

Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Strain Isolation
2.2. Morphological Description
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogeny
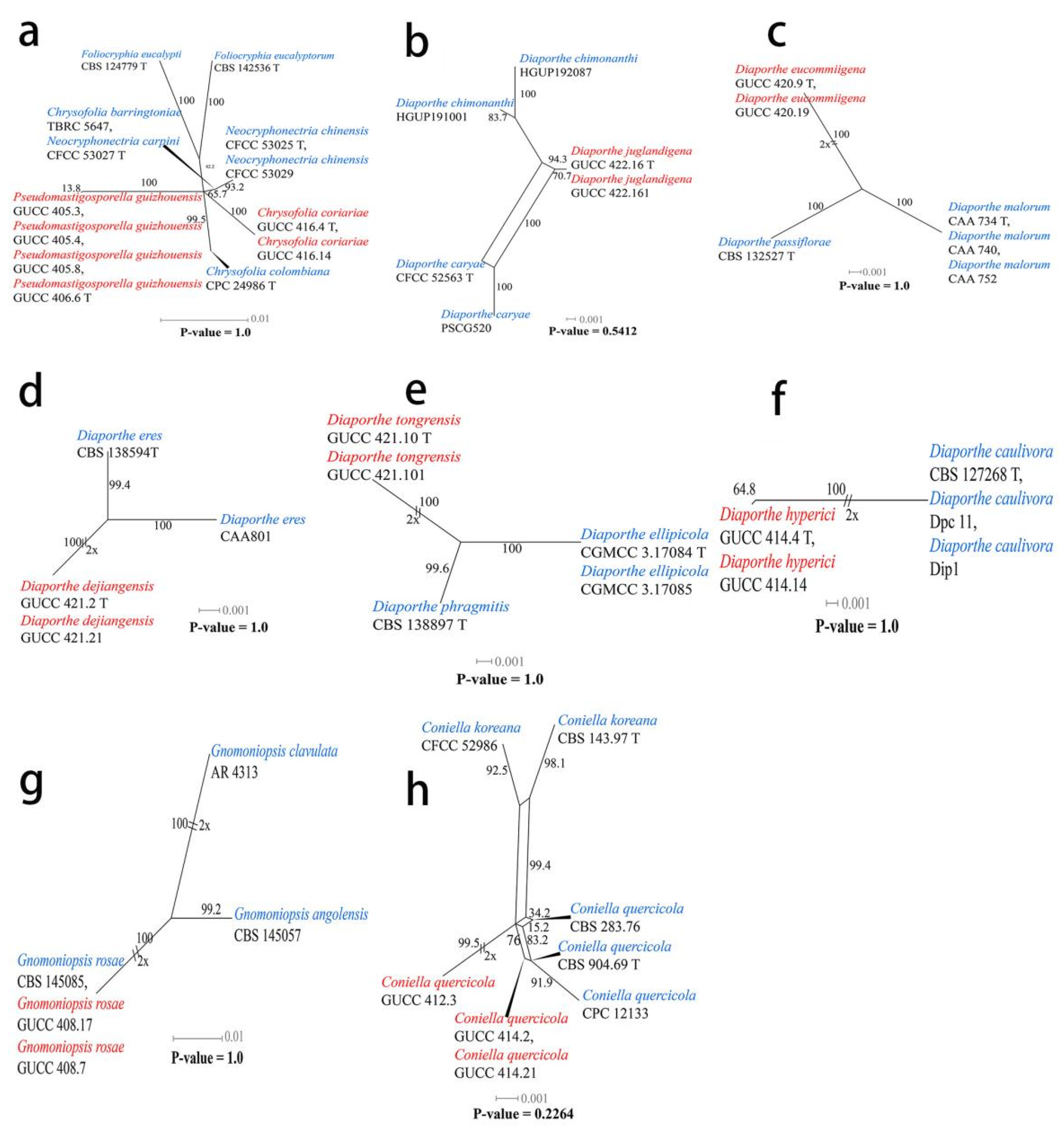
2.5. Genealogical Phylogenetic Species Recognition (GCPSR) Analysis
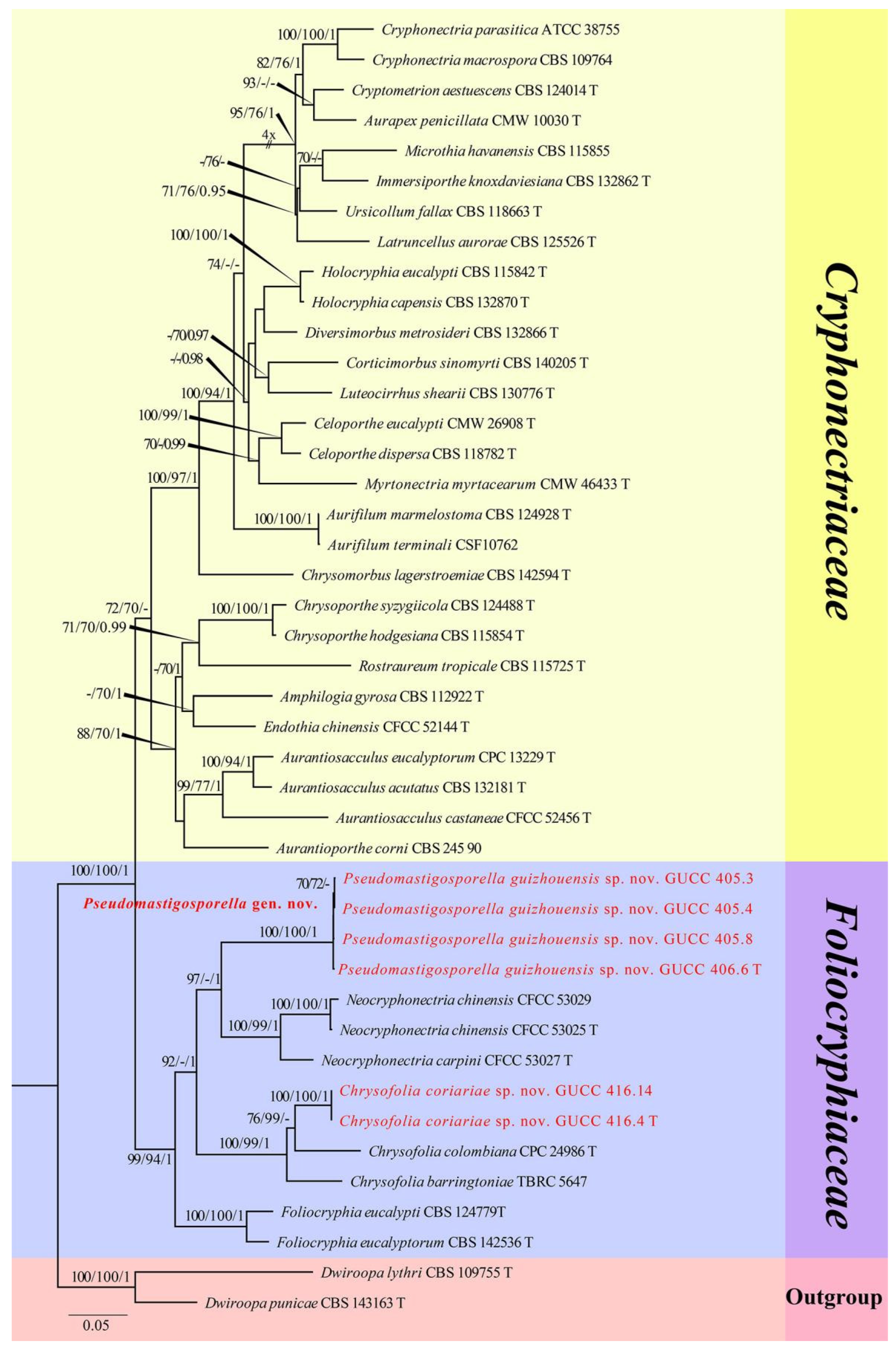
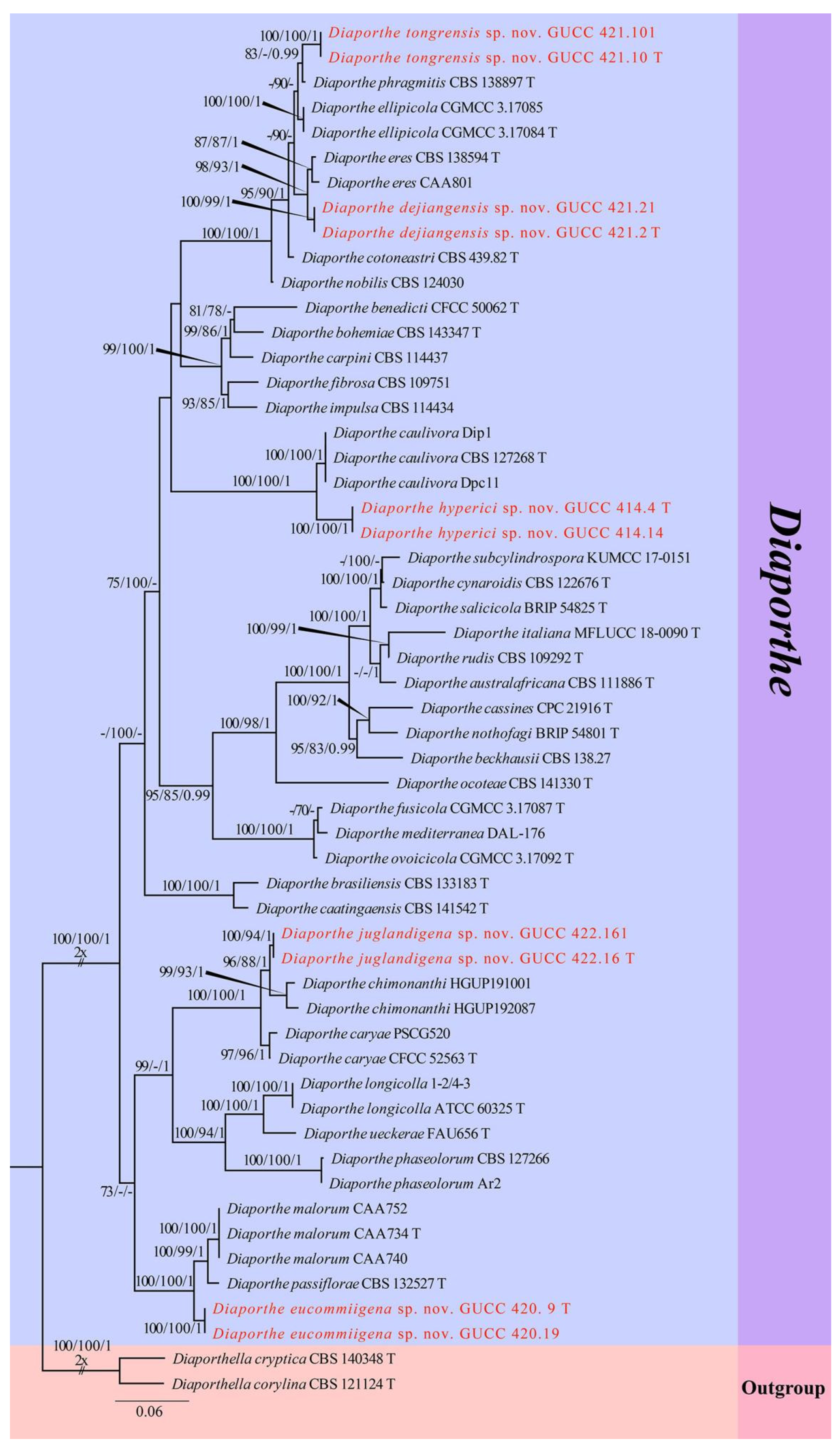
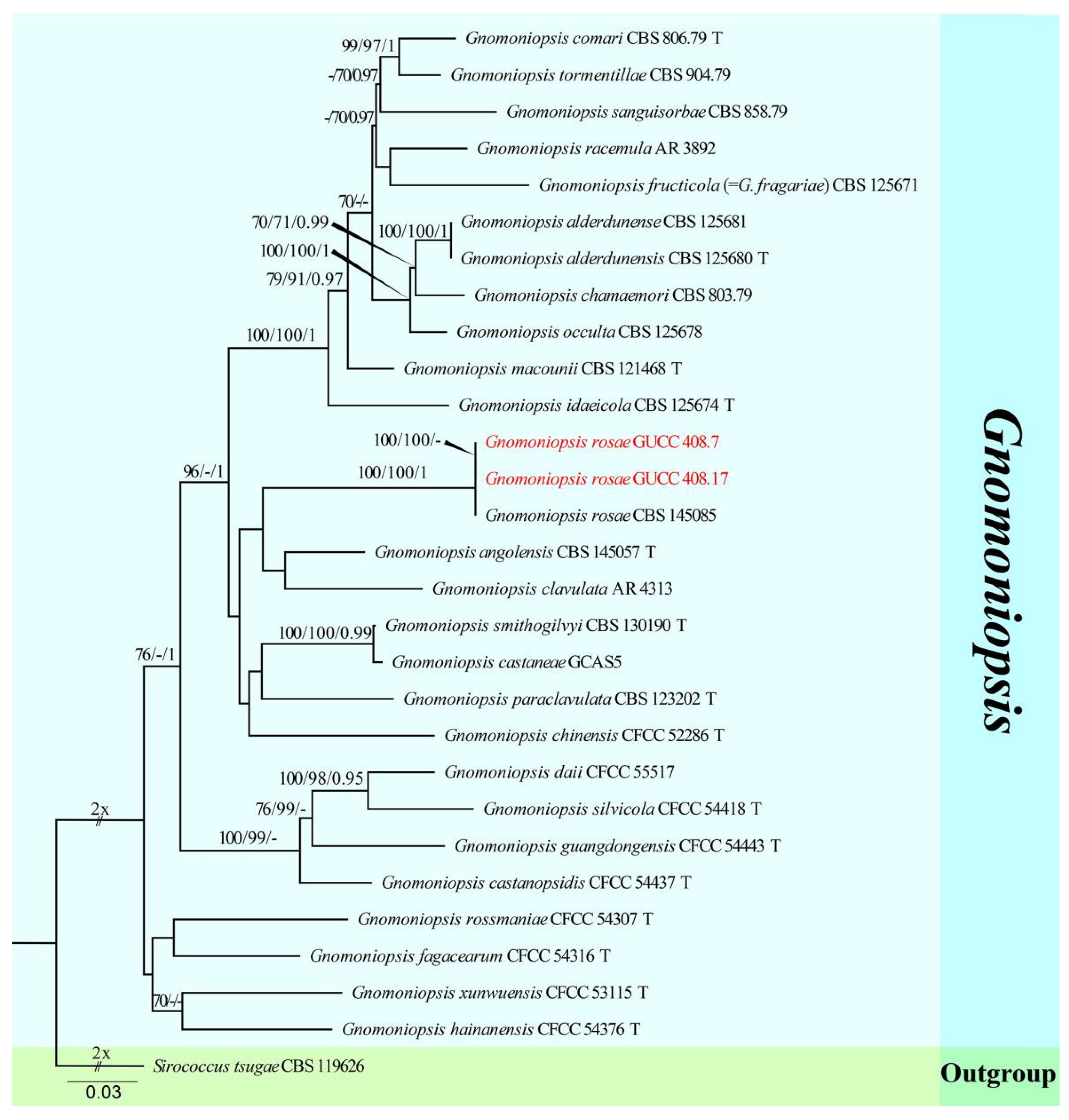
3. Results
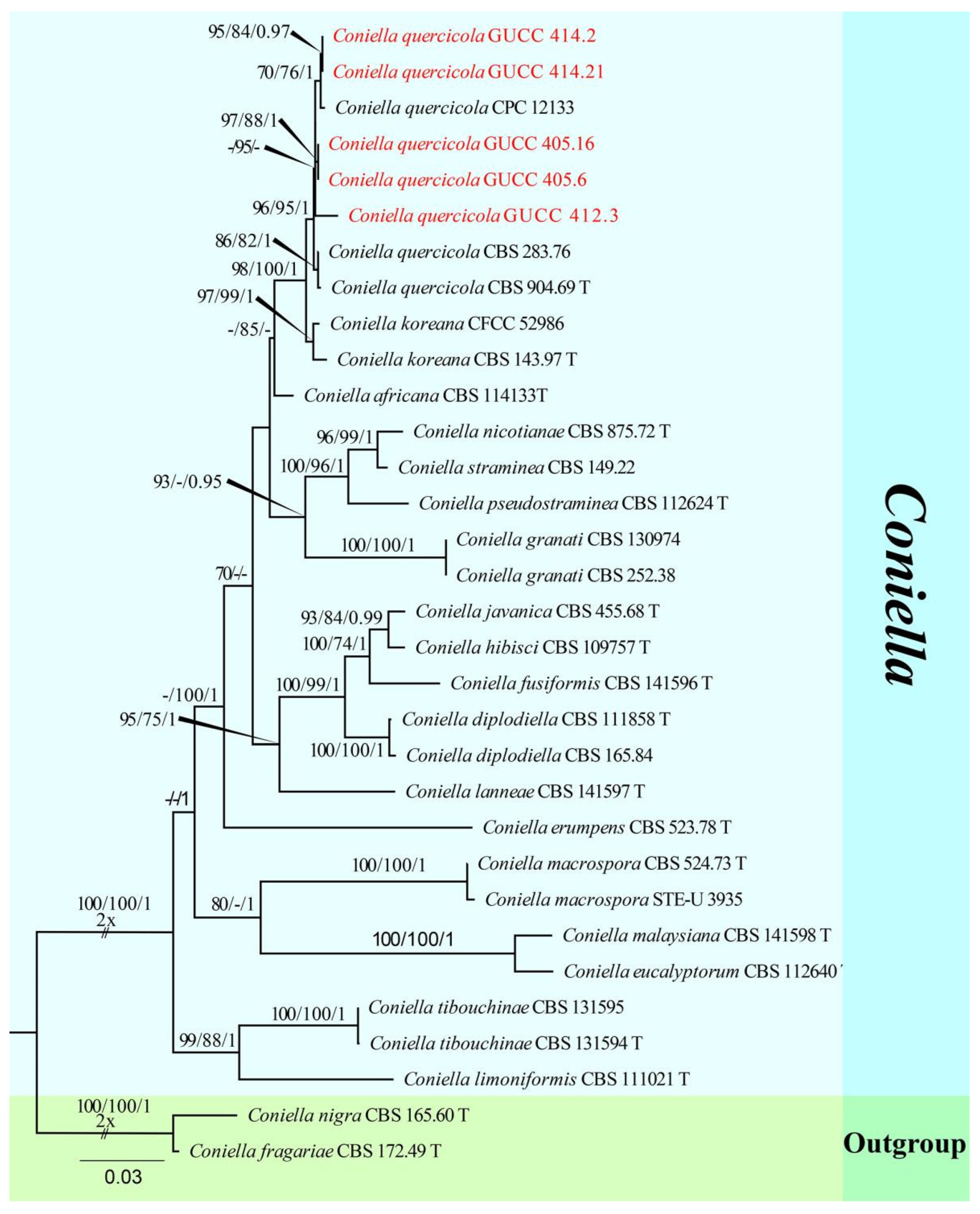
3.1. Phylogenetic Analyses
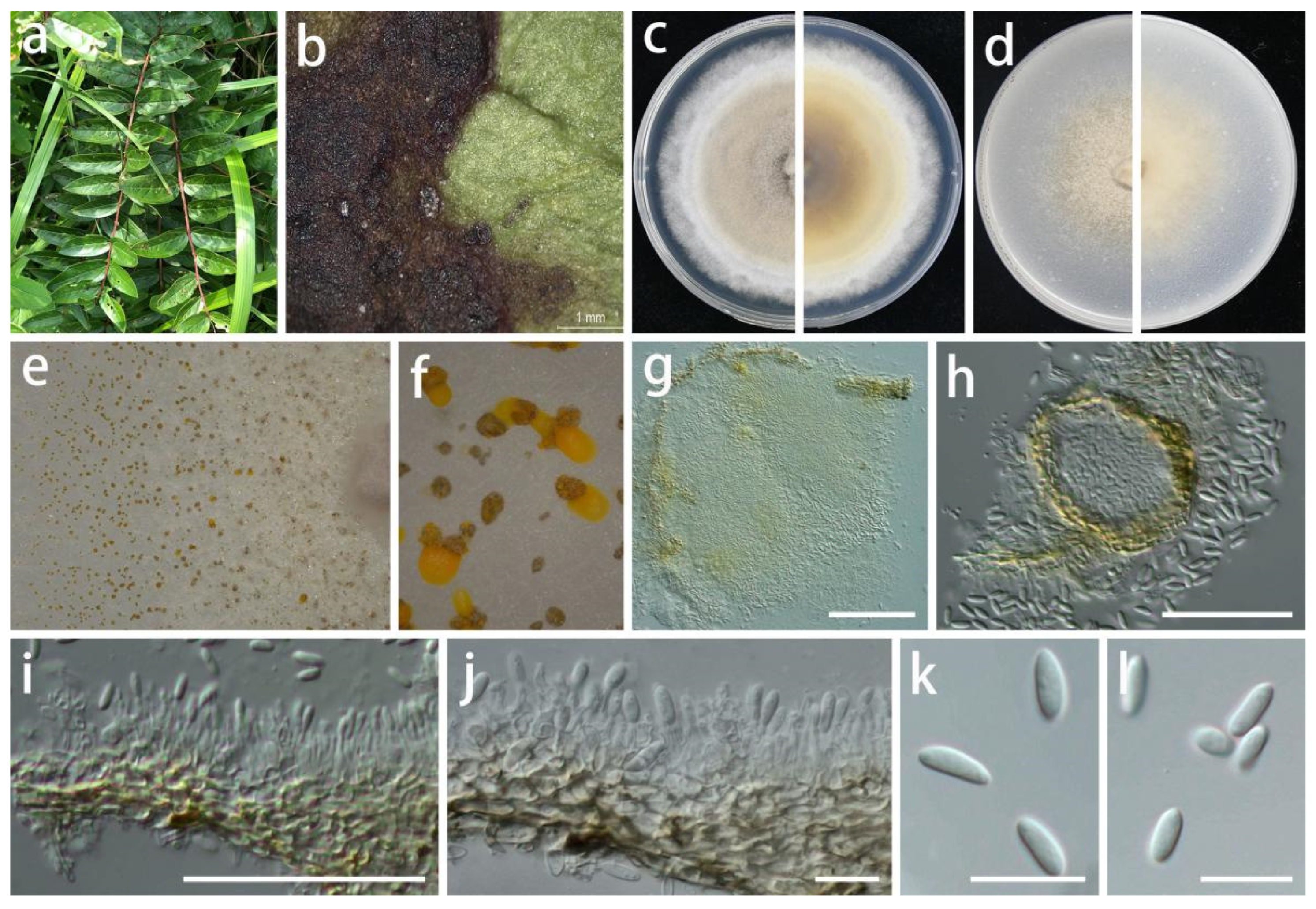
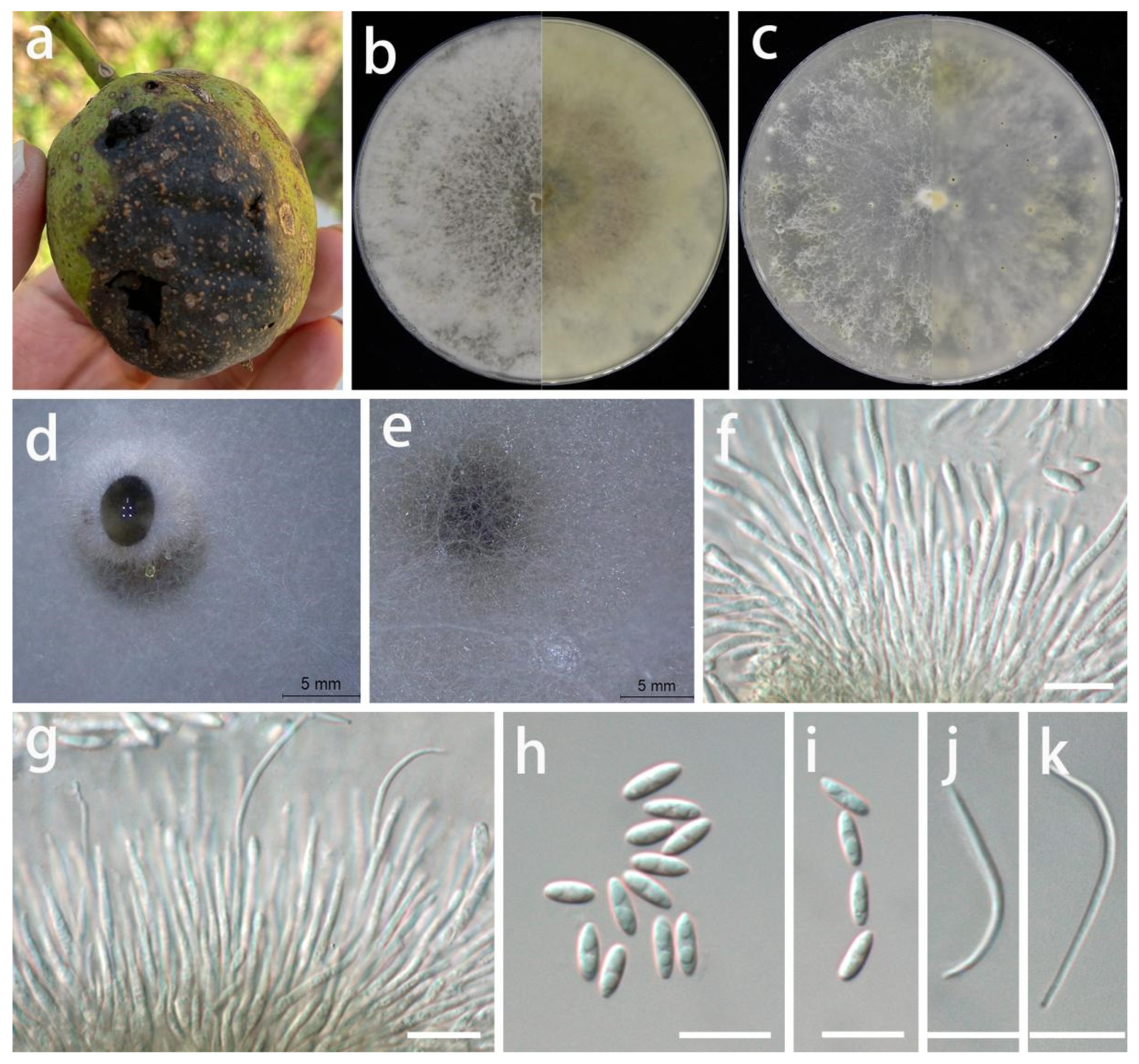
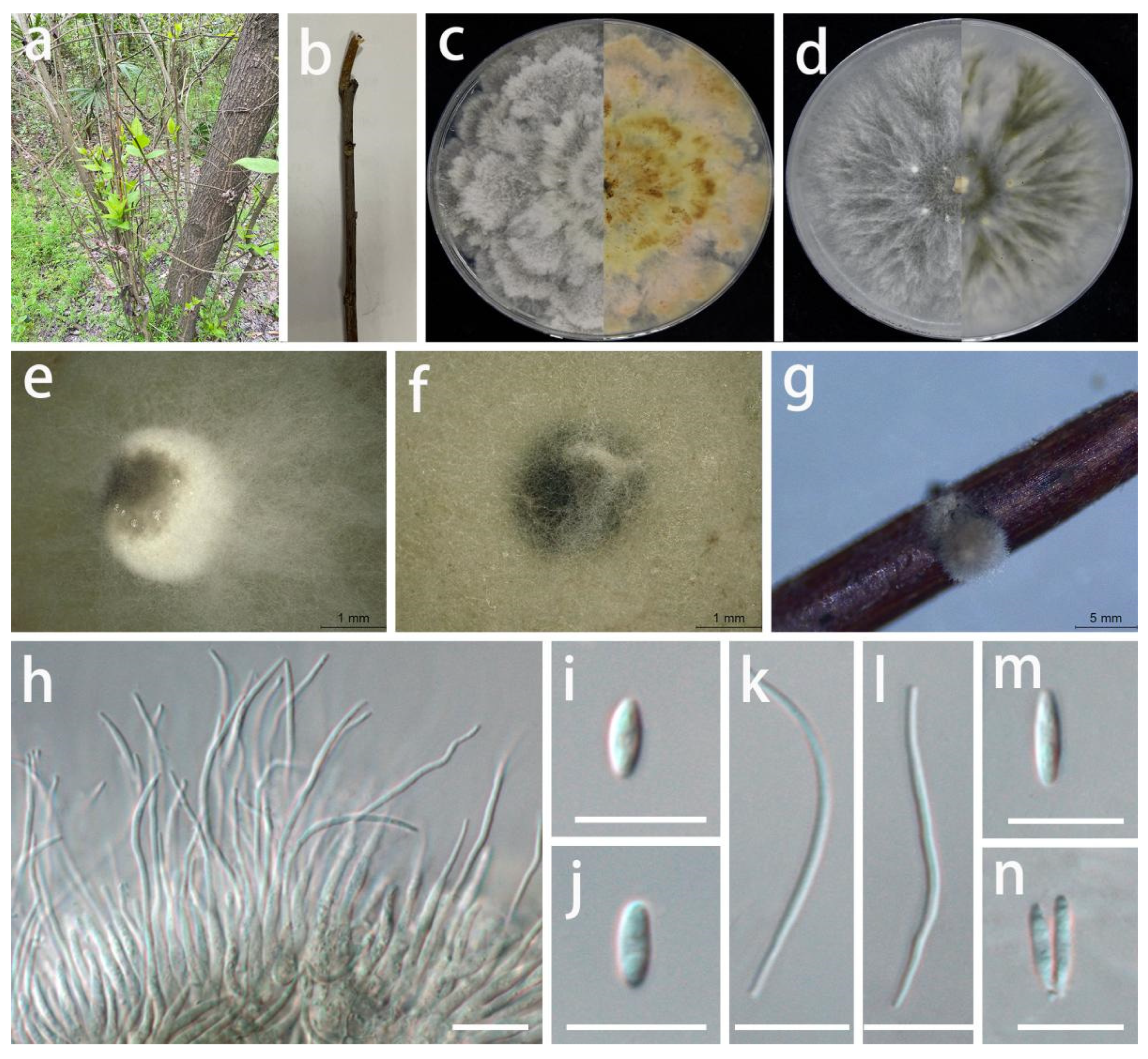
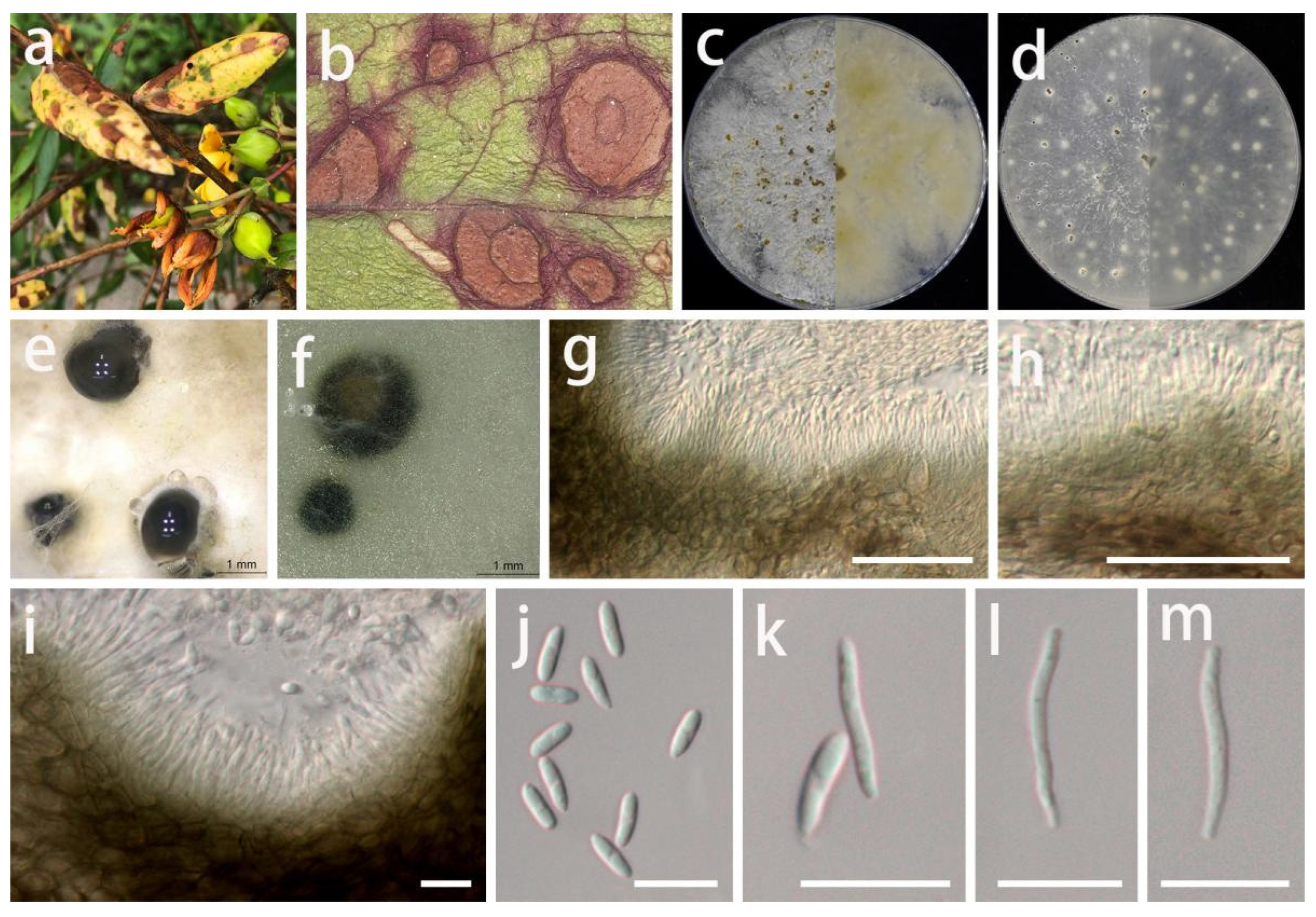
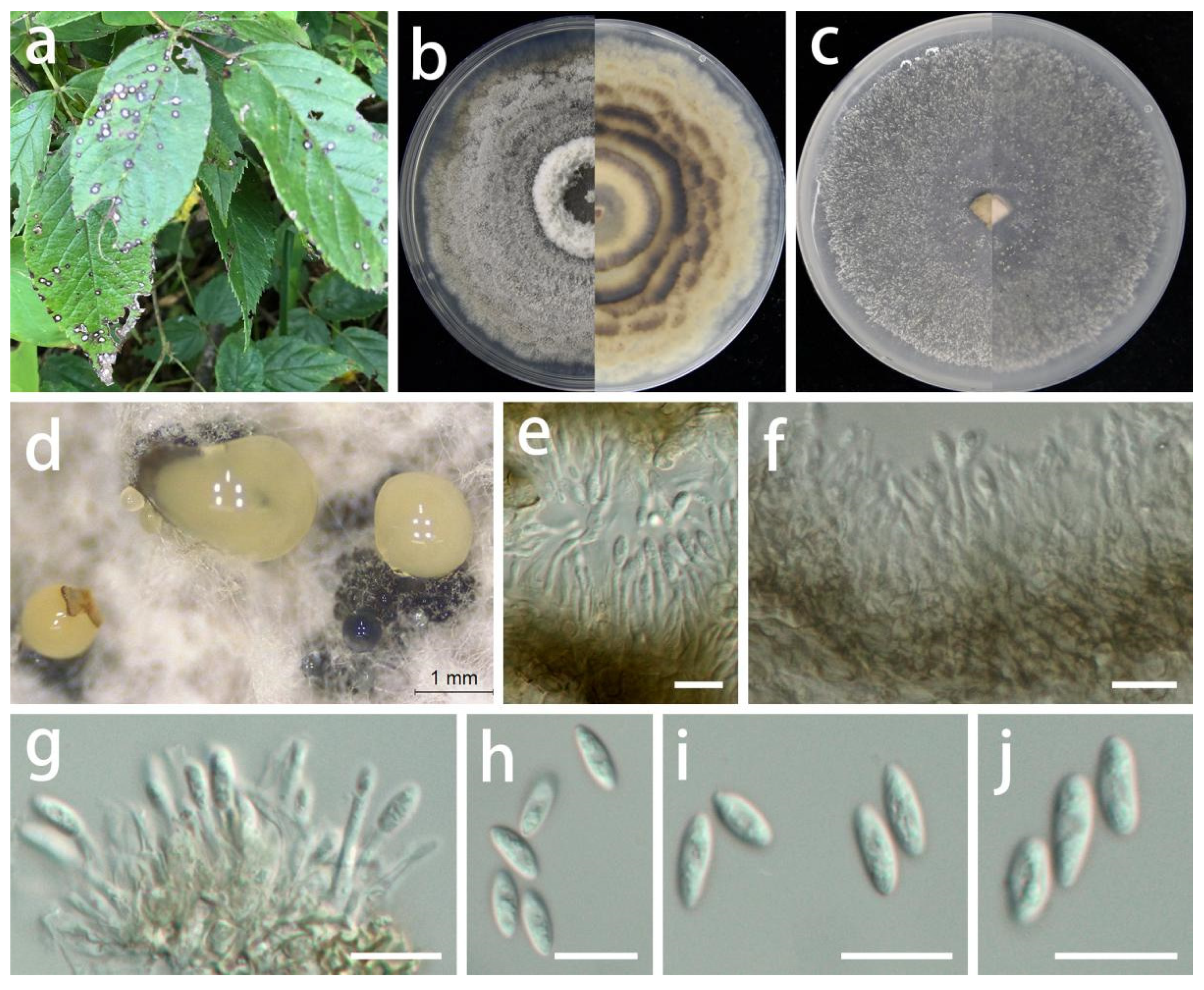
3.2. Taxonomy
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, N.; Fan, X.L.; Tian, C.M.; Crous, P.W. Reevaluating Cryphonectriaceae and allied families in Diaporthales. Mycologia 2020, 112, 267–292. [Google Scholar] [CrossRef] [PubMed]
- Barr, M.E. The Diaporthales in North America: With emphasis on Gnomonia and its segregates. Mycol. Mem. 1978, 7, 1–232. [Google Scholar]
- Alexopoulus, C.J.; Mims, C.W. Introductory Mycology, 3rd ed.; Wiley: New York, NY, USA, 1978. [Google Scholar]
- Castlebury, L.A.; Rossman, A.Y.; Jaklitsch, W.J.; Vasilyeva, L.N. A preliminary overview of the Diaporthales based on large subunit nuclear ribosomal DNA sequences. Mycologia 2002, 94, 1017–1031. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Ainsworth & Bisby’s Dictionary of the Fungi, 8th ed.; CAB International: Oxford, UK, 1995. [Google Scholar]
- Rossman, A.Y.; Farr, D.F.; Castlebury, L.A. A review of the phylogeny and biology of the Diaporthales. Mycoscience 2007, 48, 135–144. [Google Scholar] [CrossRef]
- Qi, P.K.; Jiang, Z.D.; Xiang, M.M. Flora Fungorum Sinicorum: Phomopsis; Science Press: Beijing, China, 2007; Volume 34, pp. 1–185. [Google Scholar]
- Fu, C.H.; Hsieh, H.M.; Chen, C.Y.; Chang, T.T.; Huang, Y.M.; Ju, Y.M. Ophiodiaporthe cyatheae gen. et sp. nov., a diaporthalean pathogen causing a devastating wilt disease of Cyathea lepifera in Taiwan. Mycologia 2013, 105, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.Q.; Wijayawardene, N.N.; Bhat, D.J.; Chukeatirote, E.; Bahkali, A.H.; Zhao, R.L.; Xu, J.C.; Hyde, K.D. Pustulomyces gen. nov. accommodated in Diaporthaceae, Diaporthales, as revealed by morphology and molecular analyses. Cryptogam. Mycol. 2014, 35, 63–72. [Google Scholar] [CrossRef]
- Du, Z.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Tian, C.M. Melansporellaceae: A novel family of Diaporthales (Ascomycota). Phytotaxa 2017, 305, 191–200. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China, with twelve new species described. MycoKeys 2018, 39, 97–149. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Bezerra, J.D.; Tian, C.M.; Crous, P.W. Families and genera of diaporthalean fungi associated with canker and dieback of tree hosts. Persoonia 2018, 40, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Yang, Q.; Fan, X.; Tian, C. Micromelanconis kaihuiae gen. et sp. nov. a new diaporthalean fungus from Chinese chestnut branches in southern China. MycoKeys 2021, 79, 1–16. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Promputtha, I.; Liu, J. Dendrostoma covidicola sp. nov. (Erythrogloeaceae, Diaporthales) on Fagus sylvatica from Sichuan Province, China. Phytotaxa 2021, 483, 85–94. [Google Scholar] [CrossRef]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 609387. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Xia, J.W.; Pan, Y.; Zhu, C.Y.; Li, Z. Morphological and molecular description of Diaporthe nannuoshanensis sp. nov. (Diaporthaceae, Diaporthales) in south-western China. Nova Hedwig. 2021, 112, 399–411. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Santos, L.; Phillips, A.J.L.; Crous, P.W.; Alves, A. Diaporthe species on Rosaceae with descriptions of D. pyracanthae sp. nov. and D. malorum sp. nov. Mycosphere 2017, 8, 485–511. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Molecular systematics of the Hypocreales: A teleomorph gene phylogeny and the status of their anamorphs. Can. J. Bot. 1990, 73, 816–823. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G. Development of primer sets designed for use with PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.; Jeewon, R.; Phillips, A.J.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Jeewon, R.; Chomnunti, P.; Wanasinghe, D.N.; Norphanphoun, C.; Karunarathna, A.; Pem, D.; Perera, R.H.; Camporesi, E.; McKenzie, E.H.C.; et al. Taxonomic circumscription of Diaporthales based on multigene phylogeny and morphology. Fungal Divers. 2018, 93, 241–443. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-Ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal planet description sheets 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.; Huang, S.K.; Abdel-Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’souza, M.J.; Goonasekara, I.D.; et al. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.-K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A.; et al. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Raudabaugh, D.B.; Iturriaga, T.; Carver, A.; Mondo, S.; Pangilinan, J.; Lipzen, A.; He, G.; Amirebrahimi, M.; Grigoriev, I.V.; Miller, A.N. Coniella lustricola, a new species from submerged detritus. Mycol. Prog. 2018, 17, 191–203. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Kuo, C.-H.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Sym. Series 1999, 41, 95–98. [Google Scholar]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 22, 3276–3278. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceeding of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Mark, V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY: A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rannala, B.; Yang, Z.H. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree-Version 1.4.3. A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 5 October 2016).
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Hespanhol, L.; Vallio, C.S.; Costa, L.M.; Saragiotto, B.T. Understanding and interpreting confidence and credible intervals around effect estimates. Braz. J. Phys. Ther. 2019, 23, 290–301. [Google Scholar] [CrossRef]
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Restrepo, M.H.; Sutton, D.A.; Acharya, K.; Barber, P.; Boekhout, T.; Dimitrov, R.A.; Dueñas, M.; et al. Fungal planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Cheewangkoon, R.; Groenewald, J.Z.; Summerell, B.A.; Hyde, K.D.; To-anun, C.; Crous, P.W. Myrtaceae, a cache of fungal biodiversity. Persoonia 2009, 23, 55–85. [Google Scholar] [CrossRef] [Green Version]
- Suwannarach, N.; Kumla, J.; Sri-Ngernyuang, K.; Lumyong, S. A new endophytic fungus, Chrysofolia barringtoniae sp. nov. from Thailand. Mycoscience 2016, 57, 361–365. [Google Scholar] [CrossRef]
- Chang, C.Q.; Xi, P.G.; Xiang, M.M.; Jiang, Z.D.; Chi, P.K. New species of Phomopsis on woody plants in Hunan province. Mycosystema 2005, 24, 145–154. [Google Scholar]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Groenewald, J.Z. Fungal planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; Crous, P.W.; McKenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Restrepo, M.H.; Sutton, D.A.; Acharya, K.; Barber, P.; Boekhout, T.; Dimitrov, R.A.; Dueñas, M.; et al. Fungal Planet Description Sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef]
- Santos, J.; Vrandečić, K.; Ćosić, J.; Duvnjak, T.; Phillips, A. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef]
- Sogonov, M.V.; Castlebury, L.A.; Rossman, A.Y.; Mejía, L.C.; White, J.F. Leaf-inhabiting genera of the Gnomoniaceae, Diaporthales. Stud. Mycol. 2008, 62, 1–77. [Google Scholar] [CrossRef]
- Walker, D.M.; Castlebury, L.A.; Rossman, A.Y.; Sogonov, M.V.; White, J.F. Systematics of genus Gnomoniopsis (Gnomoniaceae, Diaporthales) based on a three gene phylogeny, host associations and morphology. Mycologia 2010, 102, 1479–1496. [Google Scholar] [CrossRef]
- Saccardo, P.A.; Saccardo, D. Supplementum universale, Pars VII. Discomycetae-Deuteromycatae. Syll. Fung. 1906, 18, 1–838. [Google Scholar]
- Alvarez, L.V.; Groenewald, J.Z.; Crous, P.W. Revising the Schizoparmaceae: Coniella and its synonyms Pilidiella and Schizoparme. Stud. Mycol. 2016, 85, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Tian, C.M. An emerging pathogen from rotted chestnut in China: Gnomoniopsis daii sp. nov. Forests 2019, 10, 1016. [Google Scholar] [CrossRef] [Green Version]
- Voglmayr, H.; Castlebury, L.A.; Jaklitsch, W.M. Juglanconis gen. nov. on Juglandaceae, and the new family Juglanconidaceae (Diaporthales). Persoonia 2017, 38, 136–155. [Google Scholar] [CrossRef] [Green Version]
- Voglmayr, H.; Jaklitsch, W.M. Stilbosporaceae resurrected: Generic reclassification and speciation. Persoonia 2014, 33, 61–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voglmayr, H.; Rossman, A.Y.; Castlebury, L.A.; Jaklitsch, W.M. Multigene phylogeny and taxonomy of the genus Melanconiella (Diaporthales). Fungal Divers. 2012, 57, 1–44. [Google Scholar] [CrossRef]
- Suetrong, S.; Klaysuban, A.; Sakayaroj, J.; Preedanon, S.; Ruang-Areerate, P.; Phongpaichit, S.; Pang, K.-L.; Jones, E. Tiarisporellaceae, a new family in the order Diaporthales (Sordariomycetes, Ascomycota). Crypt. Mycol. 2015, 36, 319–330. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Hongsanan, S.; Doilom, M.; Bhat, D.J.; Wen, T.-C.; Senanayake, I.C.; Bulgakov, T.; Hyde, K.D. Lamproconiaceae fam. nov. to accommodate Lamproconium desmazieri. Phytotaxa 2016, 270, 89–102. [Google Scholar]
- Yang, Q.; Fan, X.L.; Du, Z.; Tian, C.M. Diaporthosporellaceae, a novel family of Diaporthales (Sordariomycetes, Ascomycota). Mycoscience 2018, 59, 229–235. [Google Scholar] [CrossRef]
- Guterres, D.C.; Galvão-Elias, S.; Dos Santos, M.D.D.M.; de Souza, B.C.P.; de Almeida, C.P.; Pinho, D.B.; Miller, R.N.G.; Dianese, J.C. Phylogenetic relationships of Phaeochorella parinarii and recognition of a new family, Phaeochorellaceae (Diaporthales). Mycologia 2019, 111, 660–675. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, N.; Tian, C.M. Three new Diaporthe species from Shaanxi Province, China. MycoKeys 2020, 67, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Zapata, M.; Palma, M.A.; Aninat, M.J.; Piontelli, E. Polyphasic studies of new species of Diaporthe from native forest in Chile, with descriptions of Diaporthe araucanorum sp. nov., Diaporthe foikelawen sp. nov. and Diaporthe patagonica sp. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 3379–3390. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications. species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejía, L.C.; Castlebury, L.A.; Rossman, A.Y.; Sogonov, M.V.; White, J.F. A systematic account of the genus Plagiostoma (Gnomoniaceae, Diaporthales) based on morphology, host-associations, and a four-gene phylogeny. Stud. Mycol. 2011, 68, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Mejía, L.C.; Rossman, A.Y.; Castlebury, L.A.; White, J.F. New species, phylogeny, host-associations and geographic distribution of genus Cryptosporella (Gnomoniaceae, Diaporthales). Mycologia 2011, 103, 379–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejia, L.C.; Rossman, A.Y.; Castlebury, L.A.; Yang, Z.L.; White, J.F. Occultocarpon, a new monotypic genus of Gnomoniaceae on Alnus nepalensis from China. Fungal Divers. 2012, 52, 99–105. [Google Scholar] [CrossRef]
- Walker, D.M.; Castlebury, L.A.; Rossman, A.Y.; Mejía, L.C.; White, J.F. Phylogeny and taxonomy of Ophiognomonia (Gnomoniaceae, Diaporthales), including twenty-five new species in this highly diverse genus. Fungal Divers. 2012, 57, 85–147. [Google Scholar] [CrossRef]
- Monod, M. Taxonomic monograph of the Gnomoniaceae (Ascomycetes of the ordre Diaporthales). I. Beih. Zur Sydowia 1983, 9, 1–315. [Google Scholar]
- Höhnel, F.V. Dritte vorlaufige Mitteilung mycologischer Ergebnisse (Nr. 201–304). Ber. Der Dtsch. Bot. Ges. 1918, 36, 309–317. [Google Scholar]
- Petrak, F.; Sydow, H. Die Gattungen der Pyrenomyzeten, Sphaeropsideen and Melanconieen. I. Der phaeosporen Sphaeropsideen und die Gattung Macrophoma. Feddes Repert. Speciarum Nov. Regni Veg. Beih. 1927, 42, 1–551. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primers | Primer Sequence (5′–3′) | PCR Thermal Cycle Protocols | References |
|---|---|---|---|---|
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG | 94 °C–2 min; 94 °C–1 min; 58 °C–1 min; 72 °C–90 s; repeat 2–4 for 35 cycles; 72 °C–10 min; 4 °C on hold | White et al. [23] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| LSU | LR0R | ACCCGCTGAACTTAAGC | Rehner and Samuels [24]; Vilgalys and Hester [25] | |
| LR5 | TCCTGAGGGAAACTTCG | |||
| rpb2 | fRPB2-5F | GAYGAYMGWGATCAYTTYGG | 94 °C–2 min; 95 °C–45 s; 57 °C–50 s; 72 °C–90 s; repeat 2–4 for 35 cycles; 72 °C–10 min; 4 °C on hold | Liu et al. [26] |
| fRPB2-7cR | CCCATRGCTTGYTTRCCCAT | |||
| tef1 | EF1-728F | CATCGAGAAGTTCGAGAAGG | 94 °C–2 min; 95 °C–30 s; 58 °C–50 s; 72 °C–1 min; repeat 2–4 for 35 cycles; 72 °C–10 min; 4 °C on hold | Carbone and Kohn [27] |
| EF1-986R | TACTTGAAGGAACCCTTACC | |||
| tub2 | Bt2a | GGTAACCAAATCGGTGCTGCTTTC | 95 °C–5 min; 95 °C–30 s; 60 °C–30 s; 72 °C–30 s; repeat 2–4 for 34 cycles; 72 °C–10 min; 4 °C on hold | Glass and Donaldson [28] |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC |
| Species | Strain Number | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | tub2 | ||
| Amphilogia gyrosa | CBS 112922T | AF452111 | AY194107 | MN271782 | MN271818 | AF525714 |
| Aurantioporthe corni | CBS 245.90 | MN172403 | MN172371 | MN271784 | MN271822 | – |
| Aurantiosacculus acutatus | CBS 132181T | JQ685514 | JQ685520 | – | MN271823 | – |
| Aurantiosacculus castaneae | CFCC 52456T | MH514025 | MH514015 | MN271786 | – | MH539688 |
| Aurantiosacculus eucalyptorum | CPC 13229T | JQ685515 | JQ685521 | MN271785 | MN271824 | – |
| Aurapex penicillata | CMW 10030T | AY214311 | AY194103 | MN271787 | – | AY214239 |
| Aurifilum marmelostoma | CBS 124928T | MH863426 | MH874934 | MN271788 | MN271827 | MN987000 |
| Aurifilum terminali | CSF10762 | MN199838 | MN258786 | – | MN258781 | MN258771 |
| Celoporthe dispersa | CBS 118782T | DQ267130 | HQ730853 | – | HQ730840 | AY214280 |
| Celoporthe eucalypti | CMW 26908T | HQ730837 | HQ730863 | MN271790 | HQ730850 | MN263386 |
| Chrysofolia barringtoniae | TBRC 5647 | KU948046 | KU948045 | – | – | – |
| Chrysofolia colombiana | CPC 24986T | KR476738 | KR476771 | – | MN271829 | – |
| Chrysofolia coriariaesp. nov. | GUCC 416.4T | OP581211 | OP581237 | – | OP688516 | OP688542 |
| Chrysofolia coriariaesp. nov. | GUCC 416.14 | OP581212 | OP581238 | – | OP688517 | OP688543 |
| Chrysomorbus lagerstroemiae | CBS 142594T | KY929338 | KY929328 | – | MN271830 | KY929348 |
| Chrysoporthe hodgesiana | CBS 115854T | AY692322 | MN172380 | MN271793 | MN271836 | – |
| Chrysoporthe syzygiicola | CBS 124488T | FJ655005 | MN172383 | – | MN271839 | FJ805236 |
| Corticimorbus sinomyrti | CBS 140205T | KT167169 | KT167179 | MN271794 | MN271841 | KT167183 |
| Cryphonectria macrospora | CBS 109764 | EU199182 | AF408340 | EU220029 | KC465405 | AH014594 |
| Cryphonectria parasitica | ATCC 38755 | MH843497 | MH514021 | DQ862017 | MN271848 | MW086477 |
| Cryptometrion aestuescens | CBS 124014T | MH863335 | MH874864 | MN271798 | MN271851 | GQ369456 |
| Diversimorbus metrosideri | CBS 132866T | JQ862871 | JQ862828 | – | MN271857 | JQ862953 |
| Dwiroopa lythri | CBS 109755T | MN172410 | MN172389 | MN271801 | MN271859 | – |
| Dwiroopa punicae | CBS 143163T | MK510676 | MK510686 | MK510692 | MH020056 | MK510714 |
| Endothia chinensis | CFCC 52144T | MH514027 | MH514017 | – | MN271860 | MH539690 |
| Foliocryphia eucalypti | CBS 124779T | GQ303276 | GQ303307 | MN271802 | MN271861 | JQ706128 |
| Foliocryphia eucalyptorum | CBS 142536T | KY979772 | KY979827 | MN271803 | MN271862 | KY979936 |
| Holocryphia capensis | CBS 132870T | JQ862854 | JQ862811 | – | MN271883 | JQ862948 |
| Holocryphia eucalypti | CBS 115842T | MN172411 | MN172391 | MN271804 | MN271882 | JQ862772 |
| Immersiporthe knoxdaviesiana | CBS 132862T | JQ862765 | JQ862755 | MN271805 | MN271886 | JQ862785 |
| Latruncellus aurorae | CBS 125526T | GU726947 | HQ730872 | – | MN271888 | GU726959 |
| Luteocirrhus shearii | CBS 130776T | KC197021 | KC197019 | MN271807 | MN271890 | KC197006 |
| Microthia havanensis | CBS 115855 | DQ368735 | MN172393 | MN271811 | – | – |
| Myrtonectria myrtacearum | CMW 46433T | MG585736 | MG585750 | – | – | MG585734 |
| Neocryphonectria carpini | CFCC 53027T | MN172413 | MN172396 | – | – | – |
| Neocryphonectria chinensis | CFCC 53025T | MN172414 | MN172397 | MN271812 | MN271893 | – |
| Neocryphonectria chinensis | CFCC 53029 | MN172415 | MN172398 | MN271813 | MN271894 | – |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 406.6T | OP581233 | OP581246 | OP688514 | OP688538 | OP688563 |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 405.3 | OP581234 | OP581247 | OP688515 | OP688539 | OP688564 |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 405.4 | OP581235 | OP581248 | – | OP688540 | OP688565 |
| Pseudomastigosporella guizhouensis sp. nov. | GUCC 405.8 | OP581236 | OP581249 | – | OP688541 | OP688566 |
| Rostraureum tropicale | CBS 115725T | AY167435 | MN172399 | MN271814 | MN271895 | AY167431 |
| Ursicollum fallax | CBS 118663T | DQ368755 | EF392860 | MN271816 | MN271897 | AH015658 |
| Species | Strain Number | GenBank Accession Number | ||
|---|---|---|---|---|
| ITS | tef1 | tub2 | ||
| Diaporthe australafricana | CBS 111886T | KC343038 | KC343764 | KC344006 |
| D. beckhausii | CBS 138.27 | KC343041 | KC343767 | KC344009 |
| D. benedicti | CFCC 50062T | KP208847 | KP208853 | KP208855 |
| D. bohemiae | CBS 143347T | MG281015 | MG281536 | MG281188 |
| D. brasiliensis | CBS 133183T | KC343042 | KC343768 | KC344010 |
| D. caatingaensis | CBS 141542T | KY085927 | KY115603 | KY115600 |
| D. carpini | CBS 114437 | KC343044 | KC343770 | KC344012 |
| D. caryae | CFCC 52563T | MH121498 | MH121540 | MH121580 |
| D. caryae | PSCG520 | MK626952 | MK654895 | MK691315 |
| D. cassines | CPC 21916T | KF777155 | KF777244 | – |
| D. caulivora | CBS 127268T | KC343045 | KC343771 | KC344013 |
| D. caulivora | Dip1 | HM347703 | HM347687 | – |
| D. caulivora | Dpc11 | HM347704 | HM347688 | – |
| D. chimonanthi | HGUP191001 | MZ724752 | – | MZ724033 |
| D. chimonanthi | HGUP192087 | MZ724753 | – | MZ724034 |
| D. cotoneastri | CBS 439.82T | MH861511 | GQ250341 | JX275437 |
| D. cynaroidis | CBS 122676T | KC343058 | KC343784 | KC344026 |
| D. dejiangensis sp. nov. | GUCC 421.2T | OP581221 | OP688526 | OP688551 |
| D. dejiangensis sp. nov. | GUCC 421.21 | OP581222 | OP688527 | OP688552 |
| D. ellipicola | CGMCC 3.17084T | KF576270 | KF576245 | KF576294 |
| D. ellipicola | CGMCC 3.17085 | KF576271 | KF576246 | KF576295 |
| D. eres | CBS 138594T | KJ210529 | KJ210550 | KJ420799 |
| D. eres | CAA801 | KY435644 | KY435631 | KY435672 |
| D. eucommiigenasp. nov. | GUCC 420.9T | OP581223 | OP688528 | OP688553 |
| D. eucommiigenasp. nov. | GUCC 420.19 | OP581224 | OP688529 | OP688554 |
| D. fibrosa | CBS 109751 | KC343099 | KC343825 | KC344067 |
| D. fusicola | CGMCC 3.17087T | KF576281 | KF576256 | KF576305 |
| D. hypericisp. nov. | GUCC 414.4T | OP581227 | OP688532 | OP688557 |
| D. hypericisp. nov. | GUCC 414.14 | OP581228 | OP688533 | OP688558 |
| D. impulsa | CBS 114434 | KC343121 | KC343847 | KC344089 |
| D. italiana | MFLUCC 18-0090T | MH846237 | MH853686 | MH853688 |
| D. juglandigena sp. nov. | GUCC 422.16T | OP581229 | OP688534 | OP688559 |
| D. juglandigena sp. nov. | GUCC 422.161 | OP581230 | OP688535 | OP688560 |
| D. longicolla | ATCC 60325T | KJ590728 | KJ590767 | KJ610883 |
| D. longicolla | 1-2/4-3 | HM347711 | HM347682 | – |
| D. malorum | CAA734T | KY435638 | KY435627 | KY435668 |
| D. malorum | CAA740 | KY435642 | KY435629 | KY435670 |
| D. malorum | CAA752 | KY435643 | KY435630 | KY435671 |
| D. mediterranea | DAL-176 | MT007496 | MT006996 | MT006693 |
| D. nobilis | CBS 124030 | KC343149 | KC343875 | KC344117 |
| D. nothofagi | BRIP 54801T | JX862530 | JX862536 | KF170922 |
| D. ocoteae | CBS 141330T | KX228293 | – | KX228388 |
| D. ovoicicola | CGMCC 3.17092T | KF576264 | KF576239 | KF576288 |
| D. passiflorae | CBS 132527T | JX069860 | KY435633 | KY435674 |
| D. phaseolorum | Ar2 | HM347705 | HM347679 | – |
| D. phaseolorum | CBS 127266 | HM347707 | HM347672 | HQ333513 |
| D. phragmitis | CBS 138897T | KP004445 | – | KP004507 |
| D. rudis | CBS 109292T | KC343234 | KC343960 | KC344202 |
| D. salicicola | BRIP 54825T | JX862531 | JX862537 | KF170923 |
| D. subcylindrospora | KUMCC 17-0151 | MG746629 | MG746630 | MG746631 |
| D. tongrensissp. nov. | GUCC 421.10T | OP581225 | OP688530 | OP688555 |
| D. tongrensissp. nov. | GUCC 421.101 | OP581226 | OP688531 | OP688556 |
| D. ueckerae | FAU656T | KJ590726 | KJ590747 | KJ610881 |
| Diaporthella corylina | CBS 121124T | KC343004 | KC343730 | KC343972 |
| Diaporthella cryptica | CBS 140348T | MN172409 | MN271854 | – |
| Species | Strain Number | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | tub2 | ||
| Gnomoniopsis alderdunense | CBS 125681 | GU320827 | MH875098 | – | GU320802 | GU320789 |
| G. alderdunensis | CBS 125680T | MH863625 | MH875097 | – | GU320801 | GU320787 |
| G. angolensis | CBS 145057T | MK047428 | MK047479 | MK047539 | – | – |
| G. castaneae | GCAS5 | MH107830 | MZ682110 | – | MH213486 | MH213481 |
| G. castanopsidis | CFCC 54437T | MZ902909 | – | – | MZ936385 | |
| G. chamaemori | CBS 803.79 | EU254808 | EU255107 | – | GU320809 | EU219155 |
| G. chinensis | CFCC 52286T | MG866032 | – | – | MH545370 | MH545366 |
| G. clavulata | AR 4313 | EU254818 | – | EU219251 | EU221934 | EU219211 |
| G. comari | CBS 806.79T | EU254821 | EU255114 | EU219286 | GU320810 | EU219156 |
| G. daii | CFCC 55517 | MZ902911 | – | – | MZ936387 | MZ936403 |
| G. fagacearum | CFCC 54316T | MZ902916 | – | – | MZ936392 | MZ936408 |
| G. fructicola (=G. fragariae) | CBS 125671 | MH863616 | MH875088 | – | GU320793 | GU320776 |
| G. guangdongensis | CFCC 54443T | MZ902918 | – | – | MZ936394 | MZ936410 |
| G. hainanensis | CFCC 54376T | MZ902921 | – | – | MZ936397 | MZ936413 |
| G. idaeicola | CBS 125674T | MH863619 | MH875091 | – | GU320796 | GU320780 |
| G. macounii | CBS 121468T | MH863110 | MH874666 | – | GU320804 | – |
| G. occulta | CBS 125678 | MH863623 | MH875095 | – | GU320800 | GU320786 |
| G. paraclavulata | CBS 123202T | GU320830 | – | – | GU320815 | GU320775 |
| G. racemula | AR 3892 | EU254841 | EU255122 | EU219241 | EU221889 | EU219125 |
| G. rosae | CBS 145085 | MK047451 | MK047501 | MK047547 | – | – |
| G. rosae | GUCC 408.7 | OP581231 | OP581244 | OP688512 | OP688536 | OP688561 |
| G. rosae | GUCC 408.17 | OP581232 | OP581245 | OP688513 | OP688537 | OP688562 |
| G. rossmaniae | CFCC 54307T | MZ902923 | – | – | MZ936399 | MZ936415 |
| G. sanguisorbae | CBS 858.79 | GU320818 | KY496735 | – | GU320805 | GU320790 |
| G. silvicola | CFCC 54418T | MZ902926 | – | – | MZ936402 | MZ936418 |
| G. smithogilvyi | CBS 130190T | MH865607 | MH877031 | JQ910648 | JQ910645 | JQ910639 |
| G. tormentillae | CBS 904.79 | EU254856 | EU255133 | – | GU320795 | EU219165 |
| G. xunwuensis | CFCC 53115T | MK432667 | MK429910 | – | MK578141 | MK578067 |
| Sirococcus tsugae | CBS 119626 | EF512472 | – | – | EF512534 | EU219140 |
| Species | Strain Number | GenBank Accession Number | ||
|---|---|---|---|---|
| ITS | LSU | tef1 | ||
| Coniella africana | CBS 114133T | AY339344 | AY339293 | KX833600 |
| C. diplodiella | CBS 111858T | MH862886 | KX833335 | KX833603 |
| C. diplodiella | CBS 165.84 | KX833529 | KX833354 | KX833622 |
| C. erumpens | CBS 523.78T | KX833535 | KX833361 | KX833630 |
| C. eucalyptorum | CBS 112640T | AY339338 | AY339290 | KX833637 |
| C. fusiformis | CBS 141596T | KX833576 | KX833397 | KX833674 |
| C. granati | CBS 130974 | JN815312 | KX833398 | KX833675 |
| C. granati | CBS 252.38 | KX833581 | AY339291 | KX833681 |
| C. hibisci | CBS 109757T | KX833589 | AF408337 | KX833689 |
| C. javanica | CBS 455.68T | KX833583 | KX833403 | KX833683 |
| C. koreana | CBS 143.97T | KX833584 | AF408378 | KX833684 |
| C. koreana | CFCC 52986 | MK432612 | MK429882 | MK578112 |
| C. lanneae | CBS 141597T | KX833585 | KX833404 | KX833685 |
| C. limoniformis | CBS 111021T | KX833586 | KX833405 | KX833686 |
| C. macrospora | CBS 524.73T | KX833587 | AY339292 | KX833687 |
| C. macrospora | STE-U 3935 | AY339343 | – | AY339363 |
| C. malaysiana | CBS 141598T | KX833588 | KX833406 | KX833688 |
| C. nicotianae | CBS 875.72T | KX833590 | KX833407 | KX833690 |
| C. pseudostraminea | CBS 112624T | KX833593 | KX833412 | KX833696 |
| C. quercicola | CBS 904.69T | KX833595 | KX833414 | KX833698 |
| C. quercicola | CBS 283.76 | KX833594 | KX833413 | KX833697 |
| C. quercicola | CPC 12133 | KX833596 | – | KX833699 |
| C. quercicola | GUCC 414.2 | OP581213 | OP581239 | OP688518 |
| C. quercicola | GUCC 414.21 | OP581214 | OP581240 | OP688519 |
| C. quercicola | GUCC 412.3 | OP581215 | OP581241 | OP688520 |
| C. quercicola | GUCC 405.6 | OP581216 | OP581242 | OP688521 |
| C. quercicola | GUCC 405.16 | OP581217 | OP581243 | OP688522 |
| C. straminea | CBS 149.22 | AY339348 | AY339296 | KX833704 |
| C. tibouchinae | CBS 131594T | JQ281774 | KX833418 | JQ281778 |
| C. tibouchinae | CBS 131595 | JQ281775 | KX833419 | JQ281779 |
| C. fragariae | CBS 172.49T | AY339317 | AY339282 | KX833663 |
| C. nigra | CBS 165.60T | AY339319 | KX833408 | KX833691 |
| Datasets | Maximum Parsimony (MP) | |||||
|---|---|---|---|---|---|---|
| TL | PT | CI | RI | RC | HI | |
| Foliocryphiaceae | 4515 | 9 | 0.5132 | 0.6746 | 0.3462 | 0.4868 |
| Diaporthe | 2261 | 9 | 0.5657 | 0.8362 | 0.4730 | 0.4343 |
| Gnomoniopsis | 2423 | 3 | 0.5712 | 0.5930 | 0.3387 | 0.4288 |
| Coniella | 1292 | 8 | 0.5735 | 0.7112 | 0.4079 | 0.4265 |
| Bayesian | ||||||
| Model | ASDSF | |||||
| ITS | LSU | rpb2 | tef1 | tub2 | ||
| Foliocryphiaceae | GTR+I+G | HKY+I+G | HKY+G | 0.009523 | ||
| Diaporthe | SYM+I+G | n/a | n/a | HKY+I+G | 0.009906 | |
| Gnomoniopsis | GTR+I+G | GTR+I | GTR+G | HKY+I+G | GTR+I+G | 0.009908 |
| Coniella | SYM+I+G | GTR+I+G | n/a | GTR+I+G | n/a | 0.009408 |
| Species | Strain Number | Gene Region and Alignment Positions | ||||
|---|---|---|---|---|---|---|
| ITS (1–769 Characters) | LSU (770–1617 Characters) | rpb2 (1618–2442 Characters) | tef1 (2443–2952 Characters) | tub2 (2953–3596 Characters) | ||
| (with gap: 151 characters) | (with gap: 41 characters) | (no data) | (no data) | (no data) | ||
| Chrysofolia barringtoniae | TBRC 5647 | – | – | n/a | n/a | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.4T | 31 | 4 | n/a | n/a | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.14 | 31 | 4 | n/a | n/a | n/a |
| (with gap: 158 characters) | (with gap: 41 characters) | (no data) | (with gap: 156 characters) | (no data) | ||
| Chrysofolia colombiana | CPC 24986 | – | – | n/a | – | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.4T | 7 | 2 | n/a | 149 | n/a |
| Chrysofolia coriariae sp. nov. * | GUCC 416.14 | 7 | 2 | n/a | 149 | n/a |
| (with gap: 152 characters) | (with gap: 34 characters) | (no data) | (no data) | (no data) | ||
| Neocryphonectria carpini | CFCC 53027T | – | – | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 406.6T | 116 | 22 | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.3 | 116 | 21 | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.4 | 116 | 21 | n/a | n/a | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.8 | 116 | 21 | n/a | n/a | n/a |
| (with gap: 139 characters) | (with gap: 34 characters) | (with gap: 53 characters) | (with gap: 177 characters) | (no data) | ||
| Neocryphonectria chinensis | CFCC 53025T | – | – | – | – | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 406.6T | 118 | 21 | 75 | 130 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.3 | 118 | 20 | 76 | 131 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.4 | 118 | 20 | n/a | 129 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.8 | 118 | 20 | n/a | 132 | n/a |
| (with gap: 333 characters) | (with gap: 34 characters) | (with gap: 200 characters) | (with gap: 177 characters) | (no data) | ||
| Neocryphonectria chinensis | CFCC 53029 | – | – | – | – | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 406.6T | 32 | 21 | 66 | 130 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.3 | 32 | 20 | 67 | 131 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.4 | 32 | 20 | n/a | 129 | n/a |
| Pseudomastigosporella guizhouensis sp. nov. * | GUCC 405.8 | 32 | 20 | n/a | 132 | n/a |
| ITS (1–640 characters) | (no data) | (no data) | tef1 (641–1116 characters) | tub2 (1117–1770 characters) | ||
| (with gap: 146 characters) | (no data) | (no data) | (with gap: 163 characters) | (with gap: 140 characters) | ||
| Diaporthe juglandigena sp. nov. * | GUCC 422.16T | – | n/a | n/a | – | – |
| Diaporthe juglandigena sp. nov. * | GUCC 422.161 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe chimonanthi | HGUP191001 | 18 | n/a | n/a | n/a | 5 |
| Diaporthe chimonanthi | HGUP192087 | 23 | n/a | n/a | n/a | 2 |
| Diaporthe caryae | CFCC 52563T | 2 | n/a | n/a | 6 | 17 |
| Diaporthe caryae | PSCG520 | 4 | n/a | n/a | 6 | 30 |
| (with gap: 98 characters) | (no data) | (no data) | (with gap: 275 characters) | (with gap: 151 characters) | ||
| Diaporthe eucommiigena sp. nov. * | GUCC 420.9T | – | n/a | n/a | – | – |
| Diaporthe eucommiigena sp. nov. * | GUCC 420.19 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe passiflorae | CBS 132527T | 11 | n/a | n/a | 23 | 13 |
| Diaporthe malorum | CAA734T | 15 | n/a | n/a | 19 | 13 |
| Diaporthe malorum | CAA740 | 15 | n/a | n/a | 19 | 13 |
| Diaporthe malorum | CAA752 | 15 | n/a | n/a | 19 | 13 |
| (with gap: 128 characters) | (no data) | (no data) | (with gap: 147 characters) | (with gap: 255 characters) | ||
| Diaporthe dejiangensis sp. nov. * | GUCC 421.2T | – | n/a | n/a | – | – |
| Diaporthe dejiangensis sp. nov. * | GUCC 421.21 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe eres | CBS 138594T | 4 | n/a | n/a | 6 | 6 |
| Diaporthe eres | CAA801 | 3 | n/a | n/a | 7 | 7 |
| Diaporthe cotoneastri | CBS 439.82T | 7 | n/a | n/a | 5 | 13 |
| (with gap: 67 characters) | (no data) | (no data) | (with gap: 105 characters) | (with gap: 138 characters) | ||
| Diaporthe tongrensis sp. nov. * | GUCC 421.10T | – | n/a | n/a | – | – |
| Diaporthe tongrensis sp. nov. * | GUCC 421.101 | 0 | n/a | n/a | 0 | 0 |
| Diaporthe phragmitis | CBS 138897T | 8 | n/a | n/a | n/a | 14 |
| (with gap: 98 characters) | (no data) | (no data) | (with gap: 145 characters) | (with gap: 216 characters) | ||
| Diaporthe hyperici sp. nov. * | GUCC 414.4T | – | n/a | n/a | – | – |
| Diaporthe hyperici sp. nov. * | GUCC 414.14 | 0 | n/a | n/a | 0 | 2 |
| Diaporthe caulivora | CBS 127268T | 17 | n/a | n/a | 30 | 11 |
| Diaporthe caulivora | Dip1 | 17 | n/a | n/a | 29 | n/a |
| Diaporthe caulivora | Dpc11 | 17 | n/a | n/a | 29 | n/a |
| ITS (1–572 characters) | LSU (573–1423 characters) | rpb2 (1424–2460 characters) | tef1 (2461–2861 characters) | tub2 (2862–3354 characters) | ||
| (with gap: 34 characters) | (with gap: 11 characters) | (with gap: 279 characters) | (no data) | (no data) | ||
| Gnomoniopsis rosae | CBS 145085 | – | – | – | (no data) | (no data) |
| Gnomoniopsis rosae * | GUCC 408.7 | 0 | 0 | 0 | n/a | n/a |
| Gnomoniopsis rosae * | GUCC 408.17 | 0 | 0 | 0 | n/a | n/a |
| ITS (1–595 characters) | LSU (596–1767 characters) | (no data) | tef1 (1768–2165 characters | (no data) | ||
| (with gap: 40 characters) | (with gap: 390 characters) | n/a | (with gap: 135 characters) | n/a | ||
| Coniella quercicola | CBS 904.69T | – | – | n/a | – | n/a |
| Coniella quercicola | CBS 283.76 | 0 | 0 | n/a | 1 | n/a |
| Coniella quercicola | CPC 12133 | 1 | n/a | n/a | 5 | n/a |
| Coniella quercicola * | GUCC 414.2 | 0 | 0 | n/a | 10 | n/a |
| Coniella quercicola * | GUCC 414.21 | 0 | 0 | n/a | 10 | n/a |
| Coniella quercicola * | GUCC 412.3 | 0 | 0 | n/a | 18 | n/a |
| Coniella quercicola * | GUCC 405.6 | 0 | 0 | n/a | 7 | n/a |
| Coniella quercicola * | GUCC 405.16 | 0 | 0 | n/a | 6 | n/a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.-Y.; McKenzie, E.H.C.; Phillips, A.J.L.; Li, Y.; Wang, Y. Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China. J. Fungi 2022, 8, 1301. https://doi.org/10.3390/jof8121301
Wang S-Y, McKenzie EHC, Phillips AJL, Li Y, Wang Y. Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China. Journal of Fungi. 2022; 8(12):1301. https://doi.org/10.3390/jof8121301
Chicago/Turabian StyleWang, Si-Yao, Eric H. C. McKenzie, Alan J. L. Phillips, Yan Li, and Yong Wang. 2022. "Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China" Journal of Fungi 8, no. 12: 1301. https://doi.org/10.3390/jof8121301
APA StyleWang, S. -Y., McKenzie, E. H. C., Phillips, A. J. L., Li, Y., & Wang, Y. (2022). Taxonomy and Multigene Phylogeny of Diaporthales in Guizhou Province, China. Journal of Fungi, 8(12), 1301. https://doi.org/10.3390/jof8121301