
Genomic Analyses of Penicillium Species Have Revealed Patulin and Citrinin Gene Clusters and Novel Loci Involved in Oxylipin Production

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Preparation and Sequencing
2.2. Bioinformatic Analyses of Penicillium spp.
2.3. Exposure of Drosophila 3rd Instar Larvae to VOCs Emitted by Seven Penicillium Species
3. Results
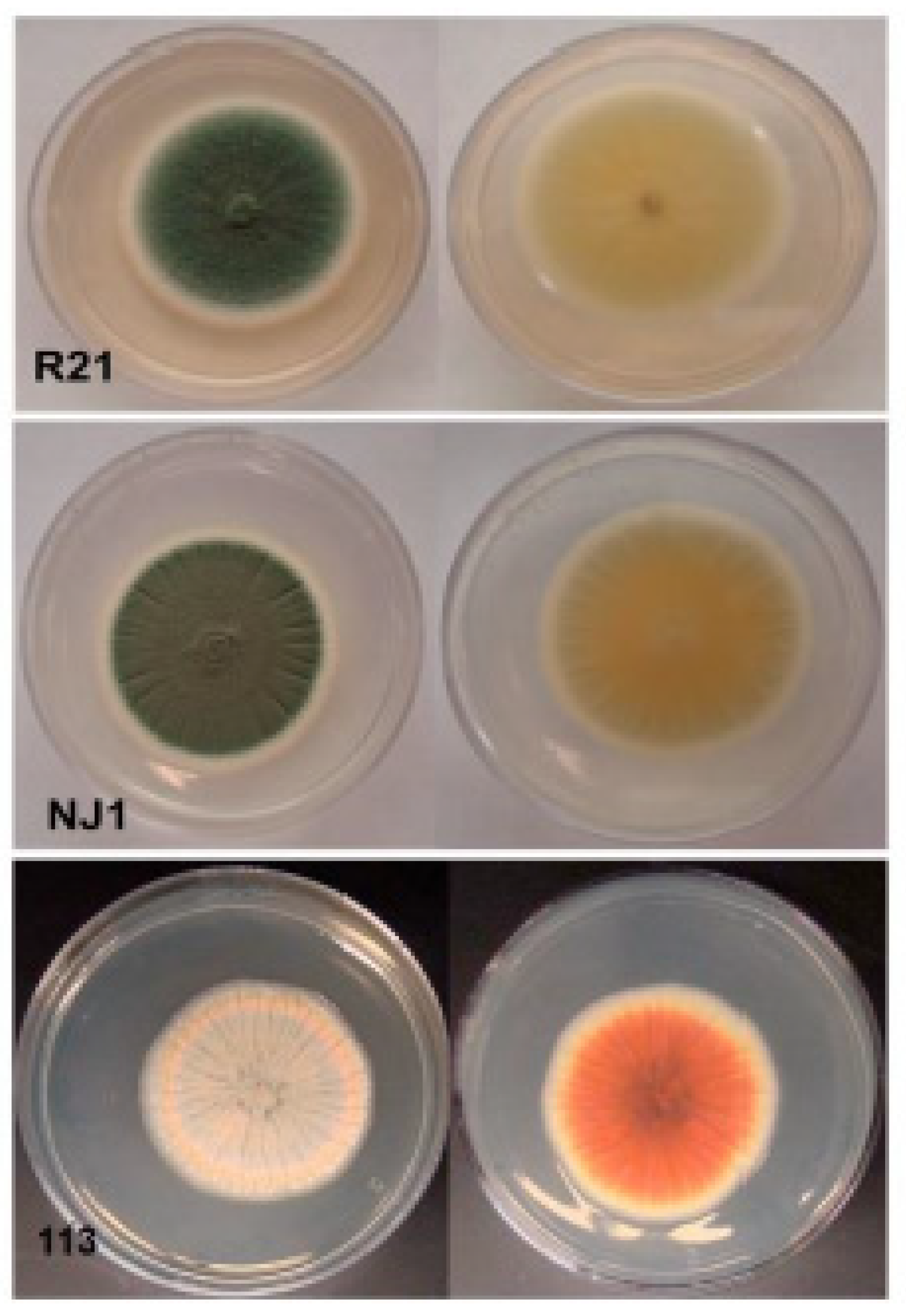
3.1. Fungal Isolation, Sequencing Data Statistics, Genome Assembly, and Gene Prediction
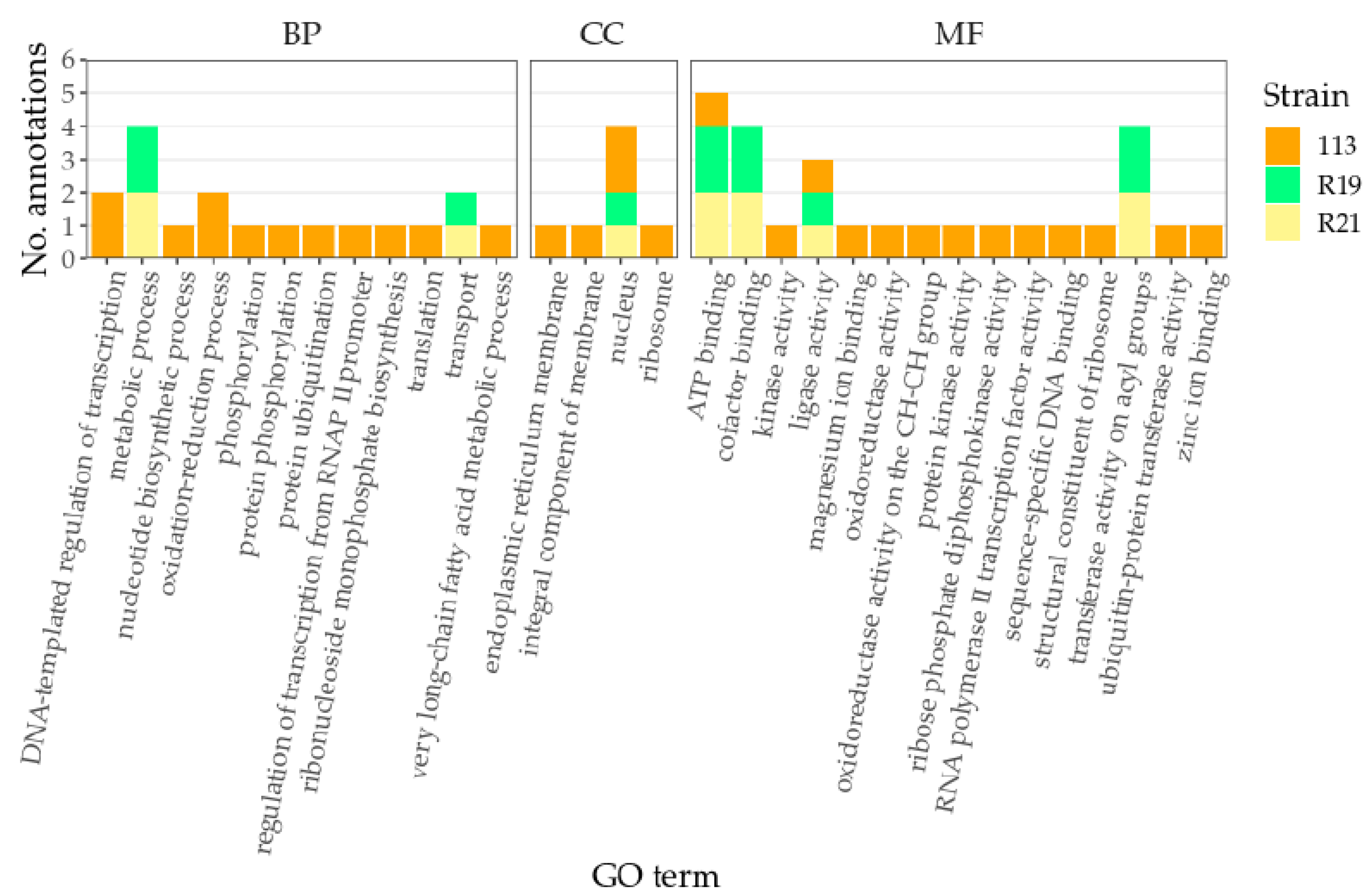
3.2. COG Analyses and GO Annotation
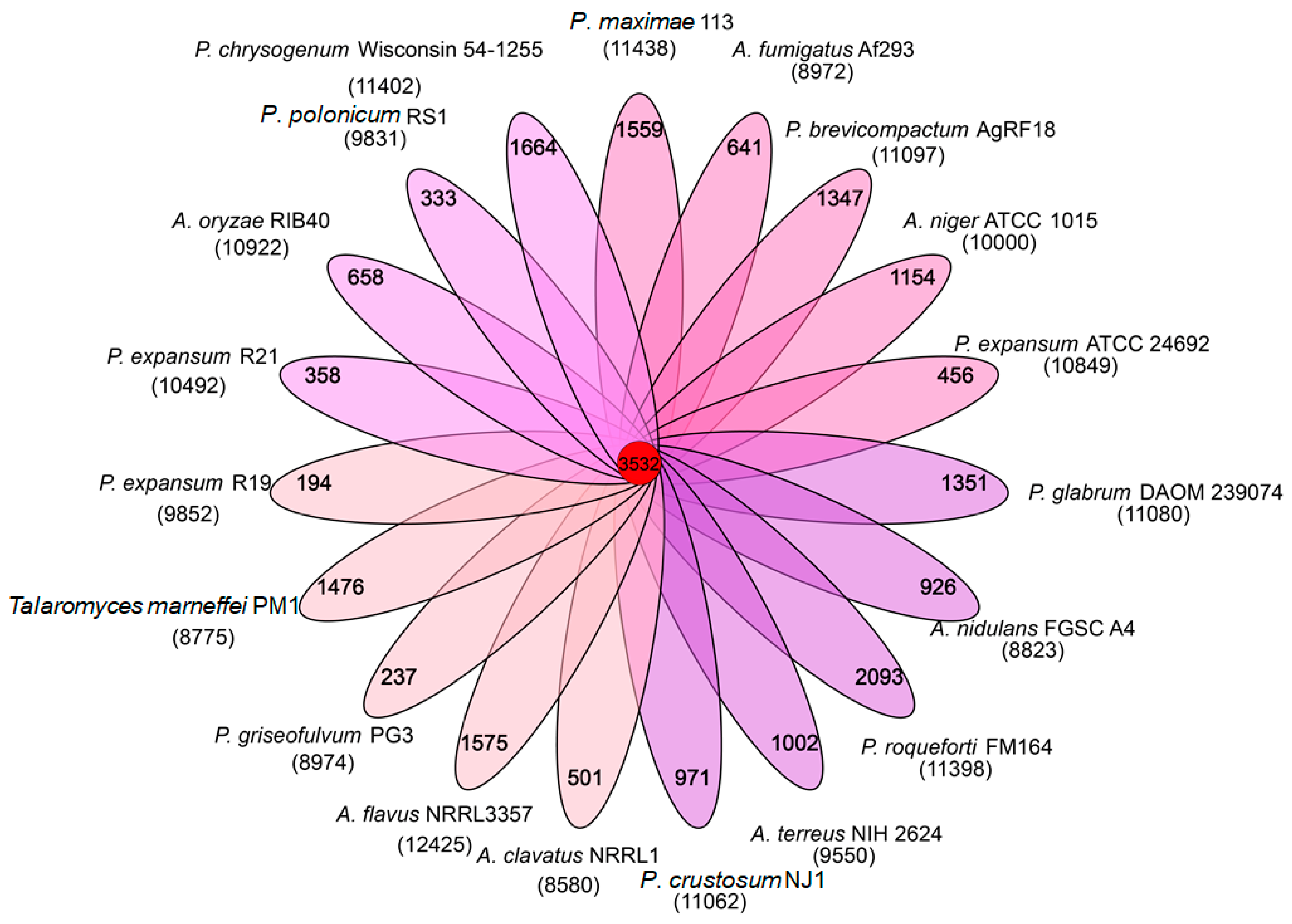
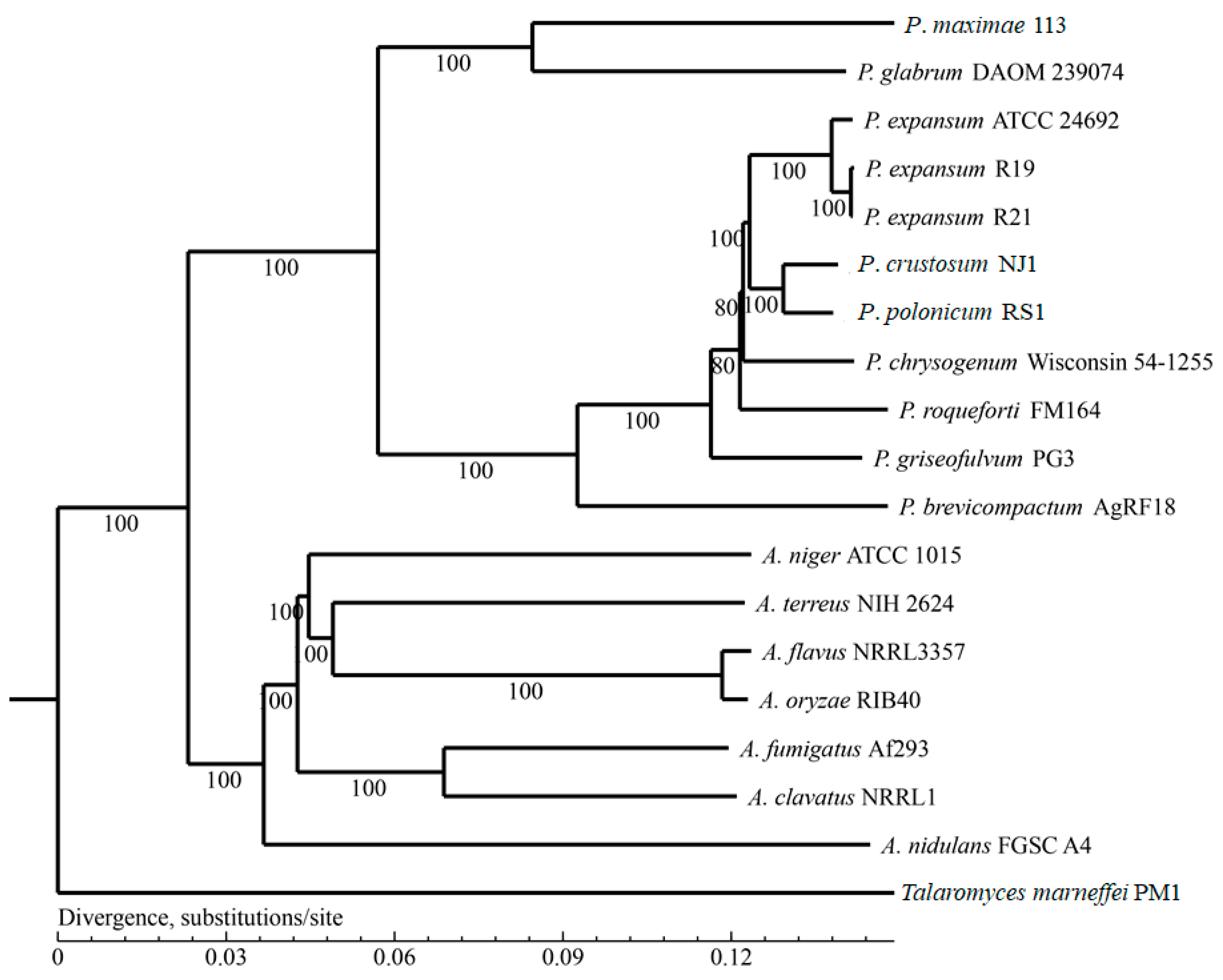
3.3. Homologous Genes and Phylogenetic Analysis
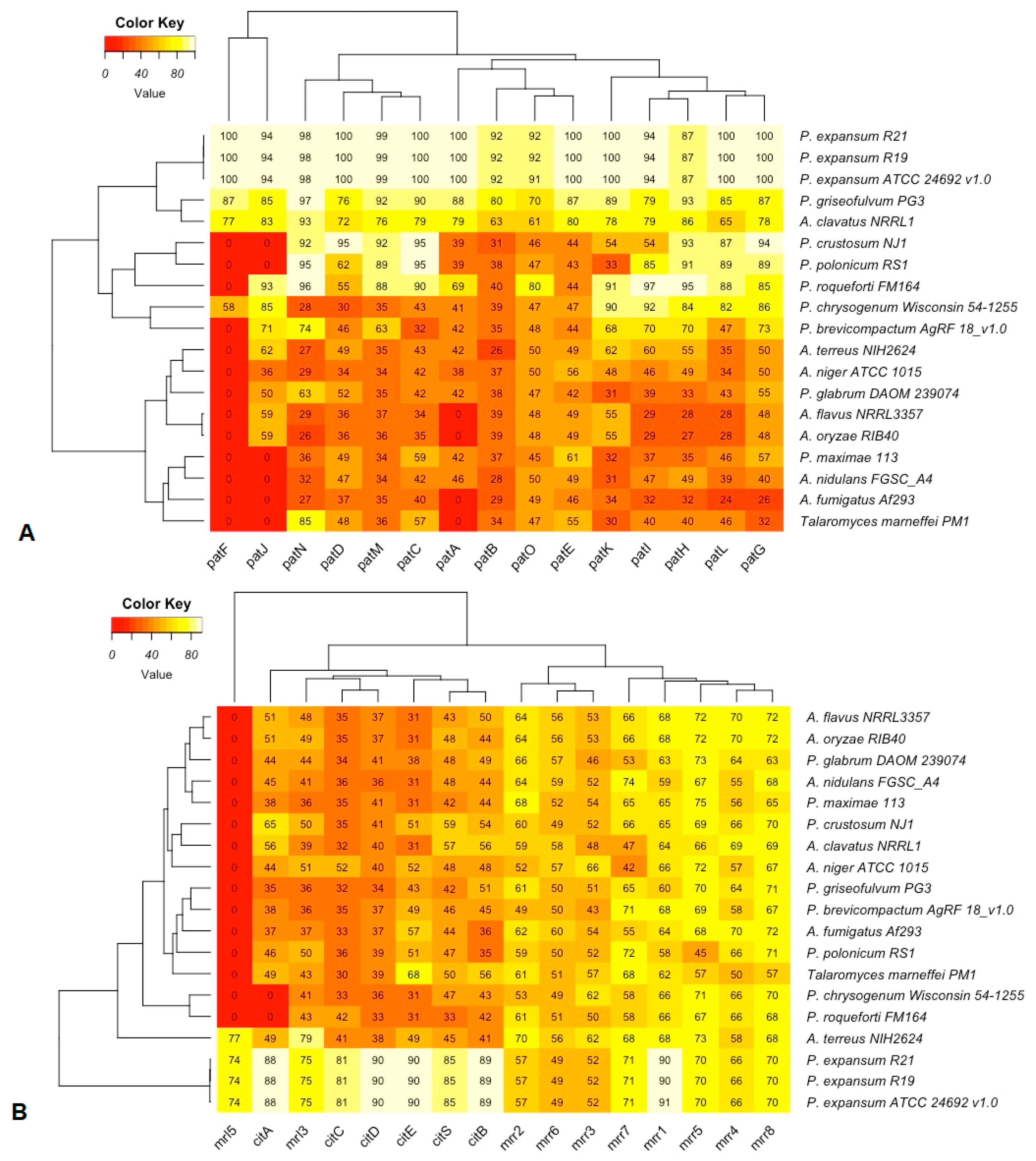
3.4. Patulin and Citrinin Gene Clusters
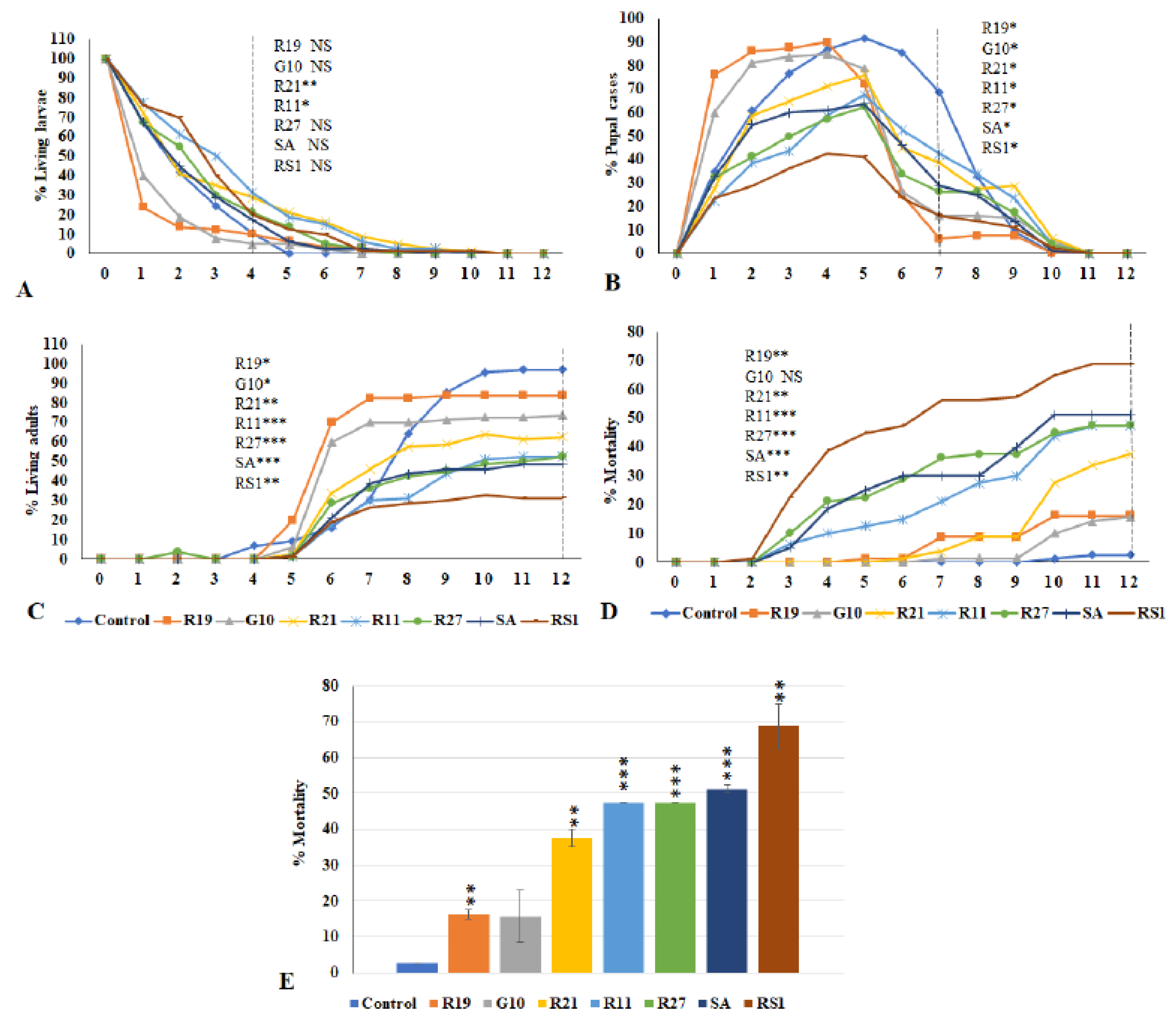
3.5. Effects of VOCs Emitted from Penicillium Species on the Third Instar Drosophila Larvae
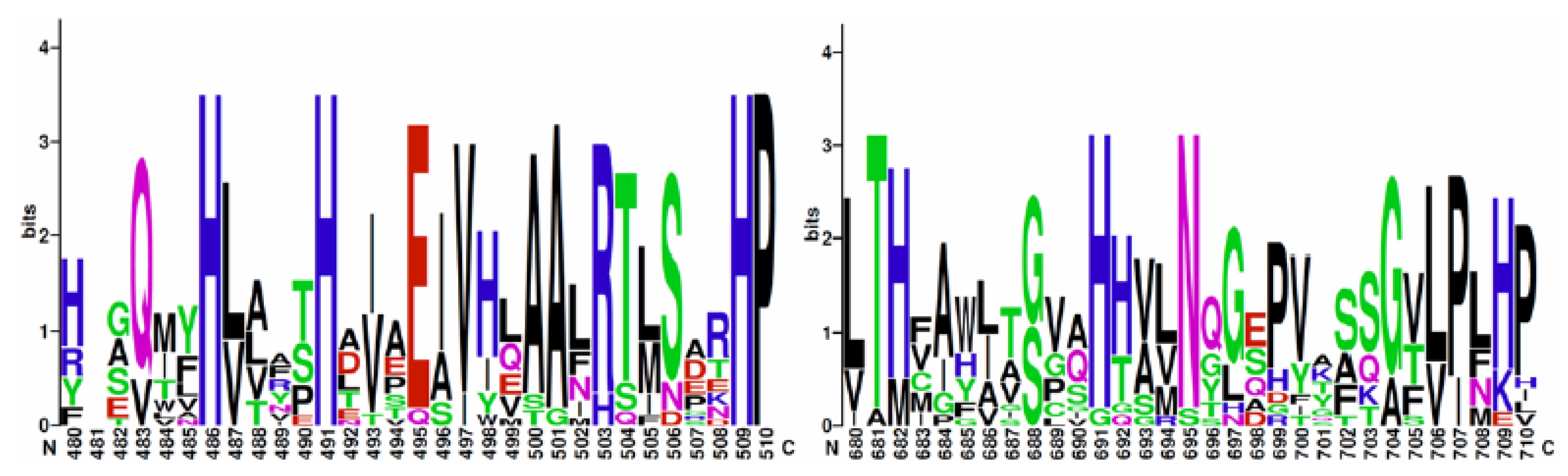
3.6. Identification of Putative Lipoxygenase Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Visagie, C.; Houbraken, J.; Frisvad, J.C.; Hong, S.-B.; Klaassen, C.; Perrone, G.; Seifert, K.; Varga, J.; Yaguchi, T.; Samson, R. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.K. New and Future Developments in Microbial Biotechnology and Bioengineering: Aspergillus System Properties and Applications; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Houbraken, J.; Kocsubé, S.; Visagie, C.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Shelton, B.G.; Kirkland, K.H.; Flanders, W.D.; Morris, G.K. Profiles of airborne fungi in buildings and outdoor environments in the United States. Appl. Environ. Microbiol. 2002, 68, 1743–1753. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, M.A.; Albang, R.; Albermann, K.; Badger, J.H.; Daran, J.M.; Driessen, A.J.; Garcia-Estrada, C.; Fedorova, N.D.; Harris, D.M.; Heijne, W.H.; et al. Genome sequencing and analysis of the filamentous fungus Penicillium chrysogenum. Nat. Biotechnol. 2008, 26, 1161–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.K.; Cooper, E.A.; Bowen, J.E. Meta-Analysis: Griseofulvin efficacy in the treatment of tinea capitis. J. Drugs Dermatol. 2008, 7, 369–372. [Google Scholar] [PubMed]
- Molimard, P.; Spinnler, H. Review: Compounds involved in the flavor of surface mold-ripened cheeses: Origins and properties. J. Dairy Sci. 1996, 79, 169–184. [Google Scholar] [CrossRef]
- Lacey, J.; Crook, B. Fungal and actinomycete spores as pollutants of the workplace and occupational allergens. Ann. Occup. Hyg. 1988, 32, 515–533. [Google Scholar]
- Frisvad, J.C.; Samson, R.A. Penicillium subgenus Penicillium. A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 1, 1–174. [Google Scholar]
- Chalutz, E.; Wilson, C. Postharvest biocontrol of green and blue mold and sour rot of citrus fruit by Debaryomyces hansenii. Plant Dis. 1990, 74, 134–137. [Google Scholar] [CrossRef]
- Eckert, J.; Sievert, J.; Ratnayake, M. Reduction of imazalil effectiveness against citrus green mold in California packinghouses by resistant biotypes of Penicillium digitatum. Plant Dis. 1994, 78, 971–974. [Google Scholar] [CrossRef]
- Li, B.; Zong, Y.; Du, Z.; Chen, Y.; Zhang, Z.; Qin, G.; Zhao, W.; Tian, S. Genomic characterization reveals insights into patulin biosynthesis and pathogenicity in Penicillium species. Mol. Plant-Microbe Interact. 2015, 28, 635–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballester, A.-R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant-Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Yin, G.; Inamda, A.A.; Luo, J.; Zhang, N.; Yang, I.; Buckley, B.; Bennett, J.W. Volatile organic compounds emitted by filamentous fungi isolated from flooded homes after Hurricane Sandy show toxicity in a Drosophila bioassay. Indoor Air 2017, 27, 518–528. [Google Scholar] [CrossRef]
- Rao, C.Y.; Riggs, M.A.; Chew, G.L.; Muilenberg, M.L.; Thorne, P.S.; Van Sickle, D.; Dunn, K.H.; Brown, C. Characterization of airborne molds, endotoxins, and glucans in homes in New Orleans after Hurricanes Katrina and Rita. Appl. Environ. Microbiol. 2007, 73, 1630–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacho, R.A.; Tang, Y.; Chooi, Y.-H. Next-Generation sequencing approach for connecting secondary metabolites to biosynthetic gene clusters in fungi. Front. Microbiol. 2015, 5, 774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcet-Houben, M.; Ballester, A.-R.; de la Fuente, B.; Harries, E.; Marcos, J.F.; González-Candelas, L.; Gabaldón, T. Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genom. 2012, 13, 646. [Google Scholar] [CrossRef] [Green Version]
- Julca, I.; Droby, S.; Sela, N.; Marcet-Houben, M.; Gabaldón, T. Contrasting genomic diversity in two closely related postharvest pathogens: Penicillium digitatum and Penicillium expansum. Genome Biol. Evol. 2016, 8, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Jurick, W.M.; Cao, H.; Yin, Y.; Gaskins, V.L.; Losada, L.; Zafar, N.; Kim, M.; Bennett, J.W.; Nierman, W.C. Draft genome sequence of Penicillium expansum strain R19, which causes postharvest decay of apple fruit. Genome Announc. 2014, 2, e00635-14. [Google Scholar] [CrossRef] [Green Version]
- Eldarov, M.A.; Mardanov, A.V.; Beletsky, A.V.; Dzhavakhiya, V.V.; Ravin, N.V.; Skryabin, K.G. Complete mitochondrial genome of compactin-producing fungus Penicillium solitum and comparative analysis of Trichocomaceae mitochondrial genomes. FEMS Microbiol. Lett. 2012, 329, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, K.; Ropars, J.; Renault, P.; Dupont, J.; Gouzy, J.; Branca, A.; Abraham, A.-L.; Ceppi, M.; Conseiller, E.; Debuchy, R. Multiple recent horizontal transfers of a large genomic region in cheese making fungi. Nat. Commun. 2014, 5, 2876. [Google Scholar] [CrossRef]
- Banani, H.; Marcet-Houben, M.; Ballester, A.-R.; Abbruscato, P.; González-Candelas, L.; Gabaldón, T.; Spadaro, D. Genome sequencing and secondary metabolism of the postharvest pathogen Penicillium griseofulvum. BMC Genom. 2016, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.; Lau, S.K.; Liu, B.; Cai, J.J.; Chong, K.T.; Tse, H.; Kao, R.Y.; Chan, C.-M.; Chow, W.-N.; Yuen, K.-Y. Draft genome sequence of Penicillium marneffei strain PM1. Eukaryot. Cell 2011, 10, 1740–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ropars, J.; de La Vega, R.R.; López-Villavicencio, M.; Gouzy, J.; Dupont, J.; Swennen, D.; Dumas, E.; Giraud, T.; Branca, A. Diversity and mechanisms of genomic adaptation in Penicillium. In Aspergillus and Penicillium in the Post-Genomic Era; Caister Academic Press: Poole, UK, 2016. [Google Scholar]
- Sanderson, P.; Spotts, R. Postharvest decay of winter pear and apple fruit caused by species of Penicillium. Phytopathology 1995, 85, 103–110. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium. A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Sutton, T.B.; Aldwinckle, H.S.; Agnello, A.M.; Walgenbach, J.F. Compendium of Apple and Pear Diseases and Pests; American Phytopath Society: St. Paul, MN, USA, 2014. [Google Scholar]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [Green Version]
- HemaMoorthy, T.; Prakasam, V. First report of Penicillium expansum causing bulb rot of lilium in India. Am.-Eurasian J. Agric. Environ. Sci. 2013, 13, 293–295. [Google Scholar]
- Wang, C.; Ai, J.; Lv, H.; Qin, H.; Yang, Y.; Liu, Y.; Fan, S. First report of Penicillium expansum causing postharvest decay on stored kiwifruit (Actinidia arguta) in China. Plant Dis. 2015, 99, 1037. [Google Scholar] [CrossRef]
- Palou, L.; Montesinos-Herrero, C.; Taberner, V.; Vilella-Esplá, J. First report of Penicillium expansum causing postharvest blue mold of fresh date palm fruit (Phoenix dactylifera) in Spain. Plant Dis. 2013, 97, 846. [Google Scholar] [CrossRef]
- Pitt, J.; Spotts, R.; Holmes, R.; Cruickshank, R. Penicillium solitum revived, and its role as a pathogen of pomaceous fruit. Phytopathology 1991, 81, 1108–1112. [Google Scholar] [CrossRef]
- Amiri, A.; Bompeix, G. Diversity and population dynamics of Penicillium spp. on apples in pre-and postharvest environments: Consequences for decay development. Plant Pathol. 2005, 54, 74–81. [Google Scholar] [CrossRef]
- Andersen, B.; Thrane, U. Food-Borne fungi in fruit and cereals and their production of mycotoxins. In Advances in Food Mycology; Springer: Boston, MA, USA, 2006; pp. 137–152. [Google Scholar]
- Vico, I.; Gaskins, V.; Duduk, N.; Vasić, M.; Yu, J.; Peter, K.A.; Jurick, W.M. First report of Penicillium crustosum causing blue mold on stored apple fruit in Serbia. Plant Dis. 2014, 98, 1430. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens—The IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Keller, N.P.; Atoui, A.; El Khoury, A.; Lteif, R.; Oswald, I.P.; Puel, O. Secondary metabolism in Penicillium expansum: Emphasis on recent advances in patulin research. Crit. Rev. Food Sci. Nutr. 2018, 58, 2082–2098. [Google Scholar] [CrossRef] [PubMed]
- Luciano-Rosario, D.; Keller, N.P.; Jurick, W.M. Penicillium expansum: Biology, omics, and management tools for a global postharvest pathogen causing blue mould of pome fruit. Mol. Plant Pathol. 2020, 21, 1391–1404. [Google Scholar] [CrossRef]
- Bennett, J.W.; Inamdar, A.A. Are some fungal volatile organic compounds (VOCs) mycotoxins? Toxins 2015, 7, 3785–3804. [Google Scholar] [CrossRef] [Green Version]
- Almaliki, H.S.; Drazanin, E.; Sadovnik, R.; Yin, G.; Bennett, J.W. Eight carbon volatiles are more toxic to wild-type Drosophila melanogaster than to flies with blocked immune system mutations. Entomol. Exp. Appl. 2021. (accepted). [Google Scholar]
- Yin, G.; Padhi, S.; Lee, S.; Hung, R.; Zhao, G.; Bennett, J.W. Effects of three volatile oxylipins on colony development in two species of fungi and on Drosophila larval metamorphosis. Curr. Microbiol. 2015, 71, 347–356. [Google Scholar] [CrossRef]
- Yin, G.; Zhang, Y.; Hua, S.S.T.; Yu, J.; Bu, L.; Pennerman, K.K.; Huang, Q.; Guo, A.; Bennett, J.W. Genome sequencing and analysis of the postharvest fungus Penicillium expansum R21. Genome Announc. 2017, 5, e01516-16. [Google Scholar] [CrossRef] [Green Version]
- Yung, P.Y.; Grasso, L.L.; Mohidin, A.F.; Acerbi, E.; Hinks, J.; Seviour, T.; Marsili, E.; Lauro, F.M. Global transcriptomic responses of Escherichia coli K-12 to volatile organic compounds. Sci. Rep. 2016, 6, 19899. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Wu, G.; Jurick, W.M.; Gaskins, V.L.; Yin, Y.; Yin, G.; Bennett, J.W.; Shelton, D.R. Genome sequence of Penicillium solitum RS1, which causes postharvest apple decay. Genome Announc. 2016, 4, e00363-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, G.; Zhang, Y.; Pennerman, K.K.; Wu, G.; Hua, S.S.T.; Yu, J.; Jurick, W.M.; Guo, A.; Bennett, J.W. Characterization of blue mold Penicillium species isolated from stored fruits using multiple highly conserved loci. J. Fungi 2017, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.; Zhang, Y.; Pennerman, K.K.; Hua, S.S.T.; Yu, J.; Guo, A.; Liu, Z.; Bennett, J.W. Draft genome sequence of the fungus Penicillium solitum NJ1. Genome Announc. 2016, 4, e01176-16. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.; Zhang, Y.; Pennerman, K.K.; Hua, S.S.T.; Huang, Q.; Guo, A.; Liu, Z.; Bennett, J.W. Genome sequencing and analysis of the filamentous fungus Penicillium sclerotiorum 113, isolated after Hurricane Sandy. Genome Announc. 2016, 4, e01153-16. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.; Jurick, W.M., II; Zhao, G.; Bennett, J.W. New names for three Penicillium strains based on updated barcoding and phylogenetic analyses. Microbiol. Resour. Announc. 2021. (Under review). [Google Scholar]
- Yang, F.; Zhang, Y.; Huang, Q.; Yin, G.; Pennerman, K.K.; Yu, J.; Liu, Z.; Li, D.; Guo, A. Analysis of key genes of jasmonic acid mediated signal pathway for defense against insect damages by comparative transcriptome sequencing. Sci. Rep. 2015, 5, 16500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Bao, E.; Jiang, T.; Girke, T. AlignGraph: Algorithm for secondary de novo genome assembly guided by closely related references. Bioinformatics 2014, 30, i319–i328. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 2047-2217X-2041-2018. [Google Scholar] [CrossRef]
- Tempel, S. Using and understanding RepeatMasker. In Mobile Genetic Elements; Humana Press: Totowa, NJ, USA, 2012; pp. 29–51. [Google Scholar]
- Holt, C.; Yandell, M. MAKER2: An annotation pipeline and genome-database management tool for second-generation genome projects. BMC Bioinform. 2011, 12, 491. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.; Chen, Y.; Zhang, Y.; Xu, Y. H-BLAST: A fast protein sequence alignment toolkit on heterogeneous computers with GPUs. Bioinformatics 2017, 33, 1130–1138. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Medema, M.H.; Kottmann, R.; Lee, S.Y.; Weber, T. The antiSMASH database, a comprehensive database of microbial secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2016, 45, D555–D559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ter-Hovhannisyan, V.; Lomsadze, A.; Chernoff, Y.O.; Borodovsky, M. Gene prediction in novel fungal genomes using an ab initio algorithm with unsupervised training. Genome Res. 2008, 18, 1979–1990. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.; Pertea, M. GFF Utilities: GffRead and GffCompare. F1000Research 2020, 9. [Google Scholar] [CrossRef]
- Knight, R.D.; Freeland, S.J.; Landweber, L.F. Selection, history and chemistry: The three faces of the genetic code. Trends Biochem. Sci. 1999, 24, 241–247. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Lichtner, F.J.; Gaskins, V.L.; Cox, K.D.; Jurick, W.M. Global transcriptomic responses orchestrate difenoconazole resistance in Penicillium spp. causing blue mold of stored apple fruit. BMC Genom. 2020, 21, 574. [Google Scholar] [CrossRef]
- Pennerman, K.K.; Scarsella, J.B.; Yin, G.-H.; Hua, S.-S.T.; Hartman, T.G.; Bennett, J.W. Volatile 1-octen-3-ol increases patulin production by Penicillium expansum on a patulin-suppressing medium. Mycotoxin Res. 2019, 35, 329–340. [Google Scholar] [CrossRef]
- Jurick, W.M.; Peng, H.; Beard, H.S.; Garrett, W.M.; Lichtner, F.J.; Luciano-Rosario, D.; Macarisin, O.; Liu, Y.; Peter, K.A.; Gaskins, V.L. Blistering1 modulates Penicillium expansum virulence via vesicle-mediated protein secretion. Mol. Cell. Proteom. 2020, 19, 344–361. [Google Scholar] [CrossRef]
- Frisvad, J. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. World Mycotoxin J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Wennman, A.; Oliw, E.H.; Karkehabadi, S.; Chen, Y. Crystal structure of manganese lipoxygenase of the rice blast fungus Magnaporthe oryzae. J. Biol. Chem. 2016, 291, 8130–8139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Fierro, F.; Gutiérrez, S.; Diez, B.; Martín, J.F. Resolution of four large chromosomes in penicillin-producing filamentous fungi: The penicillin gene cluster is located on chromosome II (9.6 Mb) in Penicillium notatum and chromosome 1 (10.4 Mb) in Penicillium chrysogenum. Mol. Gen. Genet. MGG 1993, 241, 573–578. [Google Scholar] [CrossRef]
- Yuen, K.-y.; Pascal, G.; Wong, S.S.; Glaser, P.; Woo, P.C.; Kunst, F.; Cai, J.J.; Cheung, E.Y.; Médigue, C.; Danchin, A. Exploring the Penicillium marneffei genome. Arch. Microbiol. 2003, 179, 339–353. [Google Scholar] [CrossRef]
- Lee, H.; Gurtowski, J.; Yoo, S.; Nattestad, M.; Marcus, S.; Goodwin, S.; McCombie, W.R.; Schatz, M. Third-Generation sequencing and the future of genomics. BioRxiv 2016, 048603. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, A.; Au, K.F. PacBio sequencing and its applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negedu, A.; Atawodi, S.; Ameh, J.; Umoh, V.; Tanko, H. Economic and health perspectives of mycotoxins: A review. Cont. J. Biomed. Sci. 2011, 5, 5–26. [Google Scholar]
- Upadhaya, S.D.; Park, M.; Ha, J.-K. Mycotoxins and their biotransformation in the rumen: A review. Asian-Australas. J. Anim. Sci. 2010, 23, 1250–1260. [Google Scholar] [CrossRef]
- Perrone, G.; Susca, A. Penicillium species and their associated mycotoxins. In Mycotoxigenic Fungi; Humana Press: New York, NY, USA, 2017; pp. 107–119. [Google Scholar]
- Andersen, B.; Smedsgaard, J.; Frisvad, J.C. Penicillium expansum: Consistent production of patulin, chaetoglobosins, and other secondary metabolites in culture and their natural occurrence in fruit products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; El Khoury, R.; Snini, S.P.; Lippi, Y.; El Khoury, A.; Atoui, A.; Lteif, R.; Oswald, I.P.; Puel, O. Sequencing, physical organization and kinetic expression of the patulin biosynthetic gene cluster from Penicillium expansum. Int. J. Food Microbiol. 2014, 189, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, H.; Kitagawa, E.; Suzuki, Y.; Ueda, Y.; Ishizawa, Y.-h.; Nobumasa, H.; Kuboki, Y.; Hosoda, H.; Iwahashi, Y. Evaluation of toxicity of the mycotoxin citrinin using yeast ORF DNA microarray and Oligo DNA microarray. BMC Genom. 2007, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-P.; Xu, Y.; Huang, Z.-B. Isolation and characterization of the citrinin biosynthetic gene cluster from Monascus aurantiacus. Biotechnol. Lett. 2012, 34, 131–136. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cox, R.J. The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.; Chen, X.; Zheng, H.; Liao, D.; Yu, Z.; Wang, Z.; Chen, J. Genomic and chemical investigation of bioactive secondary metabolites from a marine-derived fungus Penicillium steckii P2648. Front. Microbiol. 2021, 12, 600991. [Google Scholar] [CrossRef] [PubMed]
- El Hajj Assaf, C.; Snini, S.P.; Tadrist, S.; Bailly, S.; Naylies, C.; Oswald, I.P.; Lorber, S.; Puel, O. Impact of veA on the development, aggressiveness, dissemination and secondary metabolism of Penicillium expansum. Mol. Plant Pathol. 2018, 19, 1971–1983. [Google Scholar] [CrossRef] [Green Version]
- Korpi, A.; Järnberg, J.; Pasanen, A. Microbial volatile organic compounds. Crit. Rev. Toxicol. 2009, 39, 139–193. [Google Scholar] [CrossRef]
- Hung, R.; Lee, S.; Bennett, J.W. Fungal volatile organic compounds and their role in ecosystems. Appl. Microbiol. Biotechnol. 2015, 99, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Almaliki, H.S.; Angela, A.; Goraya, N.J.; Yin, G.; Bennett, J.W. Volatile organic compounds produced by human pathogenic fungi are toxic to Drosophila melanogaster. Front. Fungal Biol. 2021, 1, 1–11. [Google Scholar] [CrossRef]
- Macedo, G.E.; de Brum Vieira, P.; Rodrigues, N.R.; Gomes, K.K.; Martins, I.K.; Franco, J.L.; Posser, T. Fungal compound 1-octen-3-ol induces mitochondrial morphological alterations and respiration dysfunctions in Drosophila melanogaster. Ecotoxicol. Environ. Saf. 2020, 206, 111232. [Google Scholar] [CrossRef]
- Inamdar, A.A.; Hossain, M.M.; Bernstein, A.I.; Miller, G.W.; Richardson, J.R.; Bennett, J.W. Fungal-Derived semiochemical 1-octen-3-ol disrupts dopamine packaging and causes neurodegeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 19561–19566. [Google Scholar] [CrossRef] [Green Version]
- Inamdar, A.A.; Masurekar, P.; Bennett, J.W. Neurotoxicity of fungal volatile organic compounds in Drosophila melanogaster. Toxicol. Sci. 2010, 117, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inamdar, A.A.; Bennett, J.W. A common fungal volatile organic compound induces a nitric oxide mediated inflammatory response in Drosophila melanogaster. Sci. Rep. 2014, 4, 3833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, G.; Zhang, Y.; Fu, M.; Hua, S.S.T.; Huang, Q.; Pennerman, K.K.; Wu, G.; Jurick, W.M.; Lee, S.; Bu, L. Influence of R and S enantiomers of 1-octen-3-ol on gene expression of Penicillium chrysogenum. J. Ind. Microbiol. Biotechnol. 2019, 46, 977–991. [Google Scholar] [CrossRef]
- Brodhun, F.; Feussner, I. Oxylipins in fungi. FEBS J. 2011, 278, 1047–1063. [Google Scholar] [CrossRef]
- Fierro, F.; Barredo, J.L.; Diez, B.; Gutierrez, S.; Fernandez, F.J.; Martin, J.F. The penicillin gene cluster is amplified in tandem repeats linked by conserved hexanucleotide sequences. Proc. Natl. Acad. Sci. USA 1995, 92, 6200–6204. [Google Scholar] [CrossRef] [Green Version]
- Wisecaver, J.H.; Rokas, A. Fungal metabolic gene clusters-caravans traveling across genomes and environments. Front. Microbiol. 2015, 6, 161. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Penicillium spp. Assembly | R21 | NJ1 | 113 |
|---|---|---|---|
| No. of all scaffolds | 257 | 91 | 228 |
| Bases in all scaffolds (bp) | 35,046,069 | 31,778,960 | 34,025,097 |
| No. of large scaffolds (>1000 bp) | 244 | 48 | 135 |
| Bases in large scaffolds (>1000 bp) | 35,037,781 | 31,755,870 | 33,977,500 |
| Largest length (bp) | 1,693,895 | 5,141,995 | 4,666,273 |
| Scaffold N50 (bp) | 377,946 | 3,327,763 | 1,432,551 |
| Scaffold N90 (bp) | 791,867 | 1,217,655 | 346,259 |
| GC content | 48.23% | 47.95% | 48.63% |
| N rate | 0.53% | 0.81% | 1.33% |
| No. of all contigs | 573 | 954 | 737 |
| Bases in all contigs (bp) | 34,858,998 | 31,522,745 | 33,572,562 |
| No. of large contigs (>1000 bp) | 556 | 712 | 511 |
| Bases in large contigs (bp) | 34,847,611 | 31,399,673 | 33,460,720 |
| Length of largest contig (bp) | 556,579 | 346,776 | 614,748 |
| Contig N50 (bp) | 108,376 | 93,897 | 157,787 |
| Contig N90 (bp) | 35,359 | 26,540 | 39,668 |
| Penicillium spp. gene prediction | |||
| No. genes | 12,707 | 12,057 | 12,649 |
| Gene total length (bp) | 18,790,875 | 17,215,819 | 17,946,096 |
| Gene density (genes per kb) | 0.362 | 0.379 | 0.371 |
| Gene average length | 1478 | 1427 | 1418 |
| GC content in gene region (%) | 52.3 | 52.2 | 52.4 |
| Gene/genome (%) | 53.6 | 54.2 | 52.7 |
| Intergenetic region length (bp) | 16,255,194 | 14,563,141 | 16,079,001 |
| GC content in intergenetic region (%) | 43.4 | 43.0 | 44.4 |
| Intergenetic length/genome (%) | 46.4 | 45.8 | 47.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, G.; Zhao, H.; Pennerman, K.K.; Jurick, W.M., II; Fu, M.; Bu, L.; Guo, A.; Bennett, J.W. Genomic Analyses of Penicillium Species Have Revealed Patulin and Citrinin Gene Clusters and Novel Loci Involved in Oxylipin Production. J. Fungi 2021, 7, 743. https://doi.org/10.3390/jof7090743
Yin G, Zhao H, Pennerman KK, Jurick WM II, Fu M, Bu L, Guo A, Bennett JW. Genomic Analyses of Penicillium Species Have Revealed Patulin and Citrinin Gene Clusters and Novel Loci Involved in Oxylipin Production. Journal of Fungi. 2021; 7(9):743. https://doi.org/10.3390/jof7090743
Chicago/Turabian StyleYin, Guohua, Hui Zhao, Kayla K. Pennerman, Wayne M. Jurick, II, Maojie Fu, Lijing Bu, Anping Guo, and Joan W. Bennett. 2021. "Genomic Analyses of Penicillium Species Have Revealed Patulin and Citrinin Gene Clusters and Novel Loci Involved in Oxylipin Production" Journal of Fungi 7, no. 9: 743. https://doi.org/10.3390/jof7090743
APA StyleYin, G., Zhao, H., Pennerman, K. K., Jurick, W. M., II, Fu, M., Bu, L., Guo, A., & Bennett, J. W. (2021). Genomic Analyses of Penicillium Species Have Revealed Patulin and Citrinin Gene Clusters and Novel Loci Involved in Oxylipin Production. Journal of Fungi, 7(9), 743. https://doi.org/10.3390/jof7090743