
Recent Developments in Metabolomics Studies of Endophytic Fungi

 , ,
, ,  , and
, and
Abstract
:1. Introduction
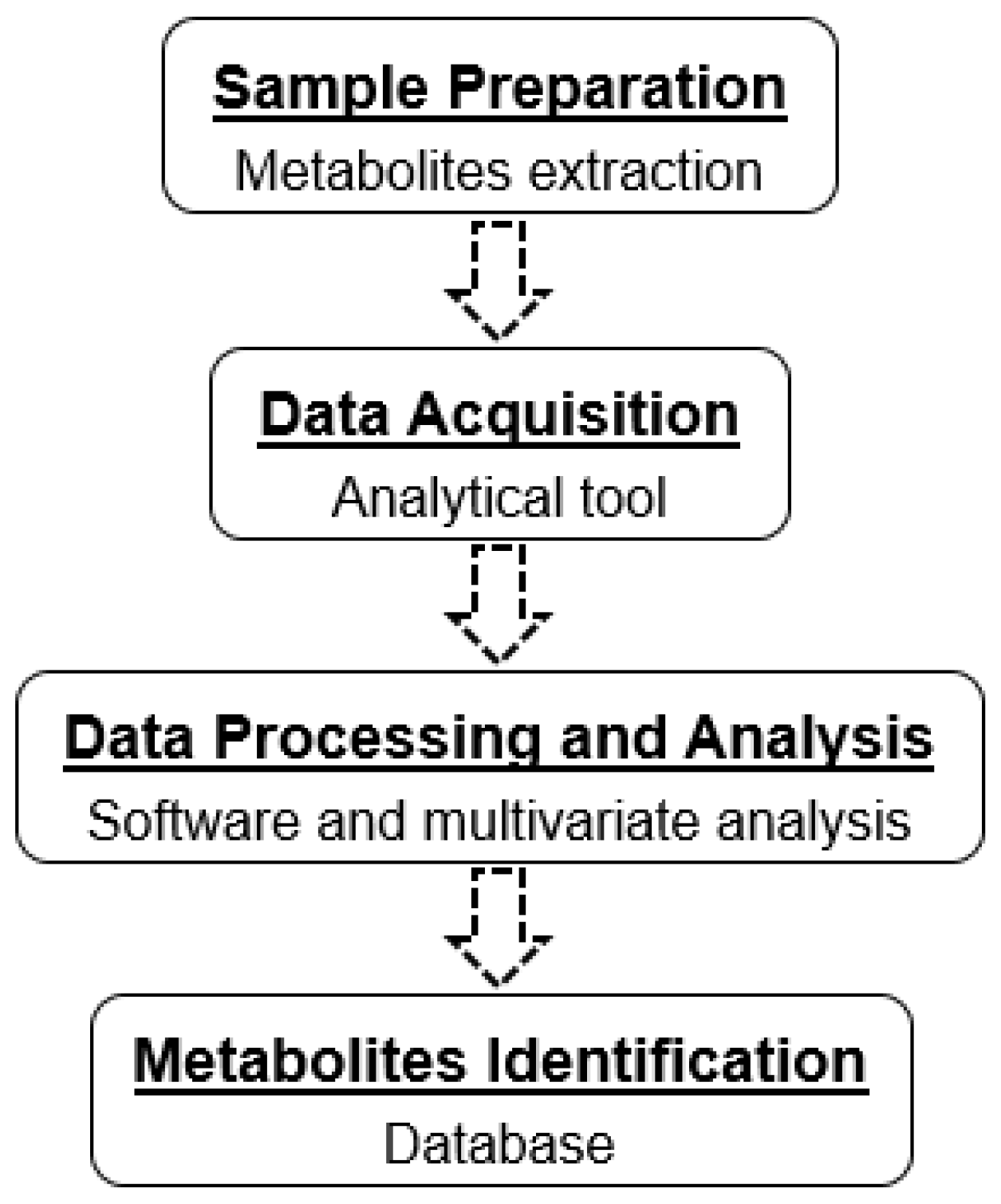
2. Multivariate Analysis
3. Metabolites Extraction
4. Advanced Analytical Tools for Metabolomics Studies
4.1. LC-HRMS-Based Metabolomics
4.2. NMR-Based Metabolomics
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ChEBI | Chemical Entities of Biological Interest |
| ESI | Electrospray ionization |
| LC | Liquid chromatography |
| LC-HRMS | Liquid chromatography–high-resolution mass spectrometry |
| LC-Q-ToF-MS | Liquid chromatography–quadrupole time-of-flight-mass spectrometry |
| LLE | Liquid–liquid extraction |
| MEB | Malt extract broth |
| MPDB | Acidified potato dextrose broth |
| MS | Mass spectrometry |
| m/z | Mass-to-charge |
| MIC | Minimum inhibitory concentration |
| NIST | National Institute of Science and Technology |
| NANPDB | Northern African Natural Products Database |
| NMR | Nuclear magnetic resonance |
| OPLS-DA | Orthogonal partial least square discriminant analysis |
| PLS-DA | Partial least square discriminant analysis |
| PCA | Principal component analysis |
| PDB | Potato dextrose broth |
| REB | Rice extract broth |
| SAB | Sabouraud broth |
| SSF | Solid-state fermentation |
| SMF | Submerged fermentation |
| T3DB | The Toxin and Toxin Target Database |
| UHPLC-HRMS | Ultra high-performance liquid chromatography–high-resolution mass spectrometry |
| UAE | Ultrasonic-assisted extraction |
| 1D 1H-NMR | One-dimensional proton-NMR |
References
- Rashmi, M.; Kushveer, J.S.; Sarma, V.V. A worldwide list of endophytic fungi with notes on ecology and diversity. Mycosphere 2019, 10, 798–1079. [Google Scholar] [CrossRef]
- Sayed, A.M.; Sherif, N.H.; El-Gendy, A.O.; Shamikh, Y.I.; Ali, A.T.; Attia, E.Z.; El-Katatny, M.H.; Khalifa, B.A.; Hassan, H.M.; Abdelmohsen, U.R. Metabolomic profiling and antioxidant potential of three fungal endophytes derived from Artemisia annua and Medicago sativa. Nat. Prod. Res. 2020. In Press. [Google Scholar] [CrossRef]
- Hartley, S.E.; Eschen, R.; Horwood, J.M.; Gange, A.C.; Hill, E.M. Infection by a foliar endophyte elicits novel arabidopside-based plant defence reactions in its host, Cirsium arvense. New Phytol. 2015, 205, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, K.; Tong, W.-Y.; Leong, C.-R.; Tan, W.-N. Potential of endophytic Diaporthe sp. as a new source of bioactive compounds. J. Microbiol. Biotechnol. 2021, 31, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, G. Analysis of secondary metabolites from plant endophytic fungi. Methods Mol. Biol. 2018, 1848, 25–38. [Google Scholar] [CrossRef]
- Vinale, F.; Nicoletti, R.; Lacatena, F.; Marra, R.; Sacco, A.; Lombardi, N.; d’Errico, G.; Digilio, M.C.; Lorito, M.; Woo, S.L. Secondary metabolites from the endophytic fungus Talaromyces pinophilus. Nat. Prod. Res. 2017, 31, 1778–1785. [Google Scholar] [CrossRef] [Green Version]
- Uzma, F.; Mohan, C.D.; Hashem, A.; Konappa, N.M.; Rangappa, S.; Kamath, P.V.; Singh, B.P.; Mudili, V.; Gupta, V.K.; Siddaiah, C.N.; et al. Endophytic fungi—Alternative sources of cytotoxic compounds: A review. Front. Pharmacol. 2018, 9, 309. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; Sharma, S. Endophytic fungi: A new hope for drug discovery. In New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, J., Gehlot, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 39–49. [Google Scholar]
- Alhadrami, H.A.; Sayed, A.M.; El-Gendy, A.O.; Shamikh, Y.I.; Gaber, Y.; Bakeer, W.; Sheirf, N.H.; Attia, E.Z.; Shaban, G.M.; Khalifa, B.A.; et al. A metabolomic approach to target antimalarial metabolites in the Artemisia annua fungal endophytes. Sci. Rep. 2021, 11, 2770. [Google Scholar] [CrossRef]
- Naik, B.S. Developments in taxol production through endophytic fungal biotechnology: A review. Orient. Pharm. Exp. Med. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Dayalan, S.; Xia, J.; Spicer, R.A.; Salek, R.; Roessner, U. Metabolome Analysis. In Encyclopedia of Bioinformatics and Computational Biology; Ranganathan, S., Gribskov, M., Nakai, K., Schönbach, C., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 396–409. [Google Scholar]
- Manchester, M.; Anand, A. Metabolomics: Strategies to define the role of metabolism in virus infection and pathogenesis. In Advances in Virus Research; Kielian, M., Mettenleiter, T.C., Roossinck, M.J., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 98, pp. 57–81. [Google Scholar]
- Ibrahim, A.; Tanney, J.B.; Fei, F.; Seifert, K.A.; Cutler, G.C.; Capretta, A.; Miller, J.D.; Sumarah, M.W. Metabolomic-guided discovery of cyclic nonribosomal peptides from Xylaria ellisii sp. nov., a leaf and stem endophyte of Vaccinium angustifolium. Sci. Rep. 2020, 10, 4599. [Google Scholar] [CrossRef] [Green Version]
- Tawfik, N.F.; Tawfike, A.F.; Abdou, R.; Abbott, G.; Abdelmohsen, U.R.; Edrada-Ebelm, R.; Haggag, E.G. Metabolomics and bioactivity guided isolation of secondary metabolites from the endophytic fungus Chaetomium sp. J. Adv. Pharm. Res. 2017, 1, 66–74. [Google Scholar] [CrossRef]
- Alonso, A.; Marsal, S.; Julia, A. Analytical methods in untargeted metabolomics: State of the art in 2015. Front. Bioeng. Biotechnol. 2015, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Fadiji, A.E.; Babalola, O.O. Elucidating mechanisms of endophytes used in plant protection and other bioactivities with multifunctional prospects. Front. Bioeng. Biotechnol. 2020, 8, 467. [Google Scholar] [CrossRef]
- Manganyi, M.C.; Ateba, C.N. Untapped potentials of endophytic fungi: A review of novel bioactive compounds with biological applications. Microorganisms 2020, 8, 1934. [Google Scholar] [CrossRef]
- Kamal, N.; Viegelmann, C.V.; Clements, C.J.; Edrada-Ebel, R. Metabolomics-guided isolation of anti-trypanosomal metabolites from the endophytic fungus Lasiodiplodia theobromae. Planta Med. 2017, 83, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Tawfike, A.F.; Abbott, G.; Young, L.; Edrada-Ebel, R. Metabolomic-guided isolation of bioactive natural products from Curvularia sp., an endophytic fungus of Terminalia laxiflora. Planta Med. 2018, 84, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, E.Z.; Farouk, H.M.; Abdelmohsen, U.R.; El-Katatny, M.H. Antimicrobial and extracellular oxidative enzyme activities of endophytic fungi isolated from alfalfa (Medicago sativa) assisted by metabolic profiling. S. Afr. J. Bot. 2020, 134, 156–162. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Mohammed, R.; Bahr, H.S.; Attia, E.Z.; El-Katatny, M.H.; Abelyan, N.; Al-Sanea, M.M.; Moawad, A.S.; Abdelmohsen, U.R. Soybean-associated endophytic fungi as potential source for anti-COVID-19 metabolites supported by docking analysis. J. Appl. Microbiol. 2021, 131, 1193–1211. [Google Scholar] [CrossRef]
- George, T.K.; Devadasan, D.; Jisha, M.S. Chemotaxonomic profiling of Penicillium setosum using high-resolution mass spectrometry (LC-Q-ToF-MS). Heliyon 2019, 5, e02484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triastuti, A.; Haddad, M.; Barakat, F.; Mejia, K.; Rabouille, G.; Fabre, N.; Amasifuen, C.; Jargeat, P.; Vansteelandt, M. Dynamics of chemical diversity during co-cultures: An integrative time-scale metabolomics study of fungal endophytes Cophinforma mamane and Fusarium solani. Chem. Biodivers. 2021, 18, e2000672. [Google Scholar] [CrossRef] [PubMed]
- Flores-Vallejo, R.C.; Folch-Mallol, J.L.; Sharma, A.; Cardoso-Taketa, A.; Alvarez-Berber, L.; Villarreal, M.L. ITS2 ribotyping, in vitro anti-inflammatory screening, and metabolic profiling of fungal endophytes from the Mexican species Crescentia alata Kunth. S. Afr. J. Bot. 2020, 134, 213–224. [Google Scholar] [CrossRef]
- Percival, B.; Gibson, M.; Leenders, J.; Wilson, P.B.; Grootveld, M. Univariate and multivariate statistical approaches to the analysis and interpretation of NMR-based metabolomics datasets of increasing complexity. In Computational Techniques for Analytical Chemistry and Bioanalysis; Wilson, P.B., Grootveld, M., Eds.; Royal Society of Chemistry: London, UK, 2020. [Google Scholar]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metab. 2013, 1, 92–107. [Google Scholar] [CrossRef]
- Wu, J.F.; Wang, Y. Multivariate analysis of metabolomics data. In Plant Metabolomics; Qi, X., Chen, X., Wang, Y., Eds.; Springer: Dordrecht, Germany, 2015. [Google Scholar]
- Ramana, P.; Adams, E.; Augustijns, P.; Van Schepdael, A. Metabonomics and drug development. In Metabonomics. Methods in Molecular Biology; Bjerrum, J., Ed.; Humana Press: New York, NY, USA, 2015; Volume 1277. [Google Scholar]
- Synytsya, A.; Monkai, J.; Bleha, R.; Macurkova, A.; Ruml, T.; Ahn, J.; Chukeatirote, E. Antimicrobial activity of crude extracts prepared from fungal mycelia. Asian Pac. J. Trop. Biomed. 2017, 7, 257–261. [Google Scholar] [CrossRef]
- Ezeobiora, C.E.; Igbokwe, N.H.; Amin, D.H.; Mendie, U.E. Endophytic microbes from Nigerian ethnomedicinal plants: A potential source for bioactive secondary metabolites—A review. Bull. Natl. Res. Cent. 2021, 45, 103. [Google Scholar] [CrossRef]
- de Carvalho, C.C.C.R. Fungi in Fermentation and Biotransformation Systems. In Biology of Microfungi. Fungal Biology; Li, D.W., Ed.; Springer: Cham, Switzerland, 2016; pp. 525–541. [Google Scholar]
- Srivastava, N.; Srivastava, M.; Ramteke, P.W.; Mishra, P.K. Solid-state fermentation strategy for microbial metabolites production: An overview. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.J., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 345–354. [Google Scholar]
- Costa, J.A.V.; Treichel, H.; Kumar, V.; Pandey, A. Advances in solid-state fermentation. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Larroche, C., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–17. [Google Scholar]
- Martínez-Medina, G.A.; Barragán, A.P.; Ruiz, H.A.; Ilyina, A.; Hernández, J.L.M.; Rodríguez-Jasso, R.M.; Hoyos-Concha, J.L.; Aguilar-González, C.N. Fungal proteases and production of bioactive peptides for the food industry. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 221–246. [Google Scholar]
- Kapoor, M.; Panwar, D.; Kaira, G.S. Bioprocesses for enzyme production using agro-industrial wastes: Technical challenges and commercialization potential. In Agro-Industrial Wastes as Feedstock for Enzyme Production; Dhillon, G.S., Kaur, S., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 61–93. [Google Scholar]
- Song, R.; Wang, J.; Sun, L.; Zhang, Y.; Ren, Z.; Zhao, B.; Lu, H. The study of metabolites from fermentation culture of Alternaria oxytropis. BMC Microbiol. 2019, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Fierascu, I.; Ortan, A.; Georgiev, M.I.; Sieniawska, E. Innovative approaches for recovery of phytoconstituents from medicinal/aromatic plants and biotechnological production. Molecules 2020, 25, 309. [Google Scholar] [CrossRef] [Green Version]
- Urkude, R.; Dhurvey, V.; Kochhar, S. Pesticide residues in beverages. In Quality Control in the Beverage Industry; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 529–560. [Google Scholar]
- Kyle, P.B. Toxicology: GCMS. In Mass Spectrometry for the Clinical Laboratory; Nair, H., Clarke, W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 131–163. [Google Scholar]
- Zahari, N.A.A.R.; Chong, G.H.; Abdullah, L.C.; Chua, B.L. Ultrasonic-assisted extraction (UAE) process on thymol concentration from Plectranthus Amboinicus leaves: Kinetic modeling and optimization. Processes 2020, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- David, A.; Rostkowski, P. Environmental Metabolomics; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 35–64. [Google Scholar]
- Zhang, T.; Chen, C.; Xie, K.; Wang, J.; Pan, Z. Current state of metabolomics research in meat quality analysis and authentication. Foods 2021, 10, 2388. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Huhman, D.V.; Sumner, L.W. Mass spectrometry strategies in metabolomics. J. Biol. Chem. 2011, 286, 25435–25442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR spectroscopy for metabolomics research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. NMR metabolomics: A look ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Sands, C.J.; Gómez-Romero, M.; Correia, G.; Chekmeneva, E.; Camuzeaux, S.; Izzi-Engbeaya, C.; Dhillo, W.S.; Takats, Z.; Lewis, M.R. Representing the metabolome with high fidelity: Range and response as quality control factors in LC-MS-based global profiling. Anal. Chem. 2021, 93, 1924–1933. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen rearrangement rules: Computational MS/MS fragmentation and structure elucidation using MS-FINDER software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2017, 15, 53–56. [Google Scholar] [CrossRef]
- Sorokina, M.; Steinbeck, C. Review on natural products databases: Where to find data in 2020. J. Cheminf. 2020, 12, 1–51. [Google Scholar] [CrossRef] [Green Version]
- Vinaixa, M.; Schymanski, E.L.; Neumann, S.; Navarro, M.; Salek, R.M.; Yanes, O. Mass spectral databases for LC/MS- and GC/MS-based metabolomics: State of the field and future prospects. Trends Anal. Chem. 2016, 78, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Zimowska, B.; Bielecka, M.; Abramczyk, B.; Nicoletti, R. Bioactive products from endophytic fungi of Sages (Salvia spp.). Agriculture 2020, 10, 543. [Google Scholar] [CrossRef]
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.X.; Ng, S.B. Fungal endophytes: A promising frontier for discovery of novel bioactive compounds. J. Fungi 2021, 7, 786. [Google Scholar] [CrossRef]
- Vinale, F.; Nicoletti, R.; Borrelli, F.; Mangoni, A.; Parisi, O.A.; Marra, R.; Lombardi, N.; Lacatena, F.; Grauso, L.; Finizio, S.; et al. Co-Culture of Plant Beneficial Microbes as Source of Bioactive Metabolites. Sci. Rep. 2017, 7, 14330. [Google Scholar] [CrossRef] [Green Version]
- Rai, N.; Keshri, P.K.; Verma, A.; Kamble, S.C.; Mishra, P.; Barik, S.; Singh, S.K.; Gautam, V. Plant associated fungal endophytes as a source of natural bioactive compounds. Mycology 2021, 12, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Ramdani, D.; Chaudhry, A.S.; Seal, C.J. Chemical composition, plant secondary metabolites, and minerals of green and black teas and the effect of different tea-to-water ratios during their extraction on the composition of their spent leaves as potential additives for ruminants. J. Agric. Food Chem. 2013, 61, 4961. [Google Scholar] [CrossRef]
- Ser, Z.; Liu, X.; Tang, N.N.; Locasale, J.W. Extraction parameters for metabolomics from cultured cells. Anal Biochem. 2015, 475, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Sitnikov, D.; Monnin, C.; Vuckovic, D. Systematic assessment of seven solvent and solid-phase extraction methods for metabolomics analysis of human plasma by LC-MS. Sci. Rep. 2016, 6, 38885. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, I.; Sternberg, S.; Williams, M.; Garrett, T.J. Comparison of global metabolite extraction strategies for soybeans using UHPLC-HRMS. Anal. Bioanal. Chem. 2017, 409, 6173–6180. [Google Scholar] [CrossRef] [PubMed]
- Tokuoka, M.; Sawamura, N.; Kobayashi, K.; Mizuno, A. Simple metabolite extraction method for metabolic profiling of the solid-state fermentation of Aspergillus oryzae. J. Biosci. Bioeng. 2010, 110, 665–669. [Google Scholar] [CrossRef]
- Vinaixa, M.; Samino, S.; Saez, I.; Duran, J.; Guinovart, J.J.; Yanes, O. A guideline to univariate statistical analysis for LC/MS-based untargeted metabolomics-derived data. Metabolites 2012, 2, 775–795. [Google Scholar] [CrossRef]
- Berg, M.; Vanaerschot, M.; Jankevics, A.; Cuypers, B.; Breitling, R.; Dujardin, J.C. LC-MS metabolomics from study design to data-analysis–Using a versatile pathogen as a test case. Comput. Struct. Biotechnol. J. 2013, 4, e201301002. [Google Scholar] [CrossRef]
- De Souza, L.P.; Alseekh, S.; Brotman, Y.; Fernie, A.R. Network-based strategies in metabolomics data analysis and interpretation: From molecular networking to biological interpretation. Expert Rev. Proteom. 2020, 17, 243–255. [Google Scholar] [CrossRef]
- Beniddir, M.A.; Bin Kang, K.; Genta-Jouve, G.; Huber, F.; Rogers, S.; van der Hooft, J.J.J. Advances in decomposing complex metabolite mixtures using substructure- and network-based computational metabolomics approaches. Nat. Prod. Rep. 2021, 38, 1967–1993. [Google Scholar] [CrossRef] [PubMed]
- Nalbantoglu, S. Metabolomics: Basic principles and strategies. In Molecular Medicine; Nalbantoglu, S., Amri, H., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Kellogg, J.J.; Graf, T.N.; Paine, M.F.; McCune, J.S.; Kvalheim, O.M.; Oberlies, N.H.; Cech, N.B. Comparison of metabolomics approaches for evaluating the variability of complex botanical preparations: Green tea (Camellia sinensis) as a case study. J. Nat. Prod. 2017, 80, 1457–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowley, T.E. Nuclear magnetic resonance spectroscopy. In Purification and Characterization of Secondary Metabolites; Crowley, T.E., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 67–78. [Google Scholar]
- Tampieri, A.; Szabó, M.; Medina, F.; Gulyás, H. A brief introduction to the basics of NMR spectroscopy and selected examples of its applications to materials characterization. Phys. Sci. Rev. 2020, 6, 1–41. [Google Scholar] [CrossRef]
- Decker, S.R.; Harman-Ware, A.E.; Happs, R.M.; Wolfrum, E.J.; Tuskan, G.A.; Kainer, D.; Oguntimein, G.B.; Rodriguez, M.; Weighill, D.; Jones, P.; et al. High throughput screening technologies in biomass characterization. Front. Energy Res. 2018, 6, 120. [Google Scholar] [CrossRef] [Green Version]
- Vögele, J.; Ferner, J.-P.; Altincekic, N.; Bains, J.K.; Ceylan, B.; Fürtig, B.; Grün, J.T.; Hengesbach, M.; Hohmann, K.F.; Hymon, D.; et al. 1H, 13C, 15N and 31P chemical shift assignment for stem-loop 4 from the 5’-UTR of SARS-CoV-2. Biomol. NMR Assign. 2021, 15, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Bao, Z.-X.; Zhao, P.-J.; Li, G.-H. Advances in the study of metabolomics and metabolites in some species interactions. Molecules 2021, 26, 3311. [Google Scholar] [CrossRef]
- Gowda, G.A.N.; Raftery, D. Can NMR solve some significant challenges in metabolomics? J. Magn. Reson. 2015, 260, 144–160. [Google Scholar] [CrossRef] [Green Version]
- Bingol, K. Recent advances in targeted and untargeted metabolomics by NMR and MS/NMR Methods. High-Throughput 2018, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Dona, A.C.; Kyriakides, M.; Scott, F.; Shephard, E.A.; Varshavi, D.; Veselkov, K.; Everett, J.R. A guide to the identification of metabolites in NMR-based metabonomics/metabolomics experiments. Comput. Struct. Biotechnol. J. 2016, 14, 135–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Perez, I.; Posma, J.M.; Serrano-Contreras, J.I.; Boulangé, C.L.; Chan, Q.; Frost, G.; Stamler, J.; Elliott, P.; Lindon, J.; Holmes, E.; et al. Identifying unknown metabolites using NMR-based metabolic profiling techniques. Nat. Protoc. 2020, 15, 2538–2567. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Endophytic Fungi | Host Plant | Metabolite Extraction | Solvent Used | Analytical Tool | Database | Metabolites | Ref. |
|---|---|---|---|---|---|---|---|
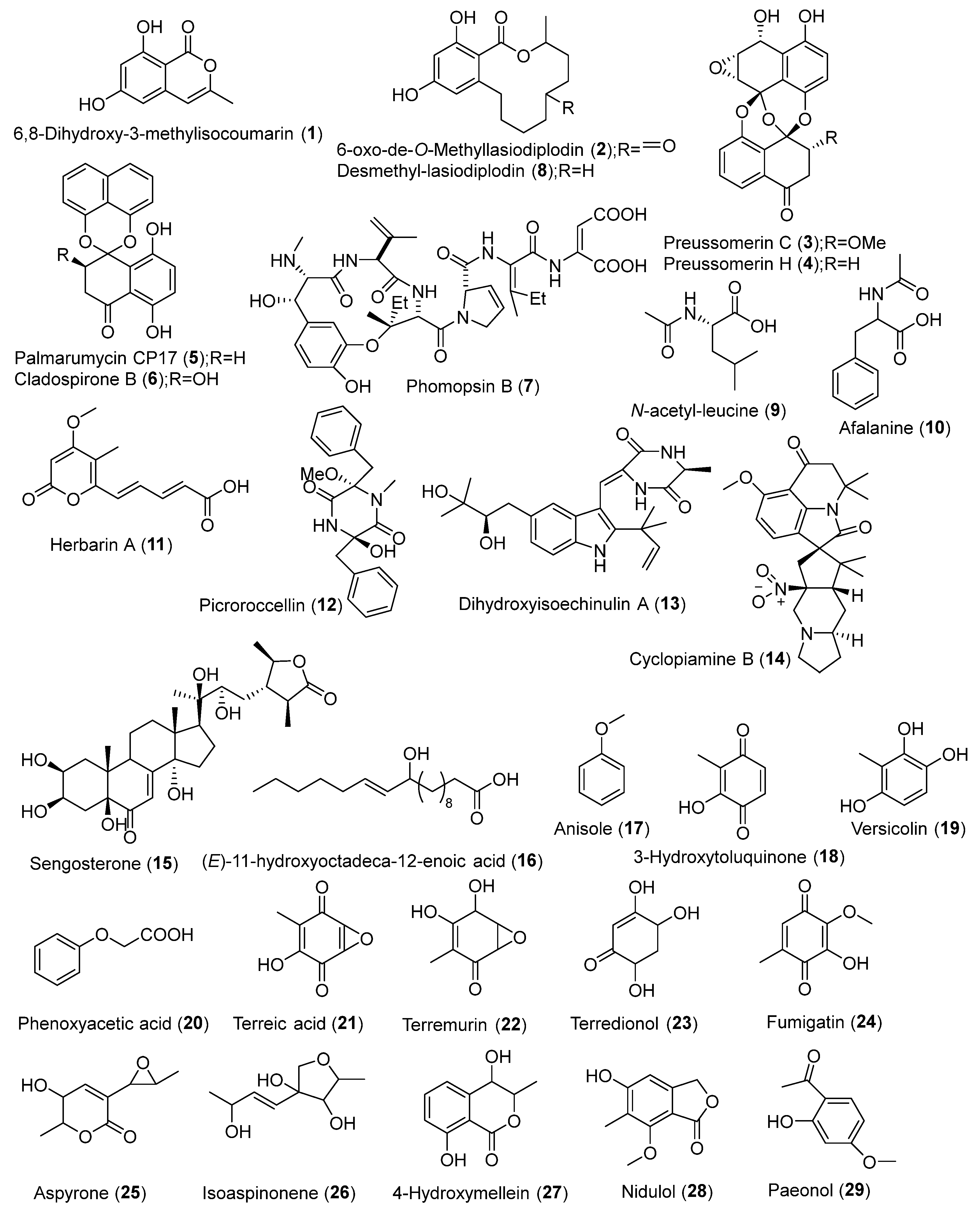
| Aspergillus terreus (AFL, AFSt, AFR) | Artemisia annua, Medicago sativa | UAE | Ethyl acetate | LC-HRMS | MarinLit, Dictionary of Natural Products | Paeonol, p-hydroxy benzoic acid, p-coumaric acid, dihydrosinapic acid, osmundacetone, shikimic acid, parvulenone, nidulol, tyrosol, asperpanoid A, maltoryzine, isopestacin, globoscinic acid, 5,7-dihydroxy-4-methylcoumarin, β-methylumbelliferone, hymecromone, 7-hydroxycoumarin, scopoletin, citropten, similanpyrone A, flavipin, gliotoxin, isotryptoquivaline, neoxaline, ochratoxin B, indole-3-acetic acid, phenethylamine, gregatin A, aflatoxin B1, aflatoxin B1 exo-8,9-epoxide, penicillic acid, terrein, physcion | [2] |
| Aspergillus terreus, A. favus, A. oryzae, Penicillium commune, P. chrysogenum, P. chrysogenum, Talaromyces piophilus, T. piophilus, Fusarium oxysporum, F. nematophilum, Pleosporaceae sp. | Artemisia annua | UAE | Ethyl acetate | LC-HRMS | Dictionary of Natural Products | Physcion, emodin, katenarin, norjavanicin, dechlorogriseofulvin, benzyl benzoate, 4-hydroxy benzyl benzoate, benzyl anisate | [9] |
| Xylaria ellisii sp. nov. | Vaccinium angustifolium | LLE | Ethyl acetate; Methanol/acetone (1:1) | LC-HRMS | - | Griseofulvin, dechlorogriseofulvin, cytochalasin D, zygosporin E, epoxycytochalasin D, hirsutatin A, piliformic acid, 2,3-dihydro,2,4- dimethylbenzofuran-7-carboxylic acid, cyclic pentapeptide 1 and 2, xylarotide A, ellisiiamides A-H | [13] |
| Lasiodiplodia theobromae | Vitex pinnata | LLE | Ethyl acetate | LC-HRMS | AntiBase, Dictionary of Natural Products | 6,8-Dihydroxy-3-methylisocoumarin, 6-oxo-de-O-methyllasiodiplodin, preussomerins-C and H, palmarumycin CP17, cladospirone B, phomopsin B, desmethyl-lasiodiplodin | [18] |
| Curvularia sp. | Terminalia laxiflora | LLE | Ethyl acetate | LC-HRMS | Natural Product Database | N-Acetyl-leucine, afalanine, herbarin A, picroroccellin, dihydroxyisoechinulin A, cyclopiamine B, sengosterone, (E)-11-hydroxyoctadeca-12-enoic acid | [19] |
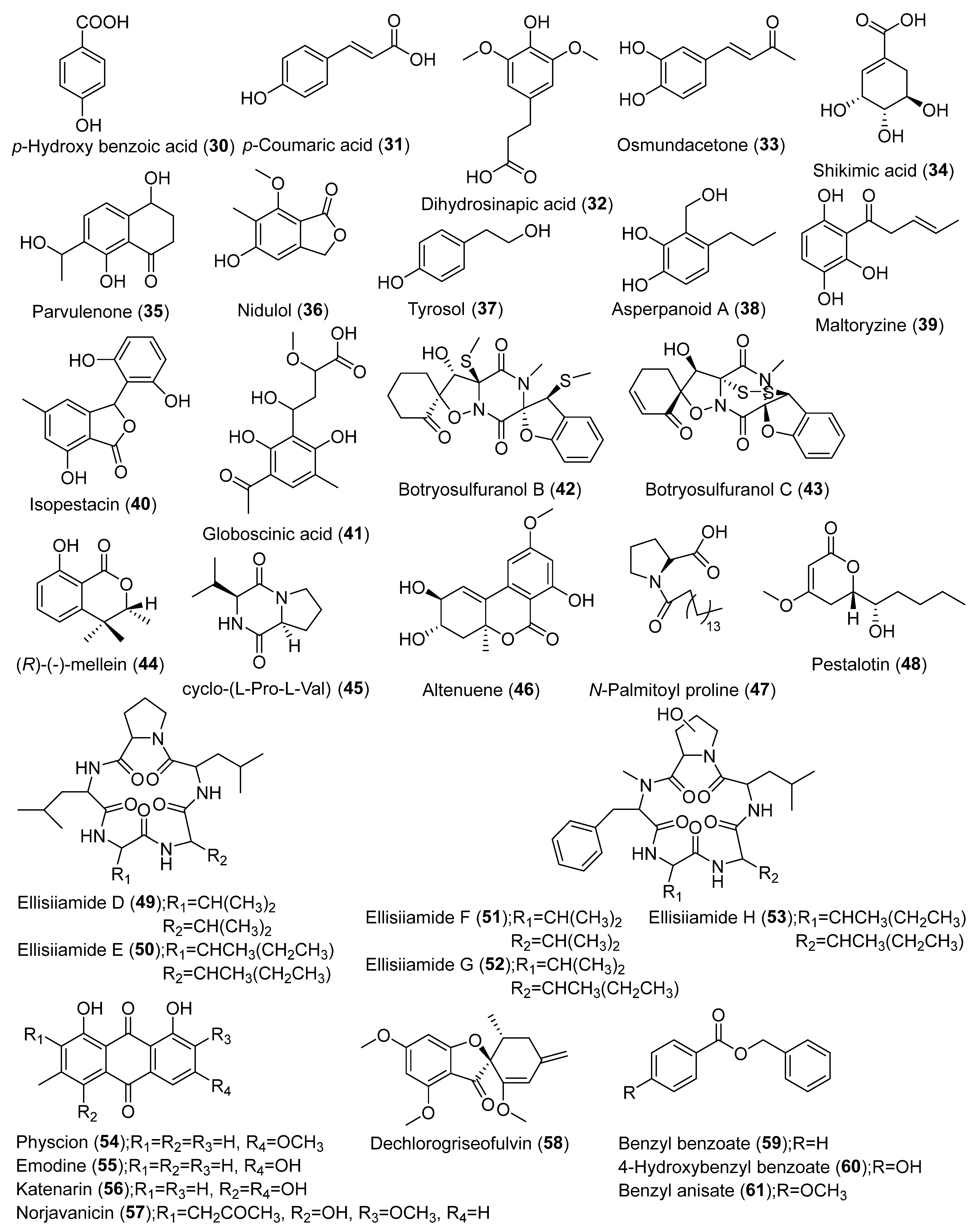
| Aspergillus ochraceus MSEF6 | Medicago sativa | LLE | Ethyl acetate | LC-HRMS | Dictionary of Natural Products, METLIN | Anisole, 3-hydroxytoluquinone, versicolin, phenoxyacetic acid, terreic acid, terremurin, terredionol, fumigatin, aspyrone, isoaspinonene, 4-hydroxymellein, nidulol, aspyrone | [20] |
| Aspergillus terreus GMEF1 | Glycine max L. | UAE | Ethyl acetate | LC-HRMS | Dictionary of Natural Products | Terreic acid, terremutin, (-)-terredionol, terremutin hydrate, 3-methylorsellinic acid, flavipin, astepyrone, reticulol, (3S,6S)-terramide A, emodin, terrelactone A, aspergiketal, 4-hydroxykigelin, 8-hydroxyquadrone, dihydrocitrinone, aspergillide B1, sulochrin, 3α-hydroxy-3,5-dihydromonacolin L | [21] |
| Penicillium setosum | Withania somnifera | UAE/LLE | Dichloromethane: ethyl acetate: methanol (3:2:1)/Ethyl acetate | LC-Q-TOF-MS | METLIN | Kaempferol, quercetin, quercetin acetate, luteolin, dihydroqueretin, dihydromyricetin, quinalizarin, isofraxidin, andrastin D, citromycetin, patulin, 6-deoxyerythronolide B, vanillic acid, 2-dehydro-3-deoxy-darabino-heptonate 7-phosphate (DAHP) | [22] |
| Coohinforma mamane, Fusarium solani | C. mamane (from Bixa orellana L.), F. solani (from Plantago lanceolata) | UAE | Dichloromethane, methanol and water | UHPLC-HRMS | Dictionary of Natural Products, SciFinder, MS Finder, Natural Product Database, KNApSAcK, Chemical Entities of Biological Interest (ChEBI), STOFF, The Toxin and Toxin Target Database (T3DB), Northern African Natural Products Database (NANPDB), Drugbank, FooDB, PlantCyc | Cyclosporins A and E, botryosulfuranol C and B, cyclo-(L-Pro-L-Val), (R)-(-)-mellein, cyclo-(L-Leu-L-Leu-D-Leu-L-Leu-L-Val) | [23] |
| Colletotrichum sp., Diaporthe sp., Periconia sp. | Crescentia alata Kunth | UAE/LLE | Methanol/Ethyl acetate | 1H-NMR | - | Terpenes, phenolics, alkaloids, pigments, steroids, polyketides, glycosides | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagarajan, K.; Ibrahim, B.; Ahmad Bawadikji, A.; Lim, J.-W.; Tong, W.-Y.; Leong, C.-R.; Khaw, K.Y.; Tan, W.-N. Recent Developments in Metabolomics Studies of Endophytic Fungi. J. Fungi 2022, 8, 28. https://doi.org/10.3390/jof8010028
Nagarajan K, Ibrahim B, Ahmad Bawadikji A, Lim J-W, Tong W-Y, Leong C-R, Khaw KY, Tan W-N. Recent Developments in Metabolomics Studies of Endophytic Fungi. Journal of Fungi. 2022; 8(1):28. https://doi.org/10.3390/jof8010028
Chicago/Turabian StyleNagarajan, Kashvintha, Baharudin Ibrahim, Abdulkader Ahmad Bawadikji, Jun-Wei Lim, Woei-Yenn Tong, Chean-Ring Leong, Kooi Yeong Khaw, and Wen-Nee Tan. 2022. "Recent Developments in Metabolomics Studies of Endophytic Fungi" Journal of Fungi 8, no. 1: 28. https://doi.org/10.3390/jof8010028
APA StyleNagarajan, K., Ibrahim, B., Ahmad Bawadikji, A., Lim, J. -W., Tong, W. -Y., Leong, C. -R., Khaw, K. Y., & Tan, W. -N. (2022). Recent Developments in Metabolomics Studies of Endophytic Fungi. Journal of Fungi, 8(1), 28. https://doi.org/10.3390/jof8010028