminary investigations in Molise region, southern Italy. Ann. Microbiol. 2010, 70, 37. [Google Scholar] [CrossRef]
- Splivallo, R.; Vahdatzadeh, M.; Maciá-Vicente, J.G.; Molinier, V.; Peter, M.; Egli, S.; Uroz, S.; Paolocci, F.; Deveau, A. Orchard Conditions and fruiting body characteristics drive the microbiome of the black truffle Tuber aestivum. Front. Microbiol. 2019, 10, 1437. [Google Scholar] [CrossRef] [PubMed]
- Antony-Babu, S.; Deveau, A.; Nostrand, J.D.V.; Zhou, J.; Tacon, F.L.; Robin, C.; Frey-Klett, P.; Uroz, S. Black truffle-associated bacterial communities during the development and maturation of Tuber melanosporum ascocarps and putative functional roles. Environ. Microbol. 2014, 16, 2831–2847. [Google Scholar] [CrossRef]
- Deveau, A.; Antony-Babu, S.; Tacon, L.F.; Robin, C.; Frey-Klett, P.; Uroz, S. Temporal changes of bacterial communities in the Tuber melanosporum ectomycorrhizosphere during ascocarp development. Mycorrhiza 2016, 26, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Li, Q.; Fu, Y.; Sourzat, P.; Tan, H.; Zou, J.; Zhang, B.; Li, X. Host species effects on bacterial communities associated with the ascocarps of Tuber species from the Sichuan Province in southwest China. Mycol. Prog. 2018, 17, 833–840. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. * | Relative Abundant (%) † | OTU Number | Observation of Fruiting Body | |||||
|---|---|---|---|---|---|---|---|---|
| 105 | 162 | 187 | 447 | 530 | 720 | |||
| B1 | 0.02 | – | + | – | – | – | – | + |
| B2 * | 0.08 | – | + | – | – | – | – | + |
| B3 | 0.04 | – | + | – | – | – | + | – |
| B4 | 0.10 | – | + | – | – | – | – | + |
| B5 | 0.05 | – | + | – | – | – | – | + |
| B6 | 0.10 | – | + | – | – | – | – | – |
| B7 | 0.20 | + | + | – | – | – | – | + |
| B8 | 0.60 | – | – | – | – | – | + | – |
| B9 | 0.07 | + | – | – | – | – | – | – |
| B10 | 1.00 | – | + | – | – | – | – | + |
| B11 | 0.03 | – | + | – | – | – | – | – |
| B12 | 0.20 | – | + | – | + | – | – | + |
| B13 | 0.06 | – | + | – | – | – | – | + |
| B14 | 0.30 | – | + | – | – | – | – | + |
| B15 | 0.03 | – | + | – | – | – | – | + |
| B16 | 0.02 | – | + | – | – | – | – | – |
| B17 | 0.02 | + | – | – | – | – | – | – |
| B18 | 0.00 | – | – | – | – | – | – | – |
| B19 | 0.00 | – | – | – | – | – | – | – |
| B20 | 4.00 | – | + | – | + | – | – | – |
| B21 | 0.02 | – | – | – | – | – | + | – |
| B22 | 0.09 | – | + | – | – | – | – | + |
| B23 | 0.20 | – | + | – | – | – | – | – |
| B24 | 0.07 | – | – | – | – | – | + | – |
| B25 | 0.00 | – | – | – | – | – | – | – |
| B26 | 0.20 | – | + | – | – | – | – | + |
| B27 * | 0.90 | – | + | + | – | – | – | + |
| B28 | 0.03 | + | – | – | – | – | – | – |
| B29 | 0.04 | – | + | – | – | – | – | + |
| B30 | 0.00 | – | – | – | – | – | – | – |
| B31 | 0.03 | – | + | – | – | – | – | + |
| B32 | 0.10 | – | + | + | – | – | – | + |
| B33 | 0.03 | – | + | – | – | – | – | + |
| B34 | 0.02 | – | – | – | + | – | – | – |
| B35 | 0.00 | – | – | – | – | – | – | – |
| B36 | 0.80 | – | + | + | – | – | – | + |
| CP1 | 0.01 | – | – | + | – | – | – | – |
| CP2 * | 0.20 | – | – | + | – | + | – | – |
| Total found | 4 | 24 | 5 | 3 | 1 | 4 | 18 | |
| OTU Number | Length (bp) | GenBank Accession Number | Closeted Species/Accession Number | Similarity (%) |
|---|---|---|---|---|
| 105 | 182 | MW330007 | Tuber thailandicum CMU-MTUF001/KP196331 | 97.52 |
| 162 | 238 | MW326971 | Tuber thailandicum CMU-MTUF001/KP196332 | 100 |
| 187 | 228 | MW330257 | Tuber lanaense CMU-MTUF007/KT758731 | 100 |
| 447 | 211 | MW327594 | Tuber bomiense SKM101/KC517480 | 100 |
| 530 | 164 | MW326781 | Tuber magnatum Tmag_Ca_09/MG992596 | 100 |
| 720 | 168 | MW326083 | Tuber sp. isolate JT9769/HM485407 | 100 |
| Location | Macromorphological Characteristics | Micromorphological Characteristics | ||||||
|---|---|---|---|---|---|---|---|---|
| Ascomata | Hair-like Structure in Peridium Surface | Number of Ascospore/Ascus | Ascospores | |||||
| Size (cm in Diam) | Color | Gleba | Shape | Ornamentation | Size (μm) | |||
| B1 | 1.5–3.5 | White | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 23–40 × 22–35 |
| B2 * | 2.2–4.3 | White to pale yellow | Brown | + | 1–4 | Globose to broadly ellipsoid | Alveolate-reticulate | 25–45 × 22–40 |
| B4 | 2.0 | White | Brown | + | 1–4 | Subglobose to broadly ellipsoid | Alveolate-reticulate | 20–35 × 18–28 |
| B5 | 1.0–3.0 | Pale yellow to light brown | Brown to dark brown | + | 1–4 | Subglobose to ellipsoid | Alveolate-reticulate | 25–55 × 22–48 |
| B7 | 2.5–3.0 | White to pale yellow | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 28–40 × 25–38 |
| B10 | 1.5–3.5 | White to pale yellow | Brown | + | 1–4 | Subglobose | Alveolate-reticulate | 25–60 × 22–55 |
| B12 | 2.5–4.5 | Pale yellow to light brown | Dark brown | + | 1–4 | Subglobose to broadly ellipsoid | Alveolate-reticulate | 25–43 × 22–35 |
| B13 | 2.0 | Pale yellow | Brown | + | 1–4 | Globose to broadly ellipsoid | Alveolate-reticulate | 25–45 × 25–35 |
| B14 | 1.0–3.2 | White to pale yellow | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 26–42 × 25–38 |
| B15 | 2.5 | White | Brown | + | 1–4 | Subglobose | Alveolate-reticulate | 20–50 × 18–47 |
| B22 | 2.5–4.0 | Pale yellow to light brown | Dark brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 20–65 × 18–62 |
| B26 | 1.4–3.8 | White | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 27–50 × 25–48 |
| B27 * | 1.0–3.2 | White to pale yellow | Dark brown | + | 1–4 | Globose to broadly ellipsoid | Alveolate-reticulate | 25–43 × 25–35 |
| B29 | 3.5 | White | Brown | + | 1–4 | Gubglobose to ellipsoid | Alveolate-reticulate | 25–50 × 22–40 |
| B31 | 2.0–4.1 | White to pale yellow | Brown | + | 1–5 | Globose to broadly ellipsoid | Alveolate-reticulate | 22–45 × 20–42 |
| B32 | 2.0–3.0 | Pale yellow to light brown | Brown to dark brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 25–45 × 20–35 |
| B33 | 2.4–3.5 | White to pale yellow | Brown | + | 1–4 | Subglobose to ellipsoid | Alveolate-reticulate | 20–55 × 15–50 |
| B36 | 1.5–3.0 | White, pale yellow to light brown | Brown to dark brown | + | 1–4 | Subglobose to ellipsoid | Alveolate-reticulate | 30–50 × 22–45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suwannarach, N.; Kumla, J.; In-on, A.; Lumyong, S. Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand. J. Fungi 2021, 7, 293. https://doi.org/10.3390/jof7040293
Suwannarach N, Kumla J, In-on A, Lumyong S. Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand. Journal of Fungi. 2021; 7(4):293. https://doi.org/10.3390/jof7040293
Chicago/Turabian StyleSuwannarach, Nakarin, Jaturong Kumla, Ammarin In-on, and Saisamorn Lumyong. 2021. "Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand" Journal of Fungi 7, no. 4: 293. https://doi.org/10.3390/jof7040293
APA StyleSuwannarach, N., Kumla, J., In-on, A., & Lumyong, S. (2021). Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand. Journal of Fungi, 7(4), 293. https://doi.org/10.3390/jof7040293