
The Siderophore Transporters Sit1 and Sit2 Are Essential for Utilization of Ferrichrome-, Ferrioxamine- and Coprogen-Type Siderophores in Aspergillus fumigatus

 , , , and
, , , and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Type | Example 1 | Exemplary Producer 2 |
|---|---|---|---|
| Hydroxymates | fusarinines | fusarinine C | A. fumigatus, A. nidulans |
| triacetylfusarinine C | A. fumigatus, A. nidulans | ||
| coprogens | coprogen | Penicillium chrysogenum | |
| coprogen B | Neurospora crassa | ||
| ferrichromes | ferrichrome | A. nidulans, Ustilago maydis | |
| ferrichrome A | Ustilago maydis | ||
| ferricrocin hydroxyferricrocin | A. fumigatus, A. nidulans A. fumigatus | ||
| ferrichrysin | A. terreus, A. flavus | ||
| ferrirubin | A. ochraceous | ||
| ferrirhodin | Ceratobasidium globisporum | ||
| VL-2397 | Acremonium persicinium | ||
| ferrioxamines | ferrioxamine B | Streptomyces pilosus | |
| ferrioxamine E | Streptomyces olivaceus | ||
| ferrioxamine G | Erwinia amylovora | ||
| rhodotorulic acid | rhodotorulic acid | Rhodotorula glutinis | |
| trishydroxamate | basidiochrome | Rhizoctonia mucoroides | |
| Carboxylates | hydroxycarboxylate | rhizoferrin | Rhizopus delemar, Francisella tularensis |
| Mixed types | hydroxamate/ hydroxycarboxylate | ornibactin schizokinen | Burkholderia cepacia Bacillus megaterium |
| pyoverdine 3 | Pseudomonas aeruginosa | ||
| Catecholates | enterobactin | Escherichia coli |
2. Materials and Methods
2.1. Growth Conditions
2.2. A. fumigatus Mutant Strains Generation
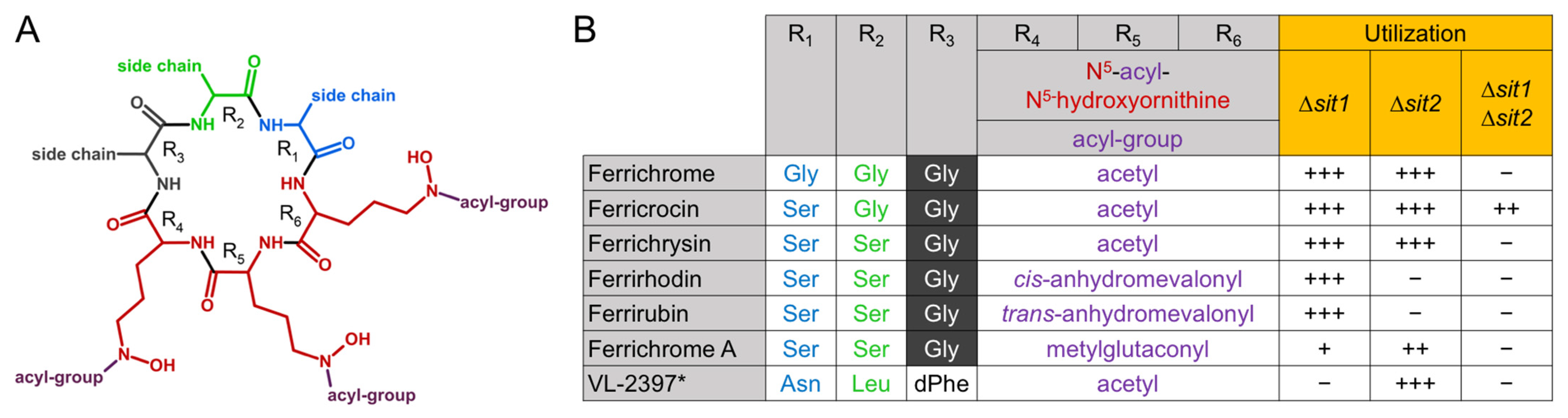
2.3. Siderophores
2.4. Fluorescence Microscopy
2.5. Bioinformatics
3. Results
3.1. Characterization of Siderophore Uptake in A. fumigatus Mutants Lacking Both RIA and Siderophore Biosynthesis in Combination with Deficiency in Sit1 and/or Sit2
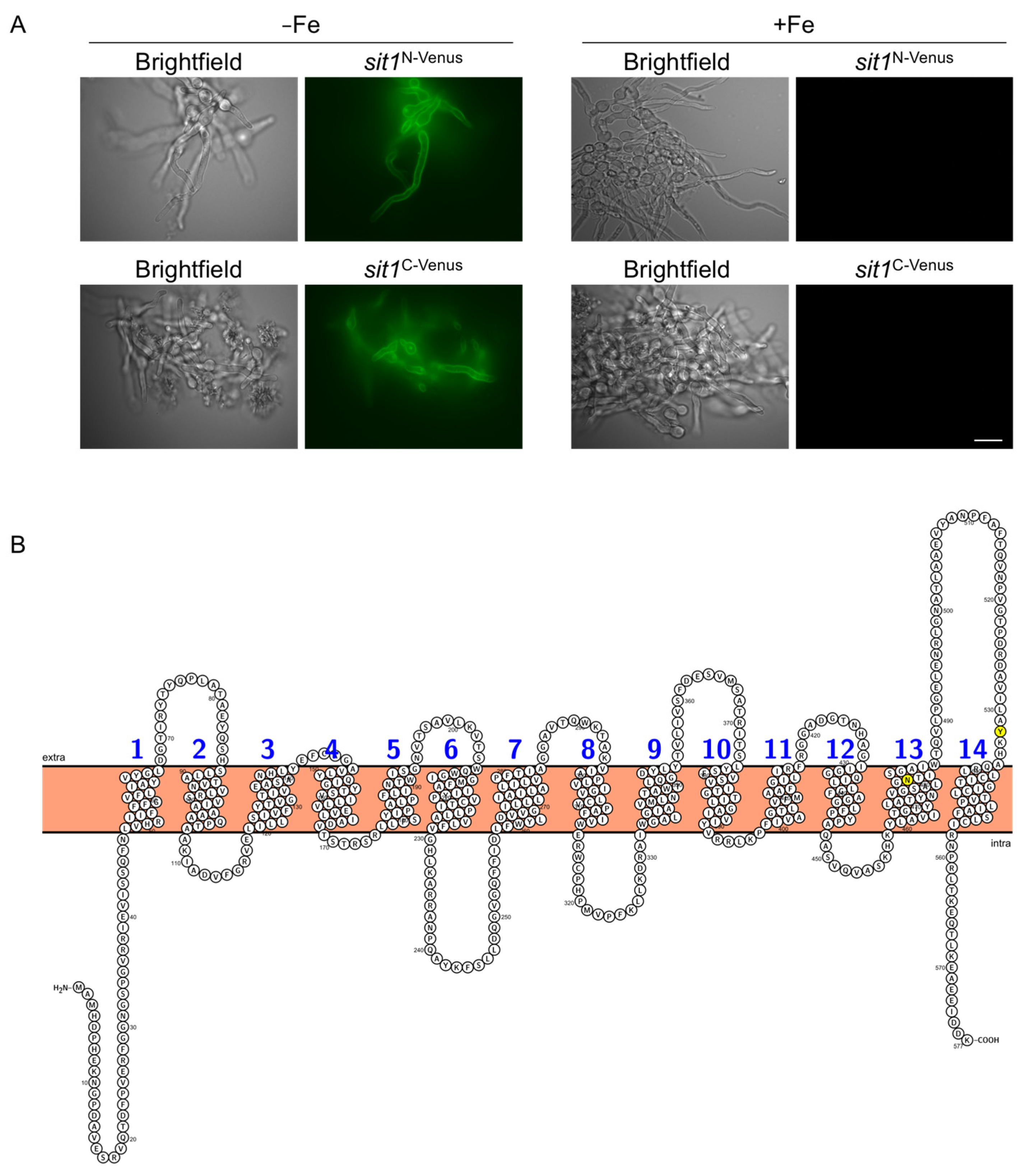
3.2. Sit1 Is Localized in the Plasma Membrane and Its Production Is Induced by Iron Starvation
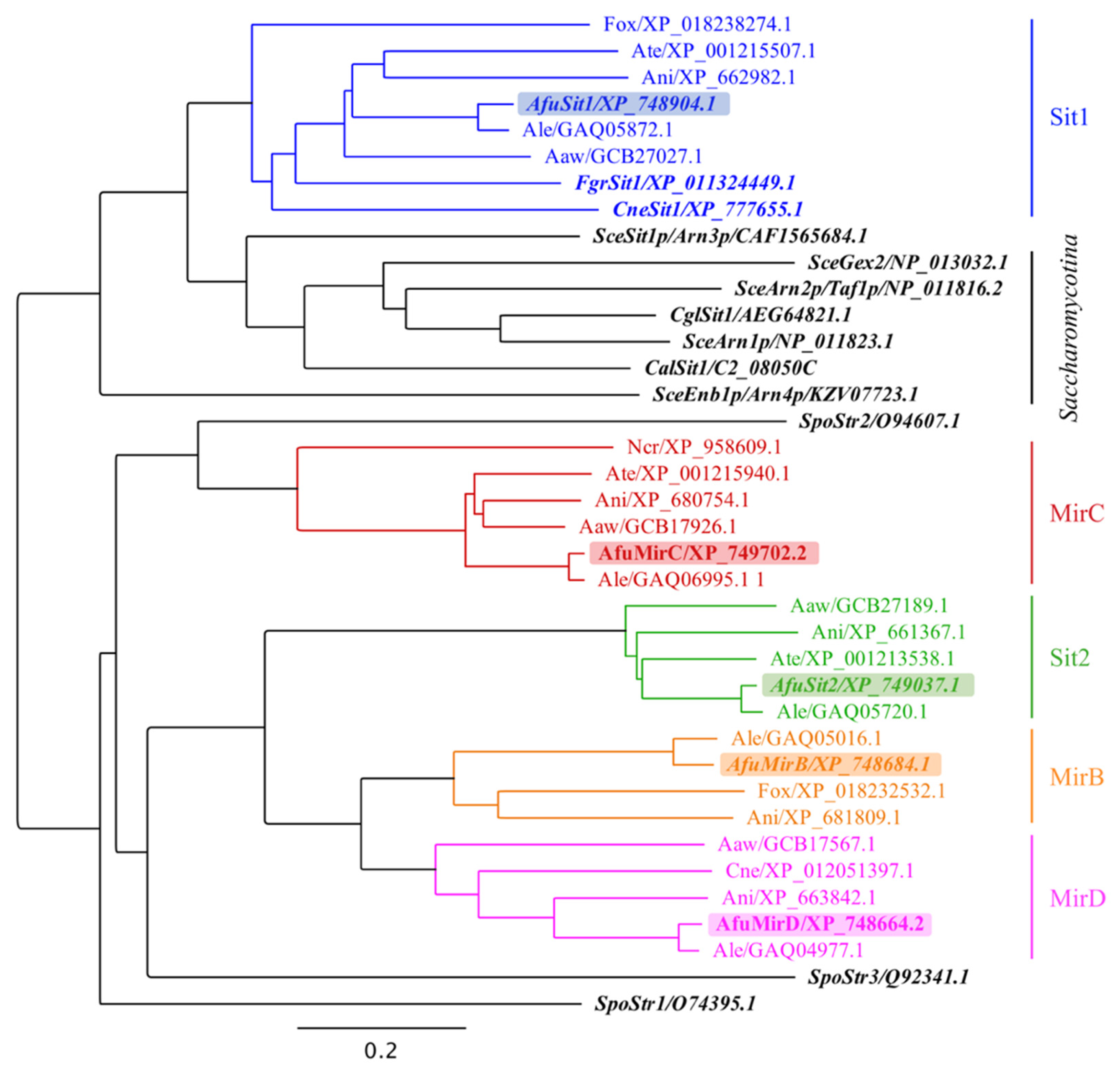
3.3. A. fumigatus Siderophore Transporters Belong to Different Subclades
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef]
- Latgé, J.-P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef]
- Misslinger, M.; Hortschansky, P.; Brakhage, A.A.; Haas, H. Fungal iron homeostasis with a focus on Aspergillus fumigatus. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1868, 118885. [Google Scholar] [CrossRef] [PubMed]
- Haas, H. Iron—A Key Nexus in the Virulence of Aspergillus fumigatus. Front. Microbiol. 2012, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, C.D.; Kaplan, J. Iron Acquisition and Transcriptional Regulation. Chem. Rev. 2009, 109, 4536–4552. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.; Ozkaya, O.; Kummerli, R. Bacterial siderophores in community and host interactions. Nat. Rev. Microbiol. 2019, 18, 152–163. [Google Scholar] [CrossRef]
- Haas, H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [Green Version]
- Gründlinger, M.; Yasmin, S.; Lechner, B.E.; Geley, S.; Schrettl, M.; Hynes, M.; Haas, H. Fungal siderophore biosynthesis is partially localized in peroxisomes. Mol. Microbiol. 2013, 88, 862–875. [Google Scholar] [CrossRef] [Green Version]
- Haas, H.; Eisendle, M.; Turgeon, B.G. Siderophores in fungal physiology and virulence. Annu. Rev. Phytopathol. 2008, 46, 149–187. [Google Scholar] [CrossRef]
- Kumar, S.; Kc, R.; Sanford, L.; Hernandez, A.; Kakarla, P.; Varela, M. Structural and functional roles of two evolutionarily conserved amino acid sequence motifs within solute transporters of the major facilitator superfamily. Trends Cell Mol. Biol. 2016, 11, 41–53. [Google Scholar]
- Bai, X.; Moraes, T.F.; Reithmeier, R.A.F. Structural biology of solute carrier (SLC) membrane transport proteins. Mol. Membr. Biol. 2017, 34, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Lekshmi, M.; Parvathi, A.; Ojha, M.; Wenzel, N.; Varela, M.F. Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps. Microorganisms 2020, 8, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philpott, C.C. Iron uptake in fungi: A system for every source. Biochim. Biophys. Acta 2006, 1763, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, H. Molecular genetics of fungal siderophore biosynthesis and uptake: The role of siderophores in iron uptake and storage. Appl. Microbiol. Biotechnol. 2003, 62, 316–330. [Google Scholar] [CrossRef]
- Raymond-Bouchard, I.; Carroll, C.S.; Nesbitt, J.R.; Henry, K.A.; Pinto, L.J.; Moinzadeh, M.; Scott, J.K.; Moore, M.M. Structural requirements for the activity of the MirB ferrisiderophore transporter of Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Kim, J.Y.; Yun, C.W. Identification of ferrichrome- and ferrioxamine B-mediated iron uptake by Aspergillus fumigatus. Biochem. J. 2016, 473, 1203–1213. [Google Scholar] [CrossRef] [Green Version]
- Challis, G.L. A widely distributed bacterial pathway for siderophore biosynthesis independent of nonribosomal peptide synthetases. Chembiochem 2005, 6, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Polsinelli, I.; Borruso, L.; Caliandro, R.; Triboli, L.; Esposito, A.; Benini, S. A genome-wide analysis of desferrioxamine mediated iron uptake in Erwinia spp. reveals genes exclusive of the Rosaceae infecting strains. Sci. Rep. 2019, 9, 2818. [Google Scholar] [CrossRef] [Green Version]
- Mulvihill, E.D.; Moloney, N.M.; Owens, R.A.; Dolan, S.K.; Russell, L.; Doyle, S. Functional Investigation of Iron-Responsive Microsomal Proteins, including MirC, in Aspergillus fumigatus. Front. Microbiol. 2017, 8, 418. [Google Scholar] [CrossRef] [Green Version]
- Petrik, M.; Haas, H.; Schrettl, M.; Helbok, A.; Blatzer, M.; Decristoforo, C. In vitro and in vivo evaluation of selected 68Ga-siderophores for infection imaging. Nucl. Med. Biol. 2012, 39, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Carroll, C.S.; Amankwa, L.N.; Pinto, L.J.; Fuller, J.D.; Moore, M.M. Detection of a Serum Siderophore by LC-MS/MS as a Potential Biomarker of Invasive Aspergillosis. PLoS ONE 2016, 11, e0151260. [Google Scholar] [CrossRef]
- Hoenigl, M.; Orasch, T.; Faserl, K.; Prattes, J.; Loeffler, J.; Springer, J.; Gsaller, F.; Reischies, F.; Duettmann, W.; Raggam, R.B.; et al. Triacetylfusarinine C: A urine biomarker for diagnosis of invasive aspergillosis. J. Infect. 2019, 78, 150–157. [Google Scholar] [CrossRef] [Green Version]
- Orasch, T.; Prattes, J.; Faserl, K.; Eigl, S.; Duttmann, W.; Lindner, H.; Haas, H.; Hoenigl, M. Bronchoalveolar lavage triacetylfusarinine C (TAFC) determination for diagnosis of invasive pulmonary aspergillosis in patients with hematological malignancies. J. Infect. 2017, 75, 370–373. [Google Scholar] [CrossRef] [Green Version]
- Petrik, M.; Pfister, J.; Misslinger, M.; Decristoforo, C.; Haas, H. Siderophore-Based Molecular Imaging of Fungal and Bacterial Infections-Current Status and Future Perspectives. J. Fungi 2020, 6, 73. [Google Scholar] [CrossRef]
- Petrik, M.; Zhai, C.; Haas, H.; Decristoforo, C. Siderophores for molecular imaging applications. Clin. Transl. Imaging 2017, 5, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misslinger, M.; Petrik, M.; Pfister, J.; Hubmann, I.; Bendova, K.; Decristoforo, C.; Haas, H. Desferrioxamine B-mediated pre-clinical in vivo imaging of infection by the mold fungus Aspergillus fumigatus. J. Fungi 2021, 7, 734. [Google Scholar] [CrossRef]
- Pfister, J.; Bata, R.; Hubmann, I.; Hörmann, A.A.; Gsaller, F.; Haas, H.; Decristoforo, C. Siderophore Scaffold as Carrier for Antifungal Peptides in Therapy of Aspergillus fumigatus Infections. J. Fungi 2020, 6, 367. [Google Scholar] [CrossRef]
- Dietl, A.M.; Misslinger, M.; Aguiar, M.M.; Ivashov, V.; Teis, D.; Pfister, J.; Decristoforo, C.; Hermann, M.; Sullivan, S.M.; Smith, L.R.; et al. The Siderophore Transporter Sit1 Determines Susceptibility to the Antifungal VL-2397. Antimicrob. Agents Chemother. 2019, 63, e00807-19. [Google Scholar] [CrossRef] [PubMed]
- Sass, G.; Nazik, H.; Penner, J.; Shah, H.; Ansari, S.R.; Clemons, K.V.; Groleau, M.-C.; Dietl, A.-M.; Visca, P.; Haas, H.; et al. Studies of Pseudomonas aeruginosa Mutants Indicate Pyoverdine as the Central Factor in Inhibition of Aspergillus fumigatus Biofilm. J. Bacteriol. 2018, 200, e00345-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontecorvo, G.; Roper, J.A.; Chemmons, L.M.; Macdonald, K.D.; Bufton, A.W.J. The Genetics of Aspergillus nidulans. Adv. Genet. 1953, 5, 141–238. [Google Scholar]
- Hartmann, T.; Dumig, M.; Jaber, B.M.; Szewczyk, E.; Olbermann, P.; Morschhauser, J.; Krappmann, S. Validation of a self-excising marker in the human pathogen Aspergillus fumigatus by employing the beta-rec/six site-specific recombination system. Appl. Environ. Microbiol. 2010, 76, 6313–6317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krappmann, S.; Sasse, C.; Braus, G.H. Gene targeting in Aspergillus fumigatus by homologous recombination is facilitated in a nonhomologous end- joining-deficient genetic background. Eukaryot. Cell 2006, 5, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Zadra, I.; Abt, B.; Parson, W.; Haas, H. xylP Promoter-Based Expression System and Its Use for Antisense Downregulation of the Penicillium chrysogenumNitrogen Regulator NRE. Appl. Environ. Microbiol. 2000, 66, 4810–4816. [Google Scholar] [CrossRef] [Green Version]
- Birštonas, L.; Dallemulle, A.; López-Berges, M.S.; Jacobsen, I.D.; Offterdinger, M.; Abt, B.; Straßburger, M.; Bauer, I.; Schmidt, O.; Sarg, B.; et al. Multiplex Genetic Engineering Exploiting Pyrimidine Salvage Pathway-Based Endogenous Counterselectable Markers. mBio 2020, 11, e00230-20. [Google Scholar] [CrossRef] [Green Version]
- Tilburn, J.; Scazzocchio, C.; Taylor, G.G.; Zabicky-Zissman, J.H.; Lockington, R.A.; Davies, R.W. Transformation by integration in Aspergillus nidulans. Gene 1983, 26, 205–221. [Google Scholar] [CrossRef]
- Oberegger, H.; Schoeser, M.; Zadra, I.; Abt, B.; Haas, H. SREA is involved in regulation of siderophore biosynthesis, utilization and uptake in Aspergillus nidulans. Mol. Microbiol. 2001, 41, 1077–1089. [Google Scholar] [CrossRef]
- Schrettl, M.; Bignell, E.; Kragl, C.; Sabiha, Y.; Loss, O.; Eisendle, M.; Wallner, A.; Arst, H.N., Jr.; Haynes, K.; Haas, H. Distinct roles for intra- and extracellular siderophores during Aspergillus fumigatus infection. PLoS Pathog. 2007, 3, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Engel, S.R.; Dietrich, F.S.; Fisk, D.G.; Binkley, G.; Balakrishnan, R.; Costanzo, M.C.; Dwight, S.S.; Hitz, B.C.; Karra, K.; Nash, R.S.; et al. The Reference Genome Sequence of Saccharomyces cerevisiae: Then and Now. G3 Genes Genomes Genet. 2014, 4, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Skrzypek, M.S.; Binkley, J.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida Genome Database (CGD): Incorporation of Assembly 22, systematic identifiers and visualization of high throughput sequencing data. Nucleic Acids Res. 2017, 45, D592–D596. [Google Scholar] [CrossRef] [Green Version]
- Sayers, E.W.; Agarwala, R.; Bolton, E.E.; Brister, J.R.; Canese, K.; Clark, K.; Connor, R.; Fiorini, N.; Funk, K.; Hefferon, T.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2019, 47, D23–D28. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [Green Version]
- Schrettl, M.; Bignell, E.; Kragl, C.; Joechl, C.; Rogers, T.; Arst, H.N., Jr.; Haynes, K.; Haas, H. Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence. J. Exp. Med. 2004, 200, 1213–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, I.; Ohsumi, K.; Takeda, S.; Katsumata, K.; Matsumoto, S.; Akamatsu, S.; Mitori, H.; Nakai, T. ASP2397 Is a Novel Natural Compound That Exhibits Rapid and Potent Fungicidal Activity against Aspergillus Species through a Specific Transporter. Antimicrob. Agents Chemother. 2019, 63, e02689-18. [Google Scholar] [CrossRef] [PubMed]
- Nevitt, T.; Thiele, D.J. Host iron withholding demands siderophore utilization for Candida glabrata to survive macrophage killing. PLoS Pathog. 2011, 7, e1001322. [Google Scholar] [CrossRef] [PubMed]
- Heymann, P.; Gerads, M.; Schaller, M.; Dromer, F.; Winkelmann, G.; Ernst, J.F. The siderophore iron transporter of Candida albicans (Sit1p/Arn1p) mediates uptake of ferrichrome-type siderophores and is required for epithelial invasion. Infect. Immun. 2002, 70, 5246–5255. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, B.; Beaudoin, J.; Philpott, C.C.; Labbé, S. Fep1 represses expression of the fission yeast Schizosaccharomyces pombe siderophore-iron transport system. Nucleic Acids Res. 2003, 31, 4332–4344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plante, S.; Labbe, S. Spore Germination Requires Ferrichrome Biosynthesis and the Siderophore Transporter Str1 in Schizosaccharomyces pombe. Genetics 2019, 211, 893–911. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Kim, T.H.; Chang, H.I.; Sung, H.C.; Yun, C.W. Cellular iron utilization is regulated by putative siderophore transporter FgSit1 not by free iron transporter in Fusarium graminearum. Biochem. Biophys. Res. Commun. 2006, 345, 1634–1642. [Google Scholar] [CrossRef]
- Tangen, K.L.; Jung, W.H.; Sham, A.P.; Lian, T.; Kronstad, J.W. The iron- and cAMP-regulated gene SIT1 influences ferrioxamine B utilization, melanization and cell wall structure in Cryptococcus neoformans. Microbiology 2007, 153, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, J.; Petrik, M.; Bendova, K.; Matuszczak, B.; Binder, U.; Misslinger, M.; Kühbacher, A.; Gsaller, F.; Haas, H.; Decristoforo, C. Antifungal Siderophore Conjugates for Theranostic Applications in Invasive Pulmonary Aspergillosis Using Low-Molecular TAFC Scaffolds. J. Fungi 2021, 7, 558. [Google Scholar] [CrossRef] [PubMed]
- Celia, H.; Noinaj, N.; Buchanan, S.K. Structure and Stoichiometry of the Ton Molecular Motor. Int. J. Mol. Sci. 2020, 21, 375. [Google Scholar] [CrossRef] [Green Version]
- Endicott, N.P.; Lee, E.; Wencewicz, T.A. Structural Basis for Xenosiderophore Utilization by the Human Pathogen Staphylococcus aureus. ACS Infect. Dis. 2017, 3, 542–553. [Google Scholar] [CrossRef]
- Haas, H.; Schoeser, M.; Lesuisse, E.; Ernst, J.F.; Parson, W.; Abt, B.; Winkelmann, G.; Oberegger, H. Characterization of the Aspergillus nidulans transporters for the siderophores enterobactin and triacetylfusarinine C. Biochem. J. 2003, 371, 505–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sass, G.; Ansari, S.R.; Dietl, A.M.; Déziel, E.; Haas, H.; Stevens, D.A. Intermicrobial interaction: Aspergillus fumigatus siderophores protect against competition by Pseudomonas aeruginosa. PLoS ONE 2019, 14, e0216085. [Google Scholar] [CrossRef] [Green Version]
- Sass, G.; Nazik, H.; Penner, J.; Shah, H.; Ansari, S.R.; Clemons, K.V.; Groleau, M.C.; Dietl, A.M.; Visca, P.; Haas, H.; et al. Aspergillus-Pseudomonas interaction, relevant to competition in airways. Med. Mycol. 2019, 57, S228–S232. [Google Scholar] [CrossRef] [PubMed]
- Dhaoui, M.; Auchere, F.; Blaiseau, P.L.; Lesuisse, E.; Landoulsi, A.; Camadro, J.M.; Haguenauer-Tsapis, R.; Belgareh-Touze, N. Gex1 is a yeast glutathione exchanger that interferes with pH and redox homeostasis. Mol. Biol. Cell 2011, 22, 2054–2067. [Google Scholar] [CrossRef] [PubMed]
- Normant, V.; Mourer, T.; Labbé, S. The major facilitator transporter Str3 is required for low-affinity heme acquisition in Schizosaccharomyces pombe. J. Biol. Chem. 2018, 293, 6349–6362. [Google Scholar] [CrossRef] [Green Version]
- McDonagh, A.; Fedorova, N.D.; Crabtree, J.; Yu, Y.; Kim, S.; Chen, D.; Loss, O.; Cairns, T.; Goldman, G.; Armstrong-James, D.; et al. Sub-telomere directed gene expression during initiation of invasive aspergillosis. PLoS Pathog. 2008, 4, e1000154. [Google Scholar] [CrossRef] [PubMed]
- Essén, S.A.; Bylund, D.; Holmström, S.J.M.; Moberg, M.; Lundström, U.S. Quantification of Hydroxamate Siderophores in Soil Solutions of Podzolic Soil Profiles in Sweden. Biometals 2006, 19, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, G. Ecology of siderophores with special reference to the fungi. Biometals 2007, 20, 379–392. [Google Scholar] [CrossRef] [PubMed]




| Strain | MIC [mg/L] |
|---|---|
| AfS77 | 1 |
| sit1N-Venus | 0.125 |
| sit1C-Venus | 0.5 |
| ∆sit1 | >16 |
| Species (Number of SITs) | SIT Name 1 | Substrate 2 | Non-Substrate 2 | Reference |
|---|---|---|---|---|
| S. cerevisiae (4) | Arn1 * | ferrichrome, coprogen | ferrioxamine B, VL2397, TAFC, enterobactin | [14,30] |
| Sit1/Arn3 * | ferrichrome, coprogen, ferrioxamine B | VL-2397 TAFC enterobactin | [14,30] | |
| C. albicans (1) | Sit1/Arn3 * | ferrichrome, ferricrocin, ferrichrysin, ferrirubin, coprogen, TAFC | ferrioxamine B, ferrioxamine E, VL-2397, enterobactin, rhodotorulic acid | [30,48] |
| C. glabrata (1) | Sit1 * | ferrichrome, ferrirubin, coprogen, VL-2397 3 | ferrioxamine B, TAFC, enterobactin | [30,47] |
| S. pombe (2) | Str1 | ferrichrome | ferrioxamine B | [49,50] |
| Str2 | ferrichrome, ferrioxamine B | [49] | ||
| Fusarium graminearum (9) | Sit1 | ferrichrome, ferrioxamine B | TAFC | [51] |
| Cryptococcus neoformans (7) | Sit1 | ferrioxamine B | [52] | |
| A. fumigatus (5) | Sit1 | Ferrichrome ferrioxamine B | [18] | |
| Sit2 | ferrichrome | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguiar, M.; Orasch, T.; Misslinger, M.; Dietl, A.-M.; Gsaller, F.; Haas, H. The Siderophore Transporters Sit1 and Sit2 Are Essential for Utilization of Ferrichrome-, Ferrioxamine- and Coprogen-Type Siderophores in Aspergillus fumigatus. J. Fungi 2021, 7, 768. https://doi.org/10.3390/jof7090768
Aguiar M, Orasch T, Misslinger M, Dietl A-M, Gsaller F, Haas H. The Siderophore Transporters Sit1 and Sit2 Are Essential for Utilization of Ferrichrome-, Ferrioxamine- and Coprogen-Type Siderophores in Aspergillus fumigatus. Journal of Fungi. 2021; 7(9):768. https://doi.org/10.3390/jof7090768
Chicago/Turabian StyleAguiar, Mario, Thomas Orasch, Matthias Misslinger, Anna-Maria Dietl, Fabio Gsaller, and Hubertus Haas. 2021. "The Siderophore Transporters Sit1 and Sit2 Are Essential for Utilization of Ferrichrome-, Ferrioxamine- and Coprogen-Type Siderophores in Aspergillus fumigatus" Journal of Fungi 7, no. 9: 768. https://doi.org/10.3390/jof7090768
APA StyleAguiar, M., Orasch, T., Misslinger, M., Dietl, A. -M., Gsaller, F., & Haas, H. (2021). The Siderophore Transporters Sit1 and Sit2 Are Essential for Utilization of Ferrichrome-, Ferrioxamine- and Coprogen-Type Siderophores in Aspergillus fumigatus. Journal of Fungi, 7(9), 768. https://doi.org/10.3390/jof7090768