
Fungal Endophytes: A Potential Source of Antibacterial Compounds

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Antibacterials from Various Class of Endophytic Fungi
2.1. Ascomycetes
2.1.1. Diaporthe (Asexual State: Phomopsis)
2.1.2. Xylaria
2.1.3. Chaetomium
2.1.4. Talaromyces
2.1.5. Minor Taxa of the Ascomycetes
2.2. Anamorphic Ascomycetes
2.2.1. Aspergillus
2.2.2. Penicillium
2.2.3. Fusarium
2.2.4. Trichoderma
2.2.5. Alternaria
2.2.6. Simplicillium
2.2.7. Cladosporium
2.2.8. Pestalotiopsis
2.2.9. Phoma
2.2.10. Colletotrichum
2.2.11. Minor Taxa of Anamorphic Ascomycetes
2.3. Basidiomycetes
2.4. Zygomycetes
3. Volatile Organic Compounds (VOCs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Fungus | Source | Locality | Compounds Isolated | Biological Target | Biological Activity (MIC/IC50/ID50) | Reference |
|---|---|---|---|---|---|---|---|
| Ascomycetes | |||||||
| Diaporthe | |||||||
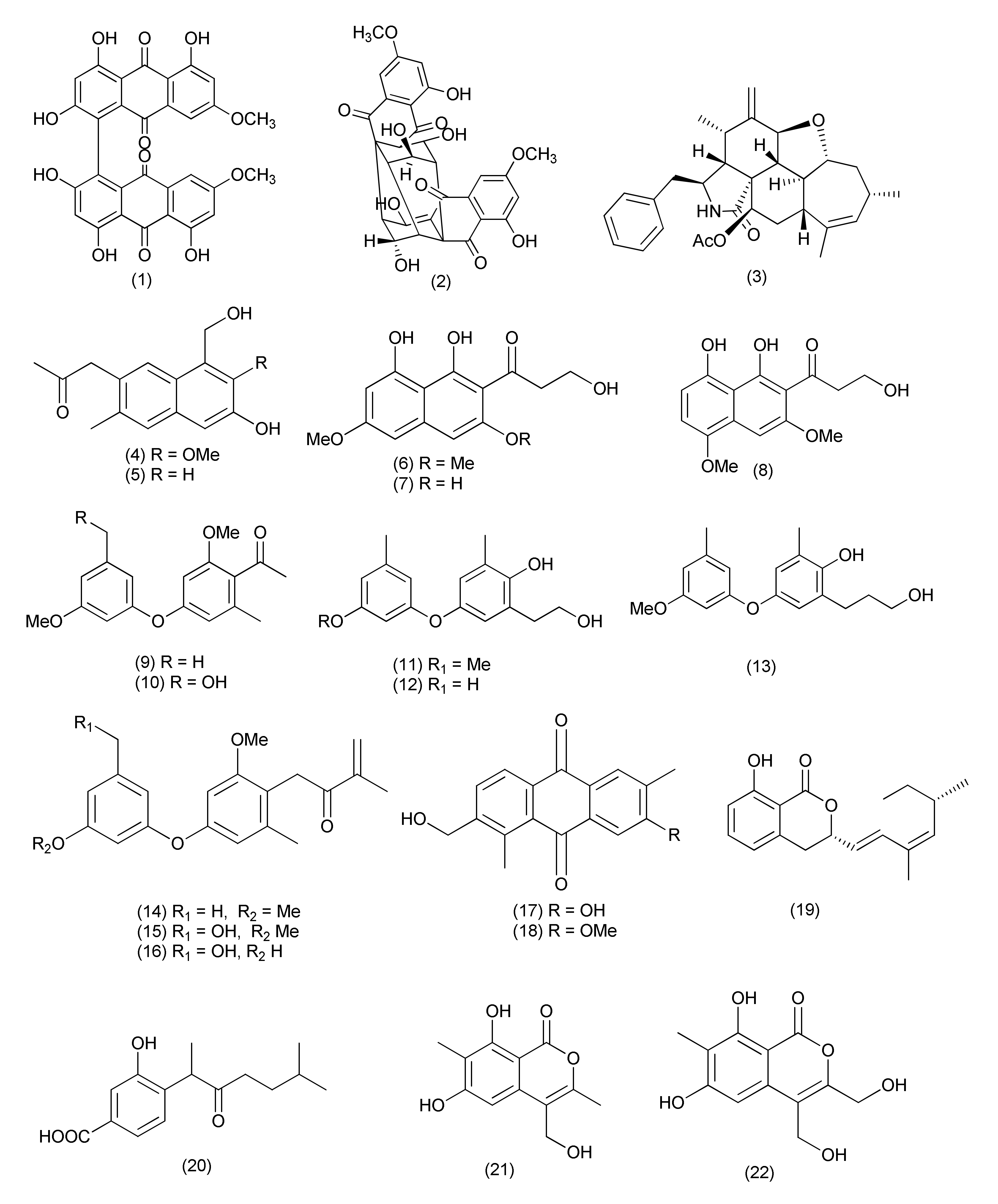
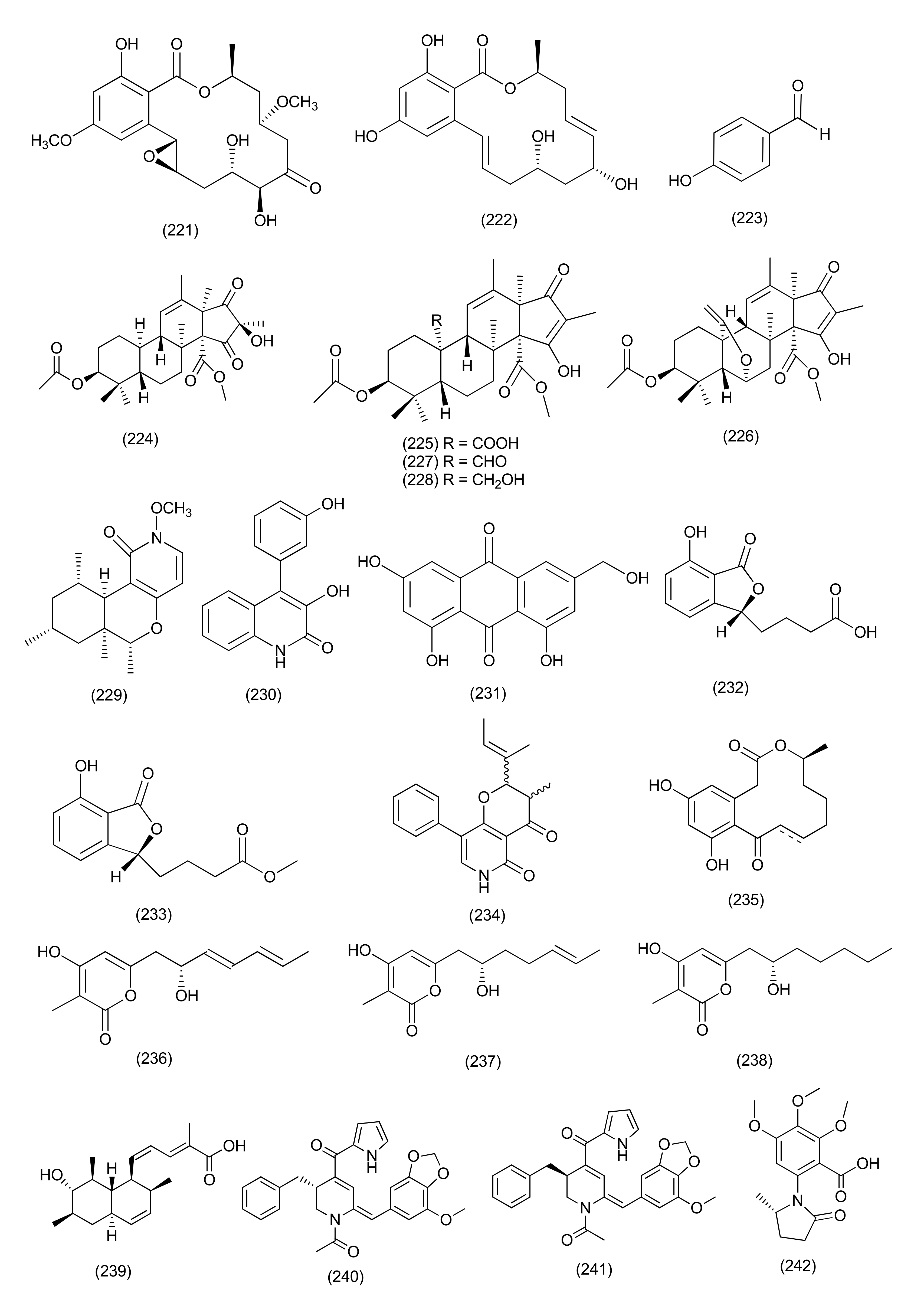
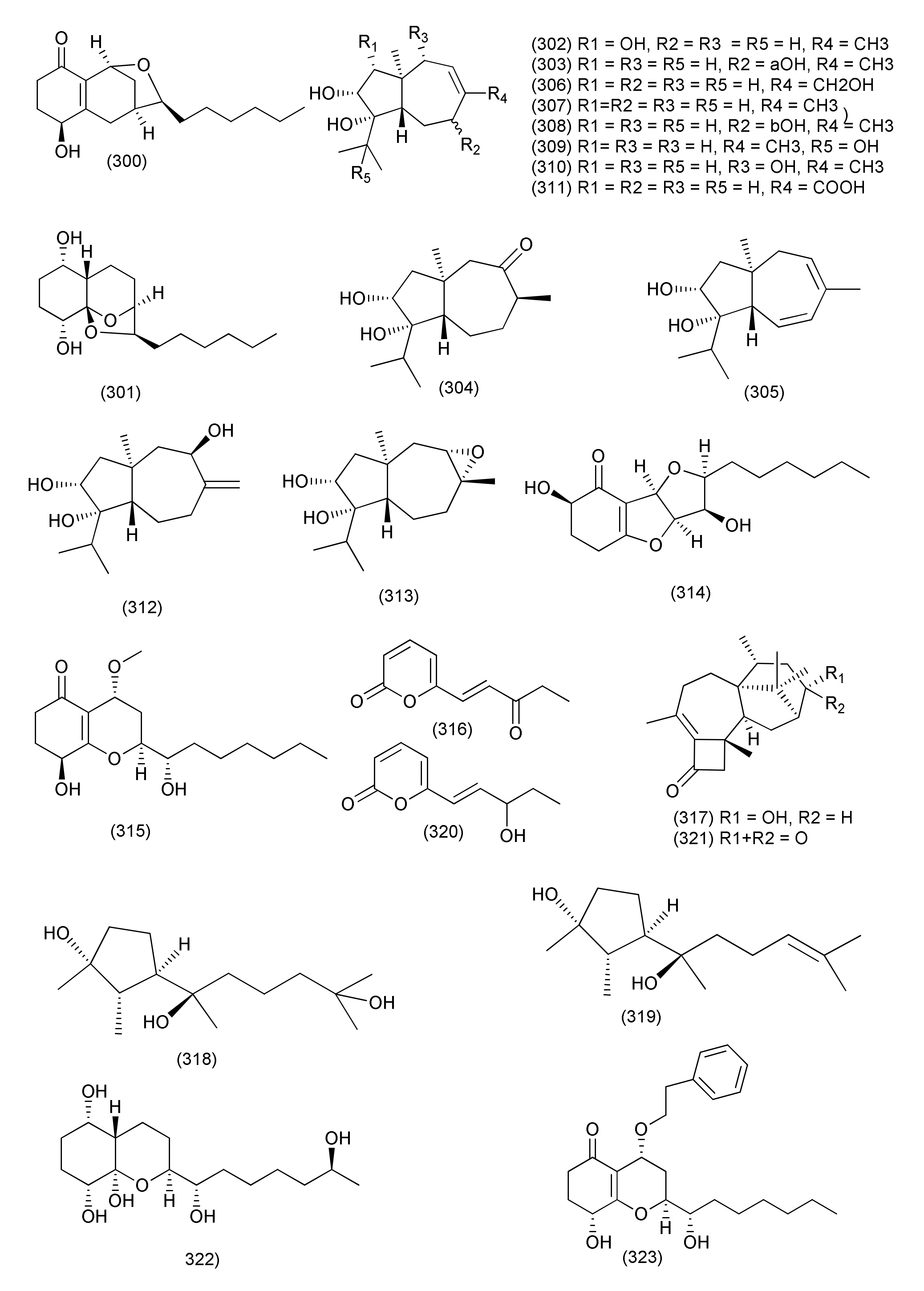
| 1 | Diaporthe sp. | Uncaria gambier | (+)-1,1′-Bislunatin (1) and (+)-2,2′- epicytoskyrin A (2) | Mycobacterium tuberculosis strains H37Rv | MICs 0.422 and 0.844 μM | [18] | |
| 2 | Diaporthe sp. GDG-118 | Sophora tonkinensis | Hechi City, China | 21-Acetoxycytochalasin J3 (3) | Bacillus anthraci and E. coli | inhibited at 12.5 μg/mL concentration | [19] |
| 3 | Phomopsis fukushii. | 1-(3-Hydroxy-1-(hydroxymethyl)-2-methoxy-6-methylnaphthalen-7-yl) propan-2-one (4) and 1-(3-hydroxy-1- (hydroxymethyl)-6-methylnaphthalen-7-yl)propan-2-one (5) | MRSA | Zone of inhibition of 10.2 and 11.3 mm (6 mm strile filterpaper disc were impregnated with 20µL (50 µg) of each compound) | [20] | ||
| 4 | Phomopsis fukushii | Paris polyphylla var. yunnanensis | Kunming, Yunnan, China | 3-Hydroxy-1-(1,8- dihydroxy- 3,6-dimethoxynaphthalen-2-yl)propan-1-one (6), 3-hydroxy-1-(1,3,8-trihydroxy-6-methoxynaphthalen-2-yl)propan-1-one (7) and 3-hydroxy-1-(1,8-dihydroxy3,5-dimethoxynaphthalen-2-yl) propan-1-one (8) | MRSA- ZR11 | MIC, 8, 4, and 4 µg/mL, | [21] |
| 5 | Phomopsis fukushii | Paris polyphylla var. yunnanensis | Kunming, Yunnan, China | 1-[2-Methoxy-4-(3-methoxy-5-methylphenoxy)-6-methylphenyl]-ethanone (9) and 1-[4-(3-(hydroxymethyl)-5-methoxyphenoxy)-2-methoxy-6-methylphenyl]-ethanone (10) | MRSA | Zone of inhibition 13.8 and 14.6 mm | [22] |
| 6 | Phomopsis fukushii | Paris polyphylla var. yunnanensis | Kunming, Yunnan, P. R. China | 4-(3-Methoxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-methylphenol (11), 4-(3-Hydroxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-methylphenol (12) and 4-(3-methoxy-5-methylphenoxy)-2-(3-hydroxypropyl) -6-methylphenol (13) | MRSA | Zone of inhibition of 20.2, 17.9 and 15.2 mm (tested at 50µg/6 mm disc) | [23] |
| 7 | Phomopsis fukushii | Paris polyphylla var. yunnanensis | Kunming, Yunnan, China. | 1-(4-(3-Methoxy-5-methylphenoxy)-2-methoxy-6-methylphenyl)-3-methylbut-3-en-2-one (14), 1-(4-(3-(hydroxymethyl)-5-methoxyphenoxy)-2-methoxy-6- methylphenyl)-3-methylbut-3-en-2-one (15), 1-(4-(3-hydroxy-5-(hydroxymethyl)phenoxy)-2-methoxy-6- methylphenyl)-3-methylbut-3-en-2-one (16) | MRSA | Zone of inhibition of 21.8, 16.8 and 15.6 mm, (50 µg/6 mm disc) | [24] |
| 8 | Phomopsis sp. | - | - | 3-Hydroxy-6-hydroxymethyl-2,5-dimethylanthraquinone (17), 6-hydroxymethyl-3-methoxy-2,5-dimethylanthraquinone (18) | MRSA | IZD 14.2 and 14.8 mm | [25] |
| 9 | Diaporthe sp. | Pteroceltis tatarinowii | Mufu Mountain of Nanjing, China. | Diaporone A (19) | B. subtilis | MIC, 66.7 μM, | [26] |
| 10 | Phomopsis prunorum (F4-3). | - | - | (−)-1 and (+)- Phomoterpenes A and B (20) phomoisocoumarins C (21), D (22) | X. citri pv. phaseoli var. fuscans | MIC, 31.2, 62.4, 31.2, and 31.2 μg/mL, | [27] |
| Pseudomonas syringae pv. Lachrymans | MIC, 31.2, 15.6, 31.2 and 15.6 μg/mL | ||||||
| 11 | Diporthe vochysiae LGMF1583 | Vochysia divergens | - | Vochysiamides A (23) | KPC (Klebsiella pneumoniae carbapenemase producing). | MIC, 1.0 μg/mL | [28] |
| Vochysiamides B (24) | KPC, MSSA, MRSA | MIC, 0.08, 1.0, and 1.0 µg/mL | |||||
| 12 | Phomopsis asparagi | Paris polyphylla var. yunnanensis | Kunming, Yunnan, China | 4-(3-Methoxy-5-methylphenoxy)-2-(2-hydroxyethyl)- 6-(hydroxymethyl)phenol (25), 4-(3-Hydroxy-5-methylphenoxy)-2-(2-hydroxyethyl)-6-(hydroxymethyl)phenol(26) | MRSA | Zone of inhibition of 10.8 and 11.4 mm | [29] |
| 13 | Phomopsis sp. | Paris polyphylla var. yunnanensis | ShiZhong, Yunnan, China | 5-Methoxy-2-methyl-7-(3-methyl-2-oxobut-3-enyl)-1-naphthaldehyde (27), 2-(hydroxymethyl)-5-methoxy-7-(3-methyl-2-oxobut-3-enyl)-1-naphthaldehyde (28) | MRSA | Zone of inhibition of 14.5 and 15.2 mm | [30] |
| 14 | Diaporthe terebinthifolii LGMF907 | Schinus terebinthifolius | Curitiba, Paraná, Brazil | Diaporthin (29) | E. coli, Micrococcus luteus, MRSA, and S. aureus | Zone of inhibition 1.73, 2.47, 9.50, and 9.0 mm tested at 100 μg/disk. | [31] |
| Orthosporin (30) | Zone of inhibition of 1.03, 1.53C, 9.0, and 9.33 mm | ||||||
| 15 | Phomopsis/Diaporthe sp. GJJM 16 | Vitex negundo | Azhiyar, Pollachi, Tamilnadu, India | (2Z)-2-(1,4-dihydro-2-hydroxy-1-((E)-2-mercapto-1 (methylimino)ethyl) pyrimidine-4-ylimino)-1-(4,5-dihydro-5-methylfuran-3-yl)-3-methylbutane-1-one (31) | S. aureus, and P. aeroginosa | MIC of 1.25 μg/mL against each organism | [32] |
| 16 | Phomopsis sp. PSU-H188 | Hevea brasiliensis | Trang Province, Thailand. | Diaporthalasin (32) | S. aureus ATCC25923, MRSA | MIC, 4 μg/mL each | [33] |
| Cytosporone B (33) | MIC, 32 and 16 μg/mL | ||||||
| Cytosporone D (34) | MIC, 64 and 32 μg/mL | ||||||
| 17 | Diaporthe terebinthifolii GG3F6 | Glycyrrhiza glabra | Jammu, J & K, India | Diapolic acid A (35), B (36) xylarolide (37) phomolide G (38) | Yersinia enterocolitica | IC50, 78.4, 73.4, 72.1 and 69.2 μM | [34] |
| 18 | Diaporthe sp. F2934 | leaves of Siparuna gesnerioides | Chagres National Park, a protected area of Panama | Phomosine A (39) | S. aureus (ATCC 25923), Streptococcus oralis (ATTC 35037), Enterococcus faecalis (ATCC 19433), Enterococcus cloacae (ATCC 13047), Bordetella bronchiseptica (CECT 440), | Zone of Inhibition 12, 9, 10, 11, 10 and 10 mm at 4 µg/mL concentration | [35] |
| Phomosine C (40) | Zone of Inhibition 9, 6, 8, 8, 8 and 9 mm at 4 µg/mL concentration | ||||||
| 19 | Phomopsis sp., | Garcinia kola nuts | bought at Mokolo local market in Yaounde (Cameroon) | 18-Methoxycytochalasin J (41), cytochalasins H (42) and J (43), alternariol (44) | Shigella flexneri | MIC, 128 μg/mL each | [36] |
| 18-Methoxycytochalasin J (41), cytochalasins H (42) | S. aureus ATCC 25923 | MIC, 128 and 256 μg/mL | |||||
| 20 | Diaporthe sp. LG23 | Mahonia fortunei | Shanghai, China | 19-nor-Lanosta-5(10),6,8,24-tetraene-1α,3β,12β,22S-tetraol (45) | S. aureus, E. coli, Bacillus subtilis, P. aeruginosa, Streptococcus pyogenes | MIC, 5.0, 5.0, 2.0, 2.0 and 0.1 µg/mL | [37] |
| 3β,5α,9α-Trihydroxy-(22E,24R)-ergosta-7,22-dien-6-one (46), and chaxine C (47) | B. subtilis | MIC, 5.0 µg/mL each | |||||
| 21 | Diaporthales sp. E6927E | Ficus sphenophyllum | Ecuadorean dry forest near the Napo River, USA | Pyrrolocin A (48) | S. aureus and E. faecalis | MICs 4 and 5 µg/mL | [38] |
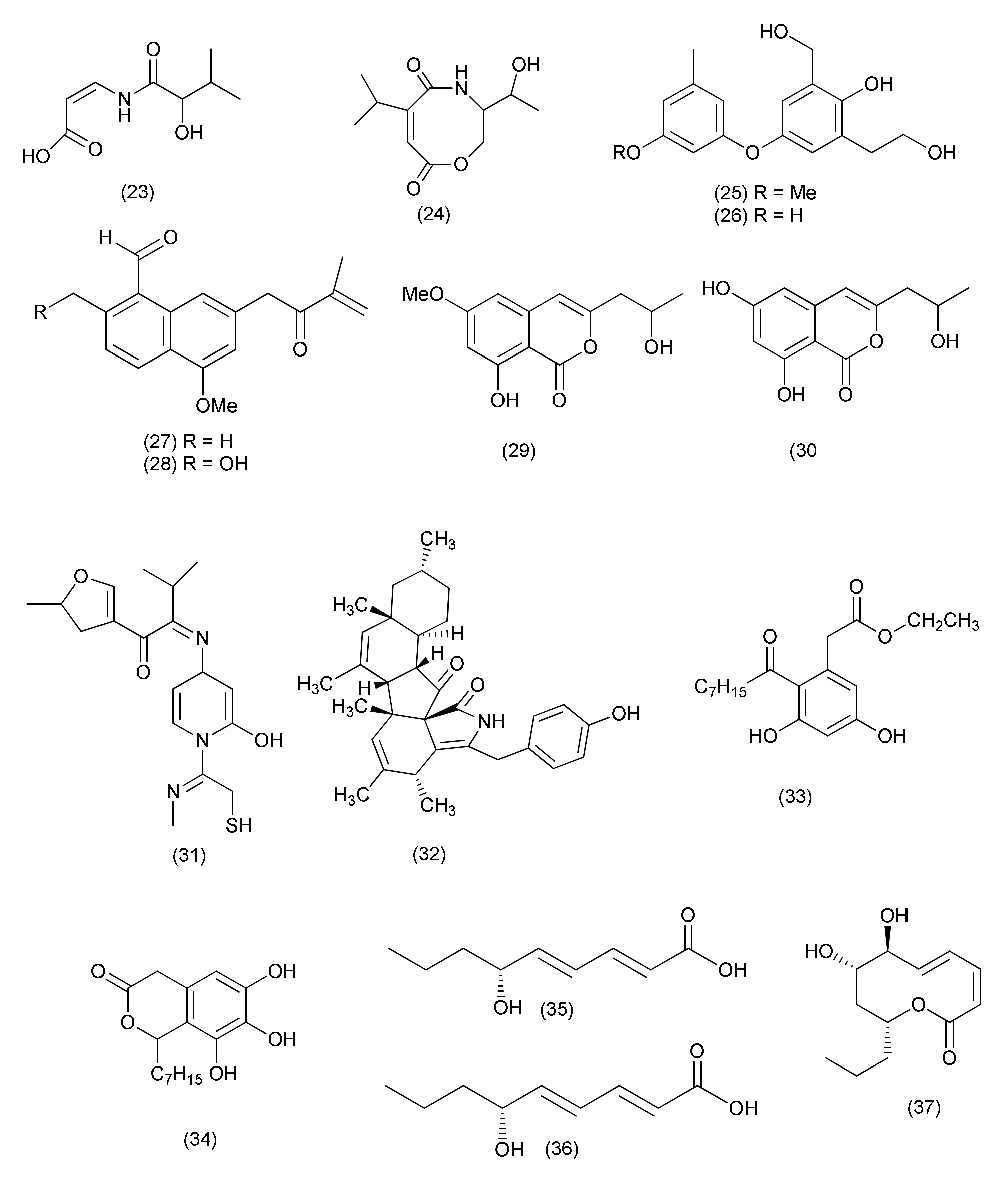
| Xylaria | |||||||
| 22 | Xylaria ellisii | Blueberry (Vaccinium angustifolium) | Ellisiiamide (49) | Escherichia coli | MIC, 100 μg/mL | [39] | |
| 23 | Xylaria sp. GDG-102 | S. tonkinensis | Hechi, Guangxi province, China | Xylareremophil (50) | Micrococcus luteus and Proteus vulgaris | MIC 25 μg/mL each | [40] |
| Mairetolides B (51) | M. luteus | MIC, 50 μg/mL | |||||
| Mairetolide G (52) | P. vulgaris M. luteus | MIC 25 and 50 μg/mL | |||||
| Xylareremophil (50), mairetolides B (51), and G (52) | Micrococcus lysodeikticus and Bacillus subtilis | MIC 100 μg/mL | |||||
| 24 | Xylaria sp. (GDG-102) | Leaves of S. tonkinensis | 6-Heptanoyl-4-methoxy-2H-pyran-2-one (53) | E. coli as well as S. aureus | MIC, 50 μg/mL | [41] | |
| 25 | Xylaria sp. GDG-102 | S. tonkinensis | Hechi, Guangxi province, China | Xylarphthalide A (54) | B. subtilis and E. coli, | MIC, 12.5 μg/mL each | [42] |
| B. megaterium, S. aureus, S. dysenteriae and S. paratyphi | MIC, 25 μg/mL each | ||||||
| (−)-5-Carboxymellein (55) | B. Subtilis | MIC, 12.5 μg/mL | |||||
| B. anthracis, B. megaterium, S. aureus, E. coli, S. dysenteriae and S. paratyphi B | MIC, 25 μg/mL | ||||||
| (−)-5-Methylmellein (56) | B. subtilis and S. aureus | MIC, 12.5 μg/mL | |||||
| B. megaterium, E. coli and S. dysenteriae | 25 μg/mL | ||||||
| 26 | Xylaria sp., | Taxus mairei. | 3,7-Dimethyl-9-(-2,2,5,5-tetramethyl-1,3-dioxolan-4-yl) nona-1,6-dien-3-ol (57) | B. subtilis ATCC 9372, B. pumilus 7061 and S. aureus ATCC 25923 | 48.1, 31.6 and 47.1% inhibition. | [43] | |
| Nalgiovensin (58) | S. aureus ATCC 25923, B. subtilis ATCC 9372, B. pumilus ATCC 7061 and E. coli ATCC 25922 | 42.1, 36.8, 47.1 and 41.2% inhibition. | |||||
| Chaetomium | |||||||
| 27 | C. globosum 7s-1, | Rhapis cochinchinensis | Xanthoquinodin B9 (59), xanthoquinodin A1 (60), xanthoquinodin A3 (61) | B. cereus | MICs of 0.87, 0.44 and 0.22 μM, | [45] | |
| Xanthoquinodin B9 (59), xanthoquinodin A1 (60), xanthoquinodin A3 (61) | S. aureus and MRSA | MIC values ranging from 0.87 to 1.75 μM | |||||
| 3-Epipolythiodioxopiperazines, chetomin (62), chaetocochin C (63) and dethio-tetra(methylthio)chetomin (64) | B. cereus ATCC 11778, S. aureus ATCC 6538, and MRSA | MIC values ranging from 0.02 pM to 10.81 μM. | |||||
| Chetomin (62) | B. cereus, S. aureus and MRSA | MICs, 0.35 μM, 10.74 and 0.02 pM | |||||
| Compounds 59–64 | E. coli ATCC 25922, P. aeruginosa ATCC 27853, and Salmonella typhimurium ATCC 13311 | MICs of 45.06 to >223.72 μM | |||||
| Epipolythiodioxopiperazines (62–64) | Mycobacterium tuberculosis | MICs, 0.55, 4.06 and 8.11 μM, | |||||
| 28 | Chaetomium sp. SYP-F7950 | Panax notoginseng | Wenshan, Yunnan, China | Chaetocochin C (63), chetomin A (65), and chetomin (62) | S. aureus, B. subtilis, Enterococcus faecium | MIC values ranging from 0.12 to 19.3 μg/mL | [46] |
| 29 | Chaetomium sp. HQ-1, | Astragalus chinensis | Tai’an, Shandong Province, China | Differanisole A (66) | L. monocytogenes S. aureus and MRSA, | MIC, 16, 128, 128 μg/mL | [47] |
| 2,6-Dichloro-4-propylphenol (67), 4,5-dimethylresorcinol (68) | L. monocytogenes | MICs of 64 and 32 μg/mL, | |||||
| 30 | Chaetomium nigricolor F5, | Mahonia fortune | Qingdao, People’s Republic of China | Chamiside A (69) | S. aureus | MIC of 25 μg/mL | [48] |
| 31 | C. globosum | Salvia miltiorrhiza | Shenyang, Liaoning province, China | Equisetin (70) | Multidrug-resistant E. faecalis, E. faecium, S. aureus, and S. epidermidis | MIC values of 3.13, 6.25, 3.13, and 6.25 μg/mL | [49] |
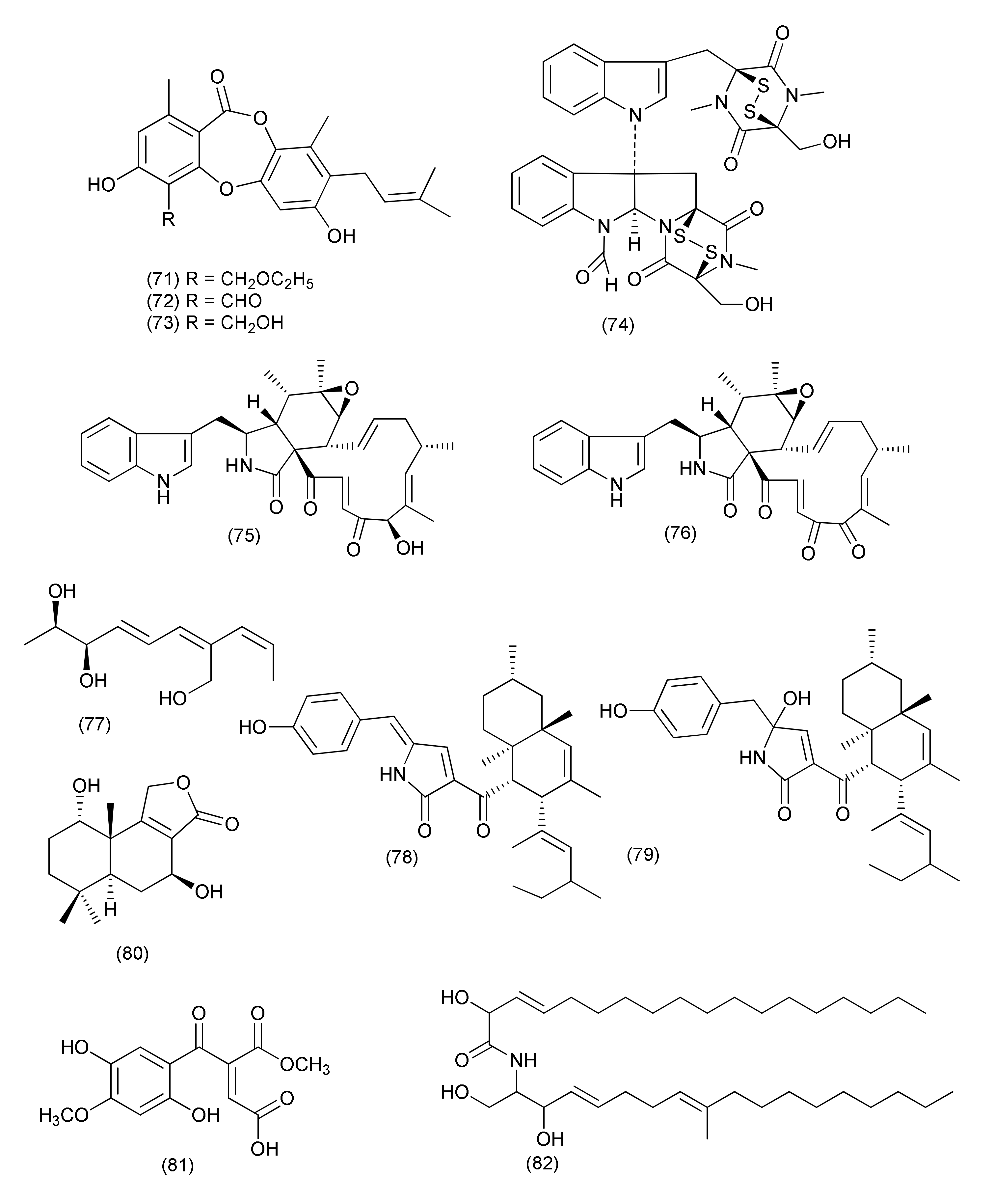
| 32 | Chaetomium sp. Eef-10, | Eucalyptus exserta | Guangdong Province, China | Mollicellins H (71) | S. aureus ATCC29213, S. aureus N50, MRSA, | IC50, 5.14, and 6.21 μg/mL | [50] |
| Mollicellin O (72) | S. aureus ATCC29213 and S. aureus N50 | IC50, 79.44 and 76.35 μg/mL | |||||
| Mollicellin I (73) | IC50, 70.14 and 63.15 μg/mL | ||||||
| 33 | Chaetomium sp. M336 | Huperzia serrata | Xichou County, Yunnan Province, China | 6-Formamidochetomin (74) | E. coli, S. aureus, S. typhimurium ATCC 6539 and E. faecalis | MIC, 0.78 μg/mL | [51] |
| 34 | Chaetomium globosum | Nymphaea nouchali | Udugampola in the Gampaha District, Sri Lanka | Chaetoglobosin A (75) | B. subtilis, S. aureus, and MRSA | MIC, 16, 32 and 32 μg/mL | [52] |
| Chaetoglobosin B (76) | >64 μg/mL | ||||||
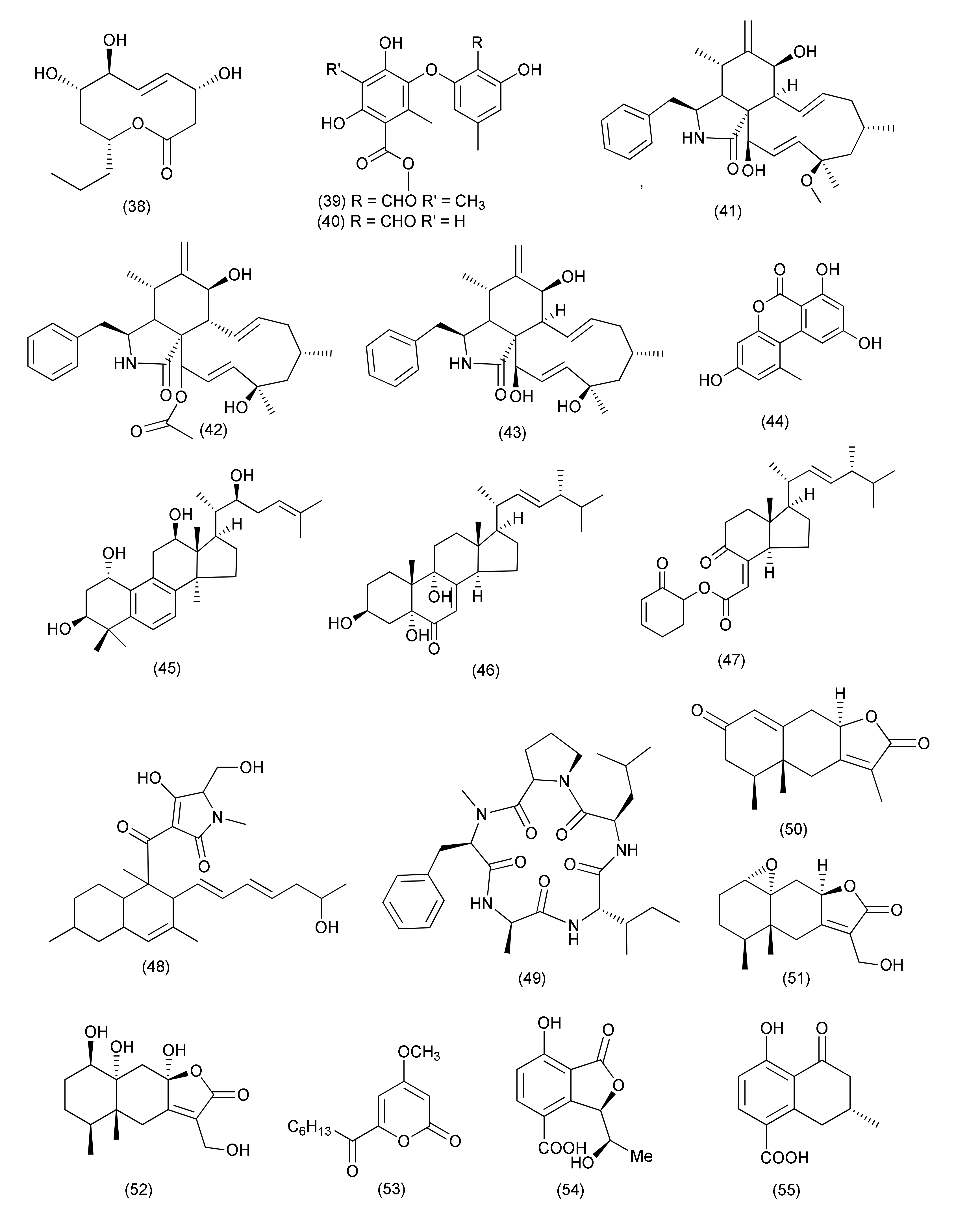
| Talaromyces | |||||||
| 35 | Talaromyces pinophilus XL-1193 | Salvia miltiorrhiza | Shenyang, Liaoning province, China | Pinophol A (77) | Bacterium paratyphosum B | MIC, 50μg/mL | [53] |
| 36 | Talaromyces purpureogenus XL-25 | Panax notoginseng | Shijiazhuang, Hebei Province, China | Talaroconvolutin A (78) | B. subtilis Micrococcus lysodeikticus, Vibrio parahaemolyticus | MIC value of 1.56 μM | [54] |
| Talaroconvolutin B (79) | MIC = 0.73 and 0.18 μM | ||||||
| 37 | Talaromyces purpureogenus | Panax notoginseng | (1S,5S,7S,10S)-dihydroxyconfertifolin (80) | E. coli | MIC, 25 μM | [55] | |
| 38 | Talaromyces funiculosus -Salicorn 58. | Talafun (81) | E. coli, S. aureus | MIC, 18 and 93 μM | [56] | ||
| N-(2′-hydroxy-3′-octadecenoyl)-9-methyl-4,8-sphingadienin (82) | Mycobacterium smegmatis, S. aureus, Micrococcus tetragenus, and E. coli | MIC, 85, 90, 24, and 68, 93 μM | |||||
| Chrodrimanin A (83) | S. aureus, M. tetragenus, Mycobacterium phlei, and E. coli | MIC, 67, 28, 47, and 26 μM | |||||
| Chrodrimanin B (84) | E.coli | MIC, 43 μM. | |||||
| 39 | Talaromyces sp. LGT-2 | Tripterygium wilfordii. | Alkaloids 85–90 | E. coli, P. aeruginosa, S. aureus, Bnfillus licheniformis, and Streptococcus pneumoniae | MICs in the range of 0.125 to 1.0 50 μg/mL | [57] | |
| 40 | Rhytidhysteron sp. BZM-9 | Leptospermum brachyandrum | Euphorbol (91) | MRSA | MIC, 62.5 ug/mL | [58] | |
| 41 | Stagonosporopsis oculihominis | Dendrobium huoshanense. | Stagonosporopsin C (92) | Staphylococcus aureus subsp. aureus ATCC29213 | MIC50, 41.3 μM | [59] | |
| 42 | Eutypella scoparia SCBG-8. | Leptospermum brachyandrum | SCBG, Chinese Academy of Sciences, China | Eutyscoparols H (93), I (94), tetrahydroauroglaucin (95), flavoglaucin (96) | Staphylococcus aureus and MRSA | MICs in the range of 1.25 to 6.25 μg/mL | [60] |
| 43 | Eutypella scoparia SCBG-8 | Leptospermum brachyandrum | SCBG, Chinese Academy of Sciences, Guangzhou 510650, China | Eutyscoparin G (97) | S. aureus and MRSA | MIC values of 6.3 μg/mL | [61] |
| 44 | Sarocladium oryzae DX-THL3, | Oryza rufipogon Griff. | Sarocladilactone A (98), sarocladilactone B (99), helvolic acid (100), helvolinic acid (101), 6- desacetoxy-helvolic acid (102), 1,2-dihydrohelvolic acid (103) | S. aureus | MIC values of 64, 4, 8, 1, 4 and 16 μg/mL | [62] | |
| Compound 101 | B. subtilis | MIC, 64 μg/mL | |||||
| Compounds 99, 101, 103 | E. coli | MIC 64 μg/mL each | |||||
| 45 | Paraphaeosphaeria sporulosa | Fragaria x ananassa | Caserta province, Southern Italy | Cyclo(L-Pro-L-Phe) (104) | Salmonella strains, S1 and S2 | MIC 71.3 and 78.6 μg/mL | [63] |
| 46 | Aplosporella javeedii | Orychophragmus violaceus | Beijing, China | Terpestacin (105), fusaproliferin (106), mutolide (108) | M. tuberculosis H37Rv | MICs of 100 μM | [64] |
| 6,7,9,10-Tetrahydromutolide (107) | S. aureus, | MICs of 100 μM | |||||
| 47 | Pleosporales sp. Sigrf05 | roots of Siraitia grosvenorii | Guangxi Province of China | Pleospyrone E (109) | B. subtilis, Agrobacterium tumefaciens, Ralstonia solanacearum, and Xanthomonas vesicatoria | MIC 100.0µM each | [65] |
| 48 | Aplosporella javeedii | Orychophragmus violaceus | Beijing, China | Aplojaveediin A (110) | Staphylococcus aureus strain ATCC 29213, S. aureus strain ATCC 700699 and Bacillus subtilis (ATCC 169) | MICs 50, 50 and 25 μM, | [66] |
| Aplojaveediin F (111) | S. aureus ATCC 29213 and ATCC 700699 | MICs of 25 and 50 μM | |||||
| 49 | Paecilomyces variotii | Lawsonia Alba | University of Karachi, Pakistan | Lawsozaheer (112) | S. aureus (NCTC 6571) | 84.26% inhibition at 150 μg/mL | [67] |
| 50 | Preussia isomera OSMAC strategy | Panax notoginseng | Wenshan, Yunnan Province, China | Setosol (113) | Multidrug-resistant E. faecium, methicinllin-resistant S. aureus and multidrug-resistant E. faecalis | MIC 25 μg/mL | [68] |
| Preussia isomera. XL-1326, | Panax notoginseng | (+)- and (−)-Preuisolactone A (114, 115) | Micrococcus luteus and B. megaterium | MIC, 10.2 and 163.4 μM | [69] | ||
| 51 | Neurospora udagawae | Quercus macranthera | Kaleybar region in northwestern Iran | Udagawanones A (116) | S. aureus | MIC, 66 μg/mL | [70] |
| 52 | Xylomelasma sp. Samif07 | Salvia miltiorrhiza Bunge | 2,6-Dimethyl-5-methoxy-7-hydroxychromone (117), 6-hydroxymethyleugenin (118), 6-methoxymethyleugenin (119), isoeugenitol (120), diaporthin (29), 8-hydroxy-6-methoxy-3-methylisocoumarin (121) | Bacillus subtilis, Staphylococcus haemolyticus, A. tumefaciens, Erwinia carotovora, and Xanthomonas vesicatoria | MIC values at the range of 25 ~ 100 μg/mL | [71] | |
| 2,6-Dimethyl-5-methoxy-7-hydroxychromone (117), diaporthin (29) | B. subtilis, E. carotovora | MIC, 50 and 100 μg/mL | |||||
| 6-Hydroxymethyleugenin (118), 6-methoxymethyleugenin (119), isoeugenitol (120), diaporthin (29) | S. haemolyticus and E. carotovora | MIC, 75 μg/mL each | |||||
| 8-Hydroxy-6-methoxy-3-methylisocoumarin (121) | B. subtilis, A. tumefaciens, and X. vesicatoria, | MICs 25, 75, and 25 μg/mL, | |||||
| 53 | Amphirosellinia nigrosporaJS-1675 | Pteris cretica | (4S,5S,6S)-5,6-epoxy-4-hydroxy-3-methoxy-5-methylcyclohex-2-en-1-one (122) | Acidovorax avenae subsp. cattlyae, Agrobacterium konjaci, A. tumefaciens, Burkholderia glumae, Clavibacter michiganensis subsp. michiganensis, Pectobacterium carotovorum subsp. carotovorum, Pectobacterium chrysanthemi, Ralstonia solanacearum, Xanthomonas arboricola pv. pruni, Xanthomonas axonopodis pv. Citri, Xanthomonas euvesicatoria, Xanthomonas oryzae pv. oryzae | MICs ranging between 31.2 and 500 µg/ml | [72] | |
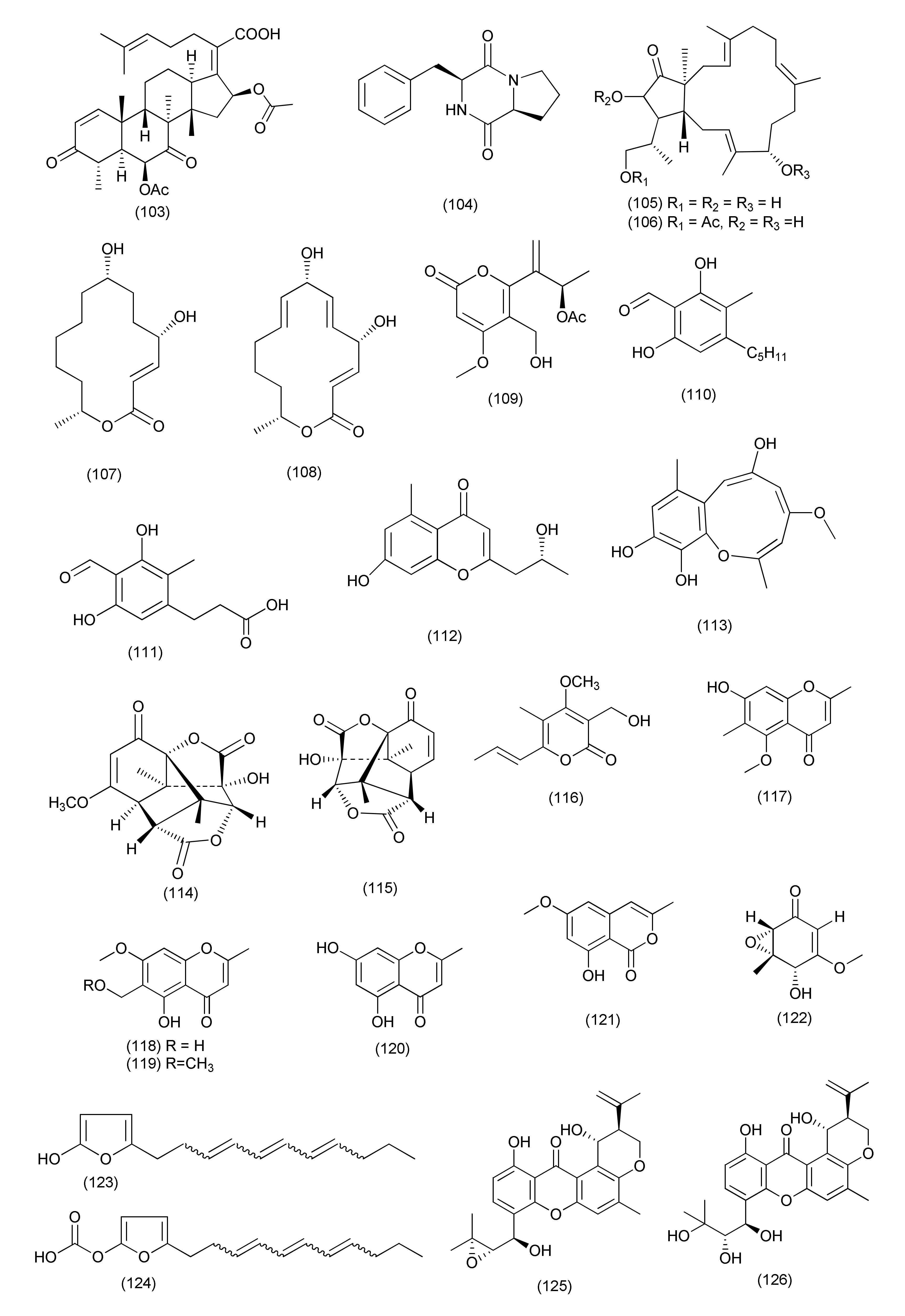
| 54 | Emericella sp. XL029 | Panax notoginseng | 5-(Undeca-3′,5′,7′-trien-1′-yl)furan-2-ol (123) and 5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-carbonate (124) | B. subtilis, B. cereus, S. aureus, B. paratyphosum B, S. typhi, P. aeruginosa, E. coli, and E. aerogenes | MIC values ranging from 6.3 to 50 μg/mL | [73] | |
| 56 | Emericella sp. XL029 | Panax notoginseng | Shijiazhuang, Hebei Province, China | 14-Hydroxytajixanthone (125), 14- hydroxytajixanthone hydrate (126), 14- hydroxy-15-chlorotajixanthone hydrate (127), 14-methoxytajixanthone-25-acetate (130), questin (132), and carnemycin B (133) | M. luteus, S. aureus, B. megaterium, B. anthracis, and B. paratyphosum B | MIC, in the range of of 12.5 and 25μg/mL | [74] |
| Epitajixanthone hydrate (128) | M. luteus, S. aureus, B. megaterium, and B. paratyphosum B | MIC 25 μg/mL | |||||
| Tajixanthone hydrate (129), 15-chlorotajixanthone hydrate (131) | S. aureus, B. megaterium, and B. paratyphosum B | MICs 25 and 12.5 μg/mL, | |||||
| 14-Hydroxytajixanthone (125) Epitajixanthone hydrate (128), carnemycin B (133) | drug resistant S. aureus | MIC 50 μg/mL | |||||
| Compounds 125–133 | P. aeruginosa, E. coli, and E. aerogenes | MIC 50 μg/mL | |||||
| 57 | Byssochlamys spectabilis | Edgeworthia chrysantha | Hangzhou Bay, Hangzhou, Zhejiang Province, China | Bysspectin C (134) | E. coli ATCC 25922 and S. aureus ATCC 25923 | MIC, 32 and 64 µg/mL | [75] |
| 58 | Poculum pseudosydowianum (TNS-F-57853), | Quercus crispula var. crispula | Yoshiwa, Hatsukaichi, Hiroshima prefecture, Japan | Sydowianumols A (135), and B (136) | MRSA | MIC90 values of 12.5 μg/mL | [76] |
| 59 | Lachnum palmae exposure to a HDAC inhibitor SAHA | Przewalskia tangutica | Linzhou Country of the Tibet Autonomous Region, China | Palmaerones A-B, E-G (137, 138, 140, 141, 142) | B. subtilis | MICs, 35, 30, 10, 50, and 55 μg/mL | [77] |
| Palmaerones A-C, E (137, 138, 139, 140) | S. aureus | MICs 65, 55, 60, and 55, μg/mL | |||||
| 60 | Nemania serpens | Vitis vinifera | Canada’s Niagara region | Nemanifuranone A (143) | E. coli | MIC 200 μg/mL | [78] |
| S. aureus, B. subtilis and M. luteus | >75% inhibition at a concentration of 100–200 μg/mL | ||||||
| Triterpenoid 144 | S. cerevisiae | (>25% inhibition) against at 200 μg/mL | |||||
| M. luteus | (>75% inhibition) of at a concentration of 100 μg/mL | ||||||
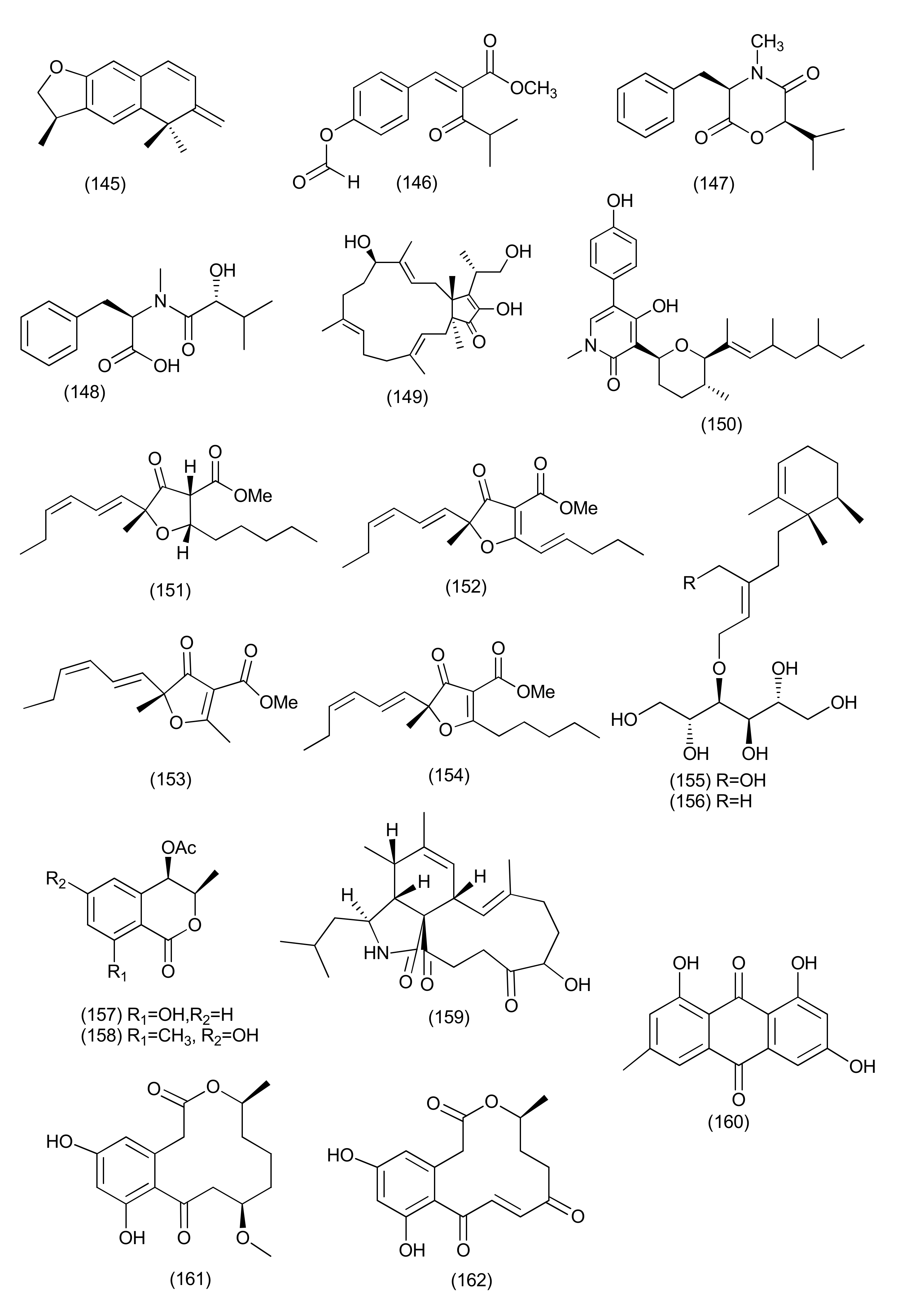
| 61 | Paraconiothyrium variabile | Cephalotaxus harringtonia | Variabilone (145) | B. subtilis | IC50 of 2.13 μg/mL after 24 h (0.36 μg/mL for kanamycin) | [79] | |
| 62 | Pyronema sp. (A2-1 & D1-2) | Taxus mairei | Shennongjia National Nature Reserve, Hubei province, China. | Methyl 2-{(E)-2-[4-(formyloxy)phenyl] ethenyl}-4-methyl-3-oxopentanoate (146), (3R,6R)-4-methyl-6-(1-methylethyl)-3-phenylmethyl-perhydro-1,4-oxazine-2,5-dione (147), (3R,6R)-N-methyl-N-(1-hydroxy-2-methylpropyl)-phenylalanine (148), siccanol (149), fusaproliferin (106), and sambutoxin (150) | Mycobacterium marinum ATCCBAA-535, | IC50 of 64, 59, 57, 84, 43 and 32 μM, (positive control rifampin IC50 of 2.1 μM) | [80] |
| 63 | Pulvinula sp. 11120 | Cupressus arizonica | Tucson, AZ, USA | Pulvinulin A (151), graminin C (152), cis-gregatin B (153), and graminin B (154) | E. coli | 12, 18, 16 and14 mm zone of inhibition at 100 μg/mL | [81] |
| 64 | Stelliosphaera formicum | Duroia hirsuta | Yasuni’ National Park off the Napo River in Ecuador | Stelliosphaerols A (155) and B (156) | S. aureus | MIC values of 250 μg/mL | [82] |
| 65 | Unidentified Ascomycete | Melilotus dentatus | cis-4-Acetoxyoxymellein (157) | E. coli and B. megaterium | Zone of inhibition of 10 and 10 mm (Partial inhibition) at a concentration of 0.05 mg | [83] | |
| 8-Deoxy-6-hydroxy-cis-4-acetoxyoxymellein (158) | E. coli and B. megaterium | Zone of inhibition of 9 and 9 mm (Partial inhibition) at a concentration of 0.05 mg | |||||
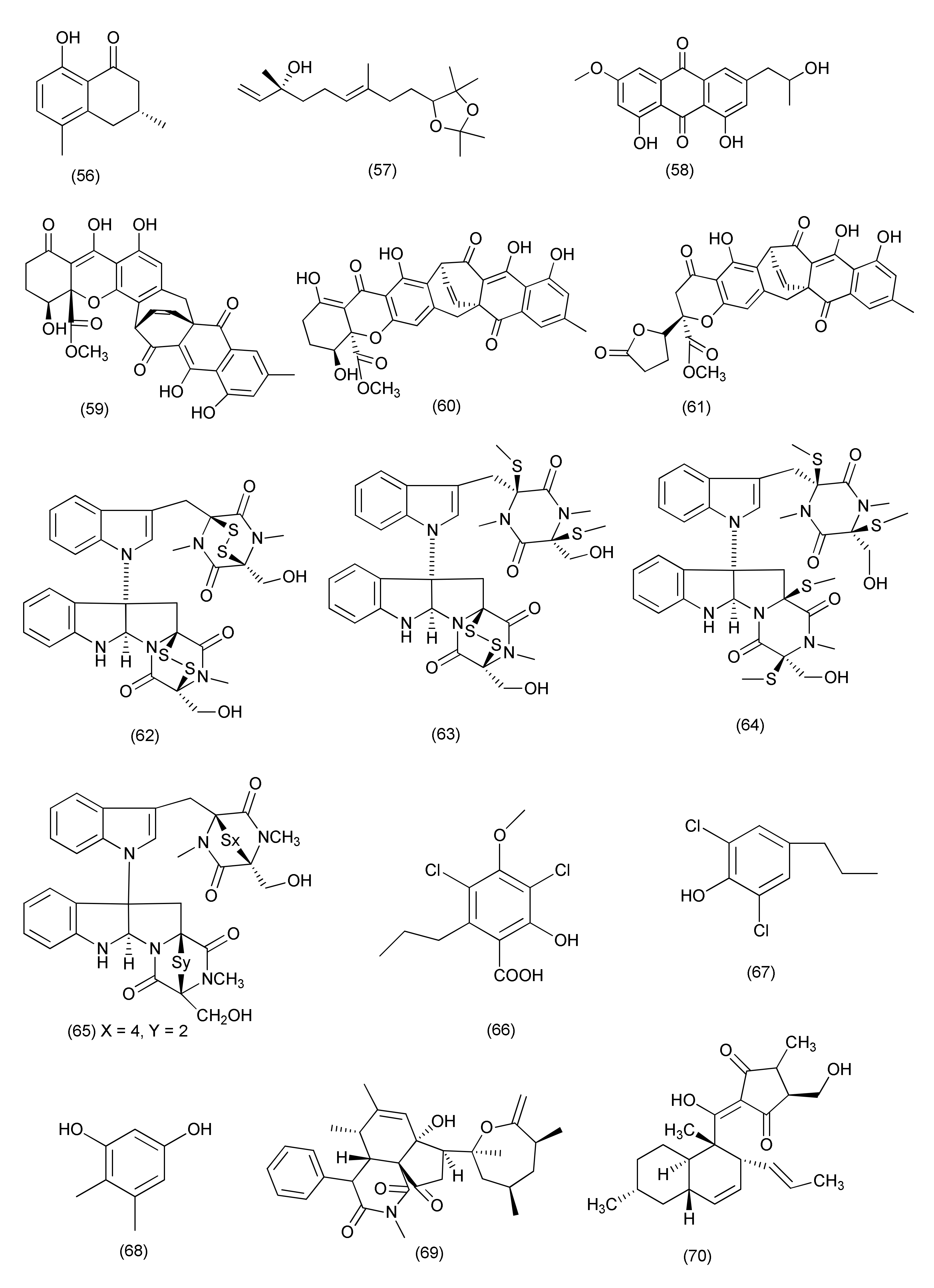
| Anamorphic Ascomycetes | |||||||
| Aspergillus | |||||||
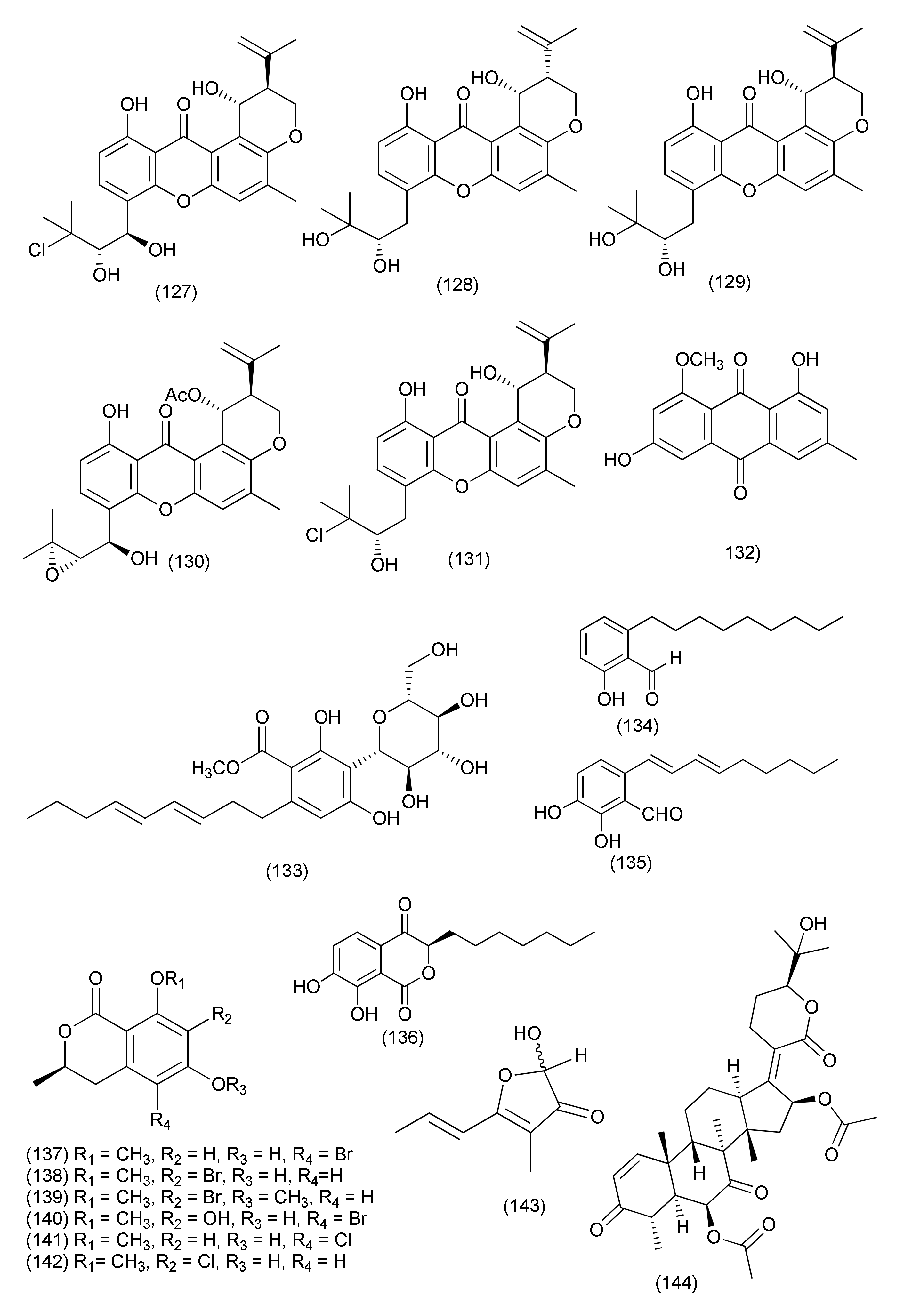
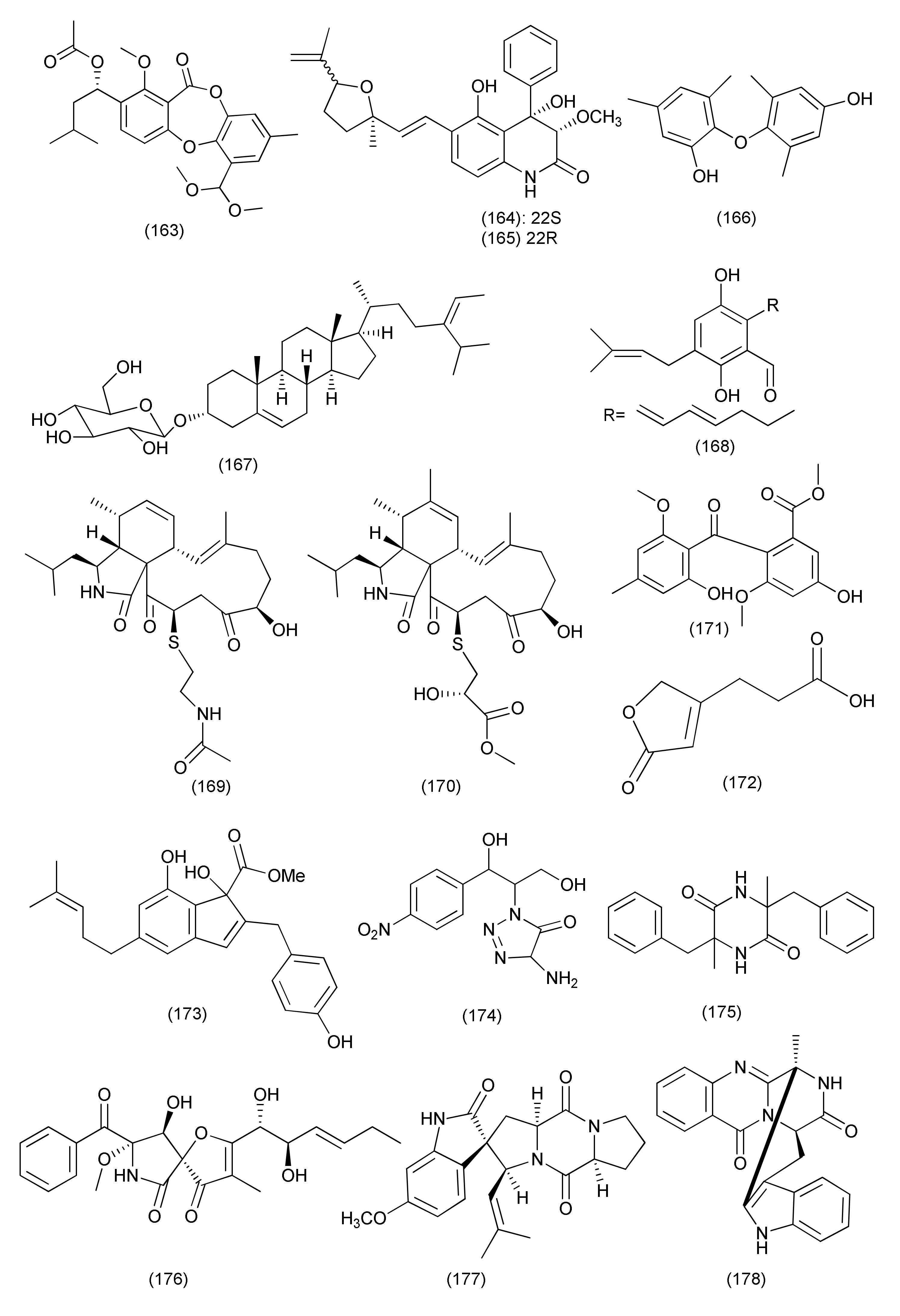
| 66 | Aspergillus sp. FT1307 | Heliotropium sp. | Aspochalasin P (159), alatinone (160), β-11-methoxy curvularine (161), 12-keto-10,11-dehydrocurvularine (162) | S. aureus ATCC12600, B. subtilis ATCC6633 and MRSA ATCC43300 | MIC in the range of 40 to 80 μg/mL | [84] | |
| 67 | Aspergillus cristatus | Pinellia ternata | Aspergillone A (163) | B. subtilis and S. aureus | MIC50, 8.5 and 32.2 μg/mL | [85] | |
| 68 | Aspergillus versicolor strain Eich.5.2.2 | Eichhornia crassipes | El-Kanater El-Khayriah in Egypt | 22S-Aniduquinolone A (164), 22R-aniduquinolone A (165) | S. aureus (ATCC700699) | MIC, 0.4 μg/mL | [86] |
| 69 | Aspergillus versicolor | roots of Pulicaria crispa | Saudi Arabia | Aspergillether B (166) | S. aureus, B. cereus, and E. coli | MICs, 4.3, 3.7, and 3.9 μg/mL | [87] |
| 70 | Aspergillus ochraceus SX-C7 eus SX-C7 | Setaginella stauntoniana | 3-O-β-D-Glucopyranosyl stigmasta-5(6),24(28)-diene (167) | Bacillus subtilis | MIC, 2 μg/mL | [88] | |
| 71 | Aspergillus amstelodami (MK215708) | Ammi majus | Egypt | Dihydroauroglaucin (168) | E. coli, Streptococcus mutans, S. aureus | MIC, 1.95, 1.95 and 3.9 μg/mL | [89] |
| S. aureus, E. coli, Streptococcus mutans, P. aeruginosa | Minimum biofilm inhibitory concentration (MBIC) = 7.81, 7.81, 15.63 and 31.25 μg/mL | ||||||
| 72 | Aspergillus micronesiensis | Phyllanthus glaucus | LuShan Mountain, Jiangxi Province, China | Cyschalasins A (169) and B (170) | MRSA | MIC50, 17.5 and 10.6 μg/mL: MIC90, 28.4 and 14.7 μg/mL | [90] |
| 73 | A. niger | Acanthus montanus | Kala Mountain neighborhood of Yaoundé, Africa | Methylsulochrin (171) | S. aureus, Enterobacter cloacae and Enterobacter aerogenes | MIC, 15.6, 7.8 and 7.8 μg/mL | [91] |
| 74 | Aspergillus tubingensis | stem of Decaisnea insignis | Qinling Mountain, Shaanxi Province, China | 3-(5-Oxo-2,5-dihydrofuran-3-yl) propanoic acid (172) | Streptococcus lactis | MIC value of 32 μg/mL | [92] |
| 75 | Aspergillus flavipes Y-62 | Suaeda glauca | Zhoushan coast, Zhejiang province, East China | Methyl 2-(4-hydroxybenzyl)-1,7-dihydroxy-6-(3-methylbut-2-enyl)-1H-indene-1-carboxylate (173) | MRSA | MIC, 128 μg/mL | [93] |
| K. pneumoniae and P. aeruginosa | MIC, of 32 μg/mL each | ||||||
| 76 | Aspergillus sp. | Rhizome of Zingiber cassumunar | 4-Amino-1-(1,3-dihydroxy-1-(4-nitrophenyl)propan-2-yl)-1H-1,2,3-triazole-5(4H)one (174) | Xanthomonas oryzae, Bacillus subtilis and E. coli | Zone of inhibition 37, 30 and 27 mm | [5] | |
| 3,6-Dibenzyl-3,6-dimethylpiperazine-2,5-dione (175) | E. coli and X. oryzae | Zone of inhibition 21 and 16 mm. | |||||
| 77 | Aspergillus fumigatus | Edgeworthia chrysantha | Hangzhou Bay (Hangzhou, China) | Pseurotin A (176), spirotryprostatin A (177) | S. aureus | MIC of 0.39 µg/mL each | [94] |
| Spirotryprostatin A (177) | E. coli | MIC, 0.39 µg/mL | |||||
| 78 | Aspergillus sp., | Astragalus membranaceus | Fumiquinazoline J (178), fumiquinazoline C (180), fumiquinazoline H (181), fumiquinazoline D (182) | B. subtilis, S. aureus, E. coli and P. aeruginosa | MICs in the range of 0.5–8 μg/mL | [95] | |
| Fumiquinazoline I (179), fumiquinazoline B (183) | MICs in the range of 4–16 μg/mL | ||||||
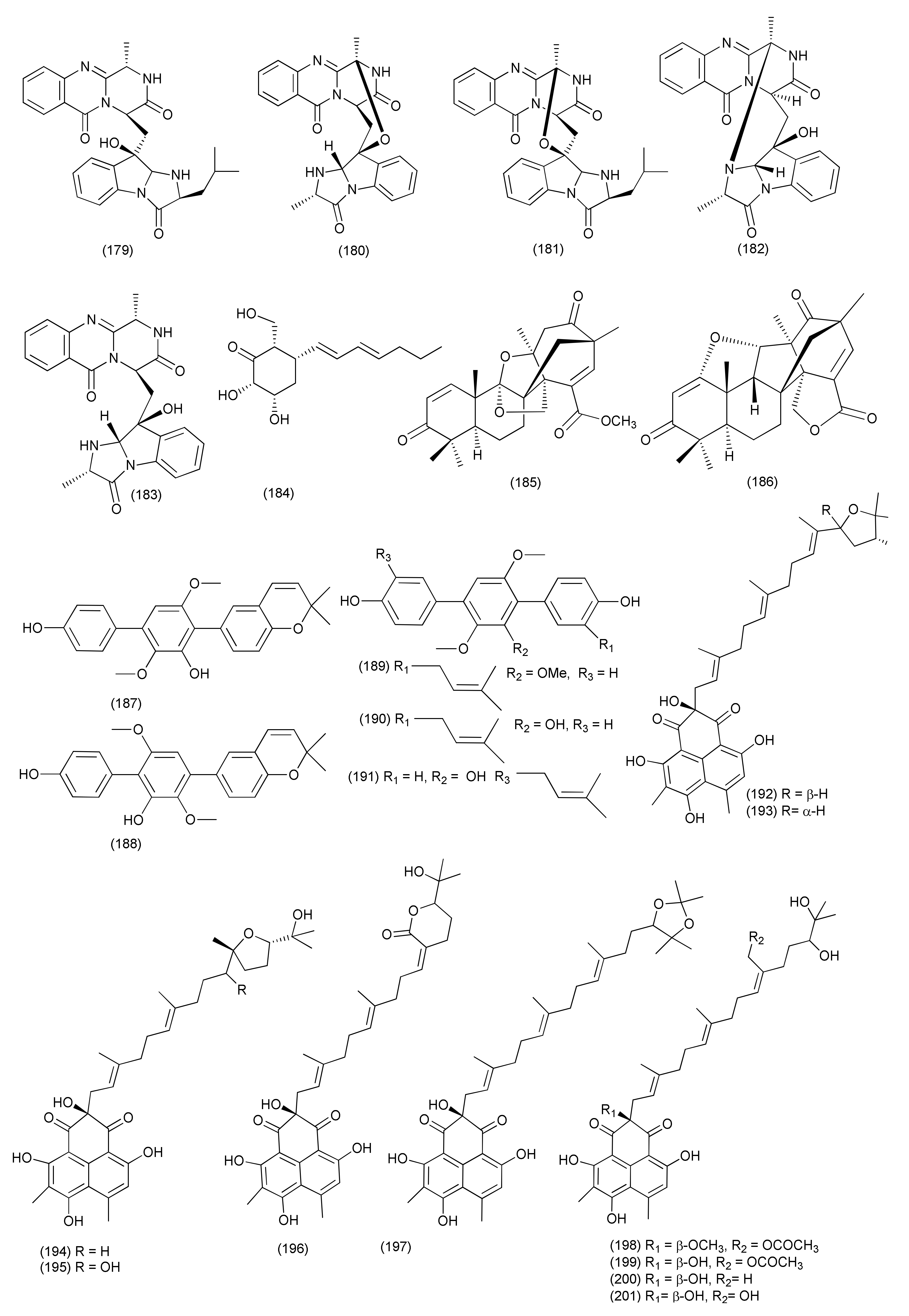
| 79 | Aspergillus fumigatiaffnis | Tribulus terestris | (−)-Palitantin (184) | E. faecalis UW 2689 and Streptococcus pneumoniae | MIC, 64μg/mL | [96] | |
| 80 | Aspergillus sp. TJ23 | Hypericum perforatum (St John’ Wort) | Shennongjia areas of Hubei Province, China | Aspermerodione (185) | MRSA | MIC, 32 μg/mL/potential inhibitor of PBP2a | [97] |
| Andiconin C (186) | marginal antimicrobial activity (>100μg/mL) | ||||||
| 81 | Aspergillus sp. YXf3 | Ginkgo biloba | Prenylterphenyllin D (187), prenylterphenyllin E (188), 2′-O-Methylprenylterphenyllin (189), prenylterphenyllin (190) | X. oryzae pv. oryzicola Swings and E. amylovora | MIC, 20 μg/mL each | [98] | |
| Prenylterphenyllin B (191) | E. amylovora | MIC, 10 μg/mL | |||||
| 82 | Aspergillus sp. | Pinellia ternata | Nanjing, Jiangsu Province, China | Aspergillussanone D (192) | P. aeruginosa, and S. aureus | MIC50, 38.47 and 29.91 μg/mL | [99] |
| Aspergillussanone E (193) | E. coli | MIC50, 7.83 μg/mL | |||||
| Aspergillussanone F (194) | P. aeruginosa, and S. aureus | MIC50, 26.56, 3.93 and 16.48 μg/mL | |||||
| Aspergillussanone G (195) | P. aeruginosa, and S. aureus, | MIC50, 24.46 and 34.66 μg/mL | |||||
| Aspergillussanone H (196) | P. aeruginosa, and E. coli, | MIC50, 8.59 and 5.87 μg/mL | |||||
| Aspergillussanone I (197) | P. aeruginosa, | MIC50, 12.0 μg/mL | |||||
| Aspergillussanone J (198) | P. aeruginosa, E. coli and S. aureus | MIC50, 28.50, 5.34 and 29.87 μg/mL | |||||
| Aspergillussanone K (199) | P. aeruginosa, and S. aureus, | MIC50, 6.55 and 21.02 μg/mL | |||||
| Aspergillussanone L (200) | P. aeruginosa, S. aureus, and B. subtilis | MIC50, 1.87, 2.77, and 4.80 μg/mL, | |||||
| Compound 201 | P. aeruginosa, and E. coli, | MIC50, 19.07 and 1.88 μg/mL | |||||
| 83 | Aspergillus terreus JAS-2 | Achyranthus aspera | Varanasi, India | Terrein (202) | E. faecalis | IC50, 20 μg/mL | [100] |
| S. aureus and Aeromonas hydrophila | 20 μg/mL | ||||||
| 84 | Aspergillus terreus | roots of Carthamus lanatus | Al-Azhar University campus in Cairo, Egypt | (22E,24R)-Stigmasta-5,7,22-trien-3-β-ol (203) | MRSA | IC50, 2.29 µM | [101] |
| 85 | Aspergillus flavus | Cephalotaxus fortunei | Taibai Mountains, Shaanxi Province, China | 5-Hydroxymethylfuran-3-carboxylic acid (204), 5-acetoxymethylfuran-3-carboxylic acid (205) | S. aureus | MIC, 31.3 and 15.6 μg/mL | [102] |
| 86 | Aspergillus allahabadii BCC45335 | root of Cinnamomum subavenium | Khao Yai National Park, Nakhon Ratchasima Province, Thailand | Allahabadolactone B (206), (22E)-5α,8α-epidioxyergosta-6,22-dien-3β-ol (207) | B. cereus | IC50, 12.50 and 3.13 µg/mL. | [103] |
| 87 | Aspergillus tubingensis | Lycium ruthenicum | 6-Isovaleryl-4-methoxypyran-2-one (208), asperpyrone A (210), campyrone A (211) | E. coli, Pseudomonas aeruginosa, Streptococcus lactis and S. aureus | MIC values ranging from 62.5 to 500 μg/mL | [104] | |
| Rubrofusarin B (209) | E. coli | MIC, 1.95 μg/mL | |||||
| 88 | Aspergillus tamarii FR02 | roots of Ficus carica | Qinling Mountain in China’s Shaanxi province | Malformin E (212) | B. subtilis, S. aureus, P. aeruginosa, and E. coli | MIC, 0.91, 0.45, 1.82, and 0.91 μM | [105] |
| 89 | Aspergillus terreus | Roots of Carthamus lanatus | Al-Azhar University campus, Egypt | (22E,24R)-Stigmasta-5,7,22-trien-3-β-ol (203) | MRSA | IC50, 0.96μg/mL | [106] |
| Aspernolide F (213) | IC50 6.39μg/mL | ||||||
| 90 | Aspergillus sp. (SbD5) | Leaves of Andrographis paniculata | Indralaya, Ogan Ilir, South Sumatra. | 1-(3,8-Dihydroxy-4,6,6-trimethyl-6H-benzochromen-2-yloxy)propane-2-one (214), 5-hydroxy-4-(hydroxymethyl)-2H-pyran-2-one (215), (5-hydroxy-2-oxo-2H-pyran-4-yl)methyl acetate (216) | S. aureus, E. coli, S. dysenteriae and Salmonella typhi | Zone of inhibition diameters ranging from 8.1 to 12.1 mm at a concentration 500 μg/mL. | [107] |
| 91 | Aspergillus sp. IFB-YXS | Ginkgo biloba | Xanthoascin (217) | X. oryzae pv. oryzicola, Swings, E.amylovora, P. syringae pv. Lachrymans and C. michiganense subsp. sepedonicus | MICs, 20, 10, 5.0 and 0.31 µg/mL | [108] | |
| Prenylterphenyllin B (218) | X. oryzae pv.oryzicola Swings, E.amylovora, P. syringae pv. Lachrymans, | MICs of 20 µg/mL each | |||||
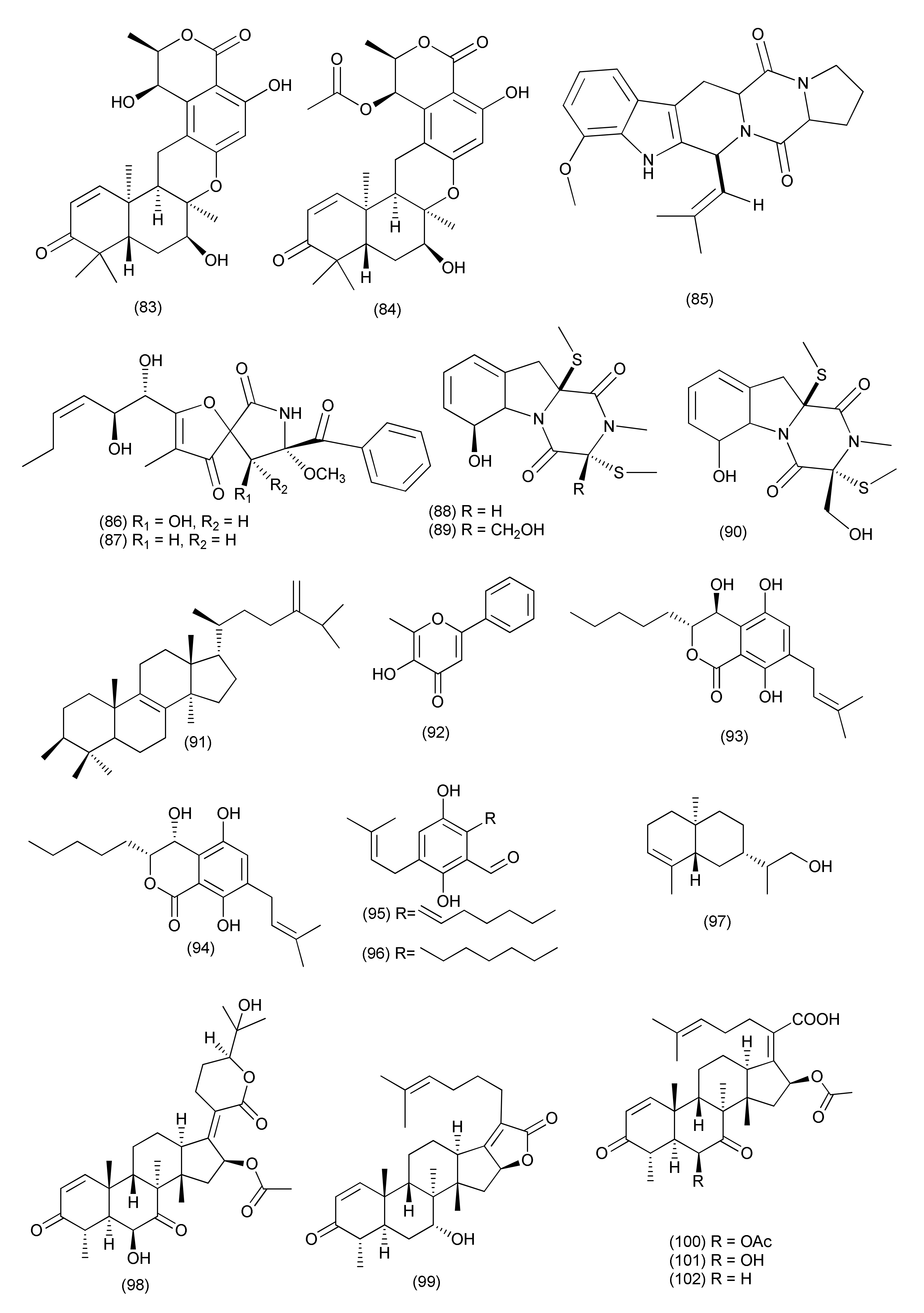
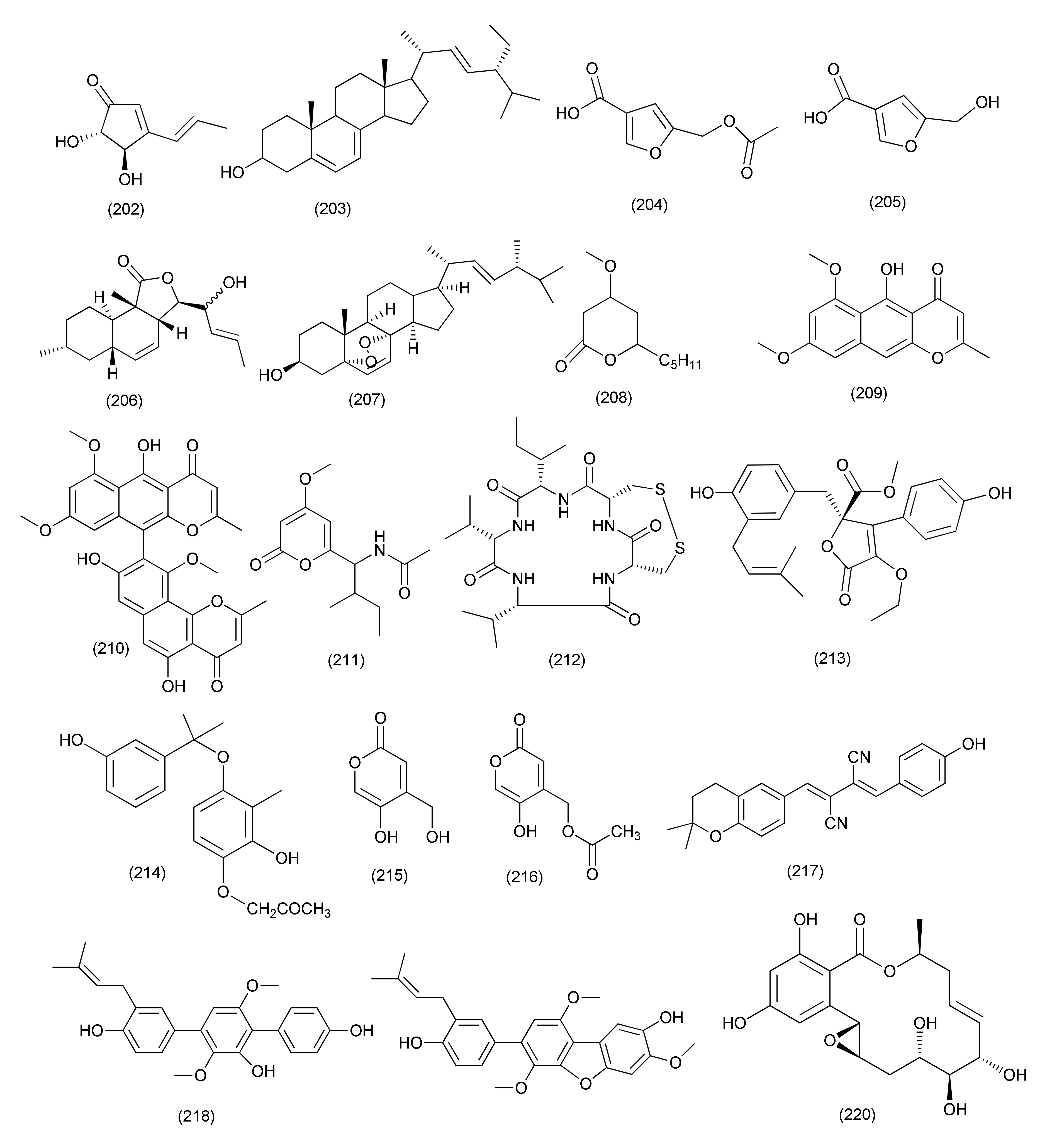
| Prenylcandidusin (219) | X. oryzae pv.oryzae Swings X. oryzae pv. oryzicola Swings | MIC values of 10 and 20 µg/mL | |||||
| Penicillium | |||||||
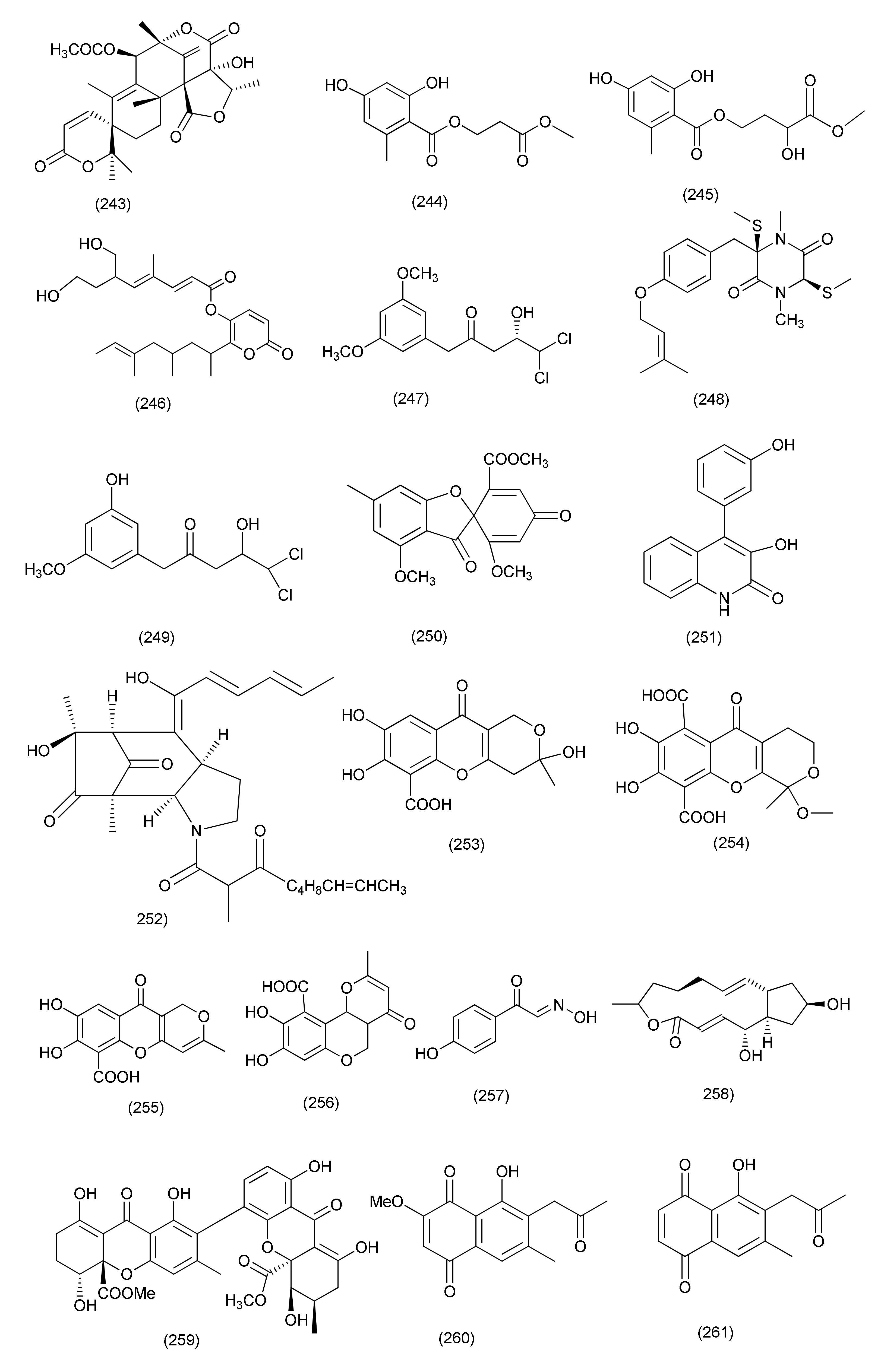
| 92 | Penicillium ochrochloron SWUKD4.1850 | Kadsura angustifolia | 4-O-Desmethylaigialomycin B (220), penochrochlactones C (221) and D (222) | Staphylococcus aureus, Bacillus subtilis, Escherichia coli, and Pseudomonas aeruginosa | MIC values between 9.7 and 32.0 μg/mL | [109] | |
| 93 | Penicillium brefeldianum | Syzygium zeylanicum | p-Hydroxybenzaldehyde (223), | S. typhi, E. coli, and B. subtilis | MIC values of 64 g/mL | [110] | |
| 94 | Penicillium vulpinum GDGJ-91 | Sophorae tonkinensis | Baise, Guangxi Province, China | 10-Demethylated andrastone A (224), andrastin A (227) | Bacillus megaterium | MIC value of 6.25 μg/mL | [111] |
| Citreohybridone E (225), citreohybridonol (226), citreohybridone B (228) | B. megaterium | MIC values of 25, 12.5 and 25 μg/mL | |||||
| Citreohybridonol (226) | B. paratyphosus B, E. coli and S. aureus | MIC, 6.25, 25 and 25 μg/mL | |||||
| 10-Demethylated andrastone A (224), citreohybridone E (225), andrastin A (227), andrastin B (228) | B. paratyphosus B | MIC, 12.5 or 25 μg/mL. | |||||
| 95 | Penicillium nothofagi P-6, | Abies beshanzuensis | Baishanzu Mountain in Lishui, Zhejiang Province of China | Chromenopyridin A (229), viridicatol (230) | S. aureus ATCC29213 | MIC, 62.5 and 15.6 μg/mL | [112] |
| 96 | Penicillium restrictum (strain G85) | Silybum marianum | Horizon Herbs, LLC (Williams, OR, USA). | ω-Hydroxyemodin (231) | Clinical isolates of MRSA | Quorum-sensing inhibition in both in vitro and in vivo models | [113] |
| 97 | Penicillium vulpinum | S. tonkinensis | Baise, Guangxi Province, China | (−)-3-Carboxypropyl-7-hydroxyphthalimide (232) | Shigella dysenteriae and Enterobacter areogenes | MIC, 12.5 μg/mL each | [114] |
| B. subtilis | MIC, 25 μg/mL | ||||||
| B. megaterium and Micrococcus lysodeikticus | MIC, 50 μg/mL | ||||||
| (−)-3-Carboxypropyl-7-hydroxyphthalide methyl ester (233) | E. areogenes | MIC, 12.5 μg/mL | |||||
| B. subtilis, B. megaterium and M. lysodeikticus | MIC, 100 μg/mL. | ||||||
| 98 | Penicillium sumatrense GZWMJZ-313 | Leaf of Garcinia multiflora | Libo, Guizhou Province of China | Citridone E (234), (–)-dehydrocurvularin (235) | S. aureus, P. aeruginosa, Clostridium perfringens, and E. coli | MIC values ranging from 32 to 64 μg/mL | [115] |
| 99 | Penicillium ochrochloronthe | Roots of Taxus media | Qingfeng Mountain, Chongqing, China | 3,4,6-Trisubstituted α-pyrone derivatives, namely 6-(2′R-hydroxy-3′E,5′E-diene-1′-heptyl)-4-hydroxy-3-methyl-2H-pyran-2-one (236), 6-(2′S-hydroxy-5′E-ene-1′-heptyl)-4-hydroxy-3-methyl-2H-pyran2-one (237), 6-(2′S-hydroxy-1′-heptyl)-4 -hydroxy-3-methyl-2H-pyran-2-one (238), trichodermic acid (239) | B. subtilis, Micrococcus luteus, S. aureus, B. megaterium, Salmonella enterica, Proteusbacillm vulgaris, Salmonella typhi, P. aeruginosa, E. coli and Enterobacter aerogenes | MIC values ranging from 25 to 50 μg/mL | [116] |
| 100 | Penicillium janthinellum SYPF 7899 | Panax notoginseng | Wenshan region, Yunnan province, China | Brasiliamide J-a (240), brasiliamide J-b (241) | B. subtilis and S. aureus | MIC, 15 and 18 μg/mL, | [117] |
| Peniciolidone (242), austin (243) | B. subtilis | MIC, 35 and 50 μg/mL | |||||
| S. aureus | MIC 39, and 60 μg/mL | ||||||
| 101 | Penicillium cataractum SYPF 7131 | Ginkgo biloba | Penicimenolidyu A (244), penicimenolidyu B (245) and rasfonin (246) | S. aureus | MIC 65, 59 and 10 μg/mL | [118] | |
| 102 | Penicillium sp., | Tubers of Pinellia ternata | suburb of Nanjing, Jiangsu, China. | 3′-Methoxycitreovirone (247), citreovirone (249) | E. coli and S. aureus | MIC = 62.6 and 76.6 μg/mL | [119] |
| Helvolic acid (100) | S. aureus, P. aeruginosa, B. subtilis and E. coli | MIC = 5.8, 4.6, 42.2 and 75.0 μg/mL | |||||
| cis-bis-(Methylthio)-silvatin (248), trypacidin A (250) | S. aureus | MIC values of 43.4 and 76.0 μg/mL | |||||
| Trypacidin A (250) | B. subtilis | MIC = 54.1 μg/mL | |||||
| 103 | Penicillium sp. R22 | Nerium indicum | Qinling Mountain, Shaanxi Province, China | Viridicatol (251) | S. aureus | MIC value of 15.6 μg/mL | [120] |
| 104 | Penicillium sp. (NO. 24) | Tapiscia sinensis | Shennongjia National Forest Park China | Penicitroamide (252) | Erwinia carotovora subsp. Carotovora | MIC50 at 45 μg/mL | [121] |
| 105 | Penicillium sp. CAM64 | Leaves of Garcinia nobilis | Mount Etinde, Southwest region Cameroon | Penialidin A (253) | Vibrio cholerae SG24 (1), V. cholerae CO6, V. cholerae NB2, V. cholerae PC2, S. flexneri SDINT, | MIC, 8–32 μg/mL | [122] |
| Penialidin B (254) | MIC, 4–32 μg/mL | ||||||
| Penialidin C (255) | MIC, 0.50, 16, 8, 0.50 and 8 μg/mL | ||||||
| Citromycetin (256), brefelfin A (258) | MIC, 64–128 μg/mL | ||||||
| p-Hydroxyphenylglyoxalaldoxime (257) | MIC, 32–64 μg/mL | ||||||
| 106 | Purpureocillium lilacinum | roots of Rauvolfia macrophylla | Mount Kalla in the Center Region of Cameroon | Purpureone (259) | B. cereus, L. monocytogenes, E. coli ATCC 8739, K. pneumoniae ATCC 1296, P. stuartii ATCC 29916, P. aeruginosa ATCC PA01 | Zone of inhibition of 10.6, 12.3, 13.0, 8.7, 12.3, and 10.0, mm against (10 μL/6 mm Filter paper disks). | [123] |
| Fusarium | |||||||
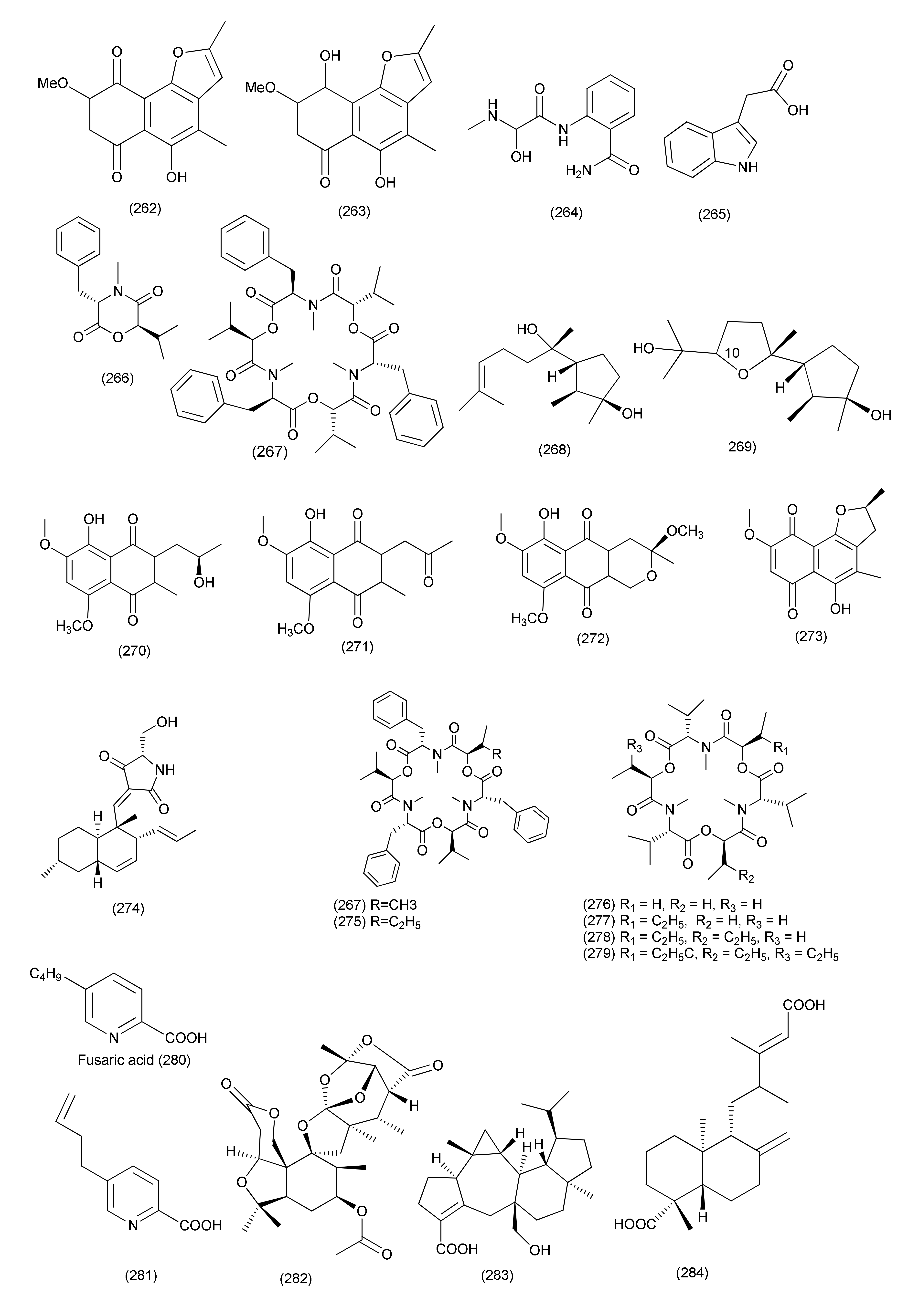
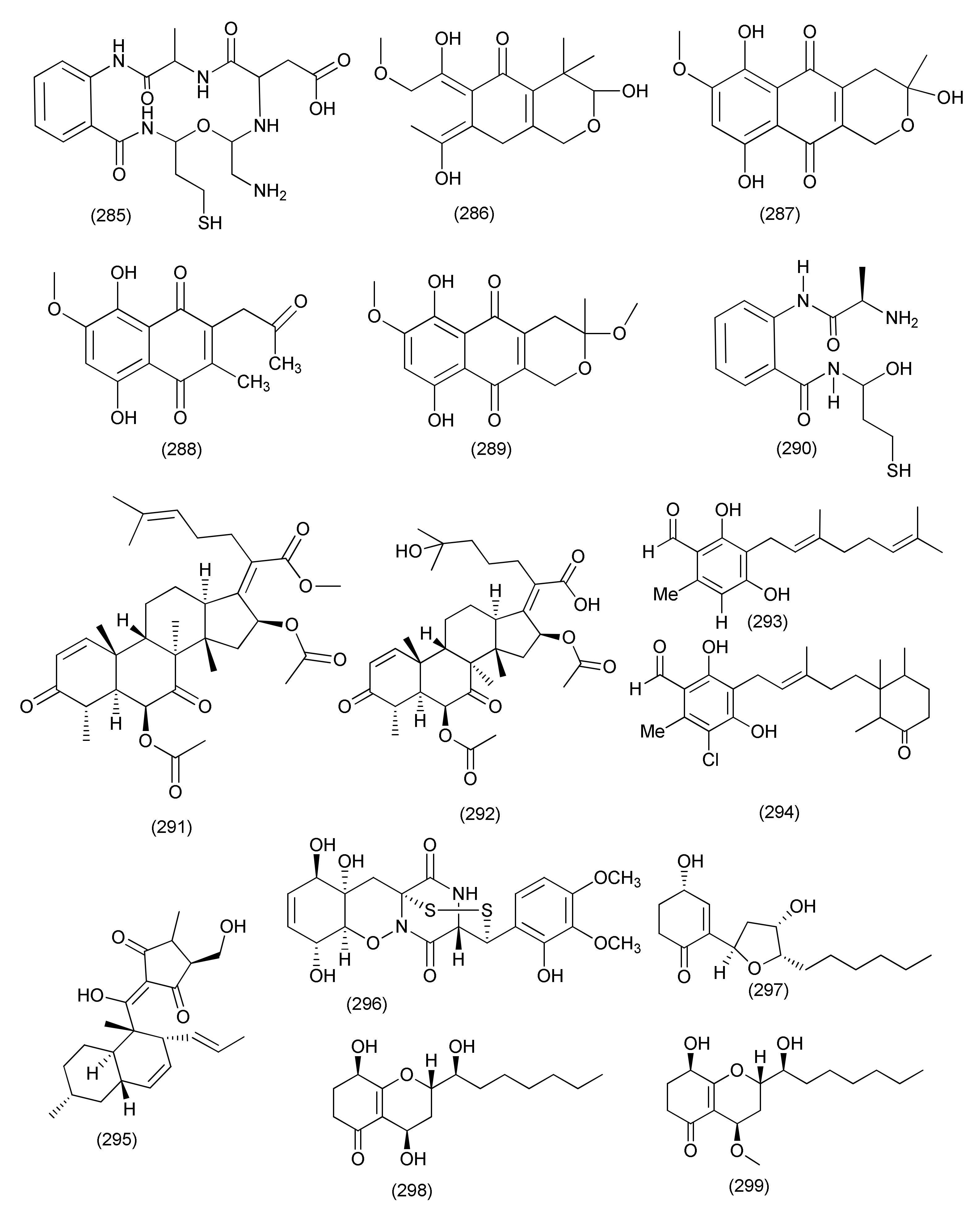
| Neocosmospora sp. MFLUCC 17-0253 | Rhizophora apiculate. | Mixture of 2-methoxy-6-methyl-7-acetonyl-8-hydroxy-1,4-naphthalenedione (260), and 5,8-dihydroxy-7-acetonyl-1,4-naphthalenedione (261) | Acidovorax citrulli | MIC value of 0.0075 mg/mL | [124] | ||
| Anhydrojavanicin (262) | 0.004 mg/mL | ||||||
| Fusarnaphthoquinone (263) | 0.025 mg/mL | ||||||
| 107 | Fusarium sp. | Mentha longifolia | Al Madinah Al Munawwarah, Saudi Arabia. | Fusaribenzamide A (264) | S. aureus and E. coli | MICs, 62.8 and 56.4 μg/disc | [125] |
| 108 | F. proliferatum AF-04 | Green Chinese onion | 5-O-Methylsolaniol (270), 5-O-methyljavanicin (271), methyl ether fusarubin (272), anhydrojavanicin (273) | B. megaterium | MICs 25 μg/mL each. | [126] | |
| 5-O-Methylsolaniol (270), 5-O-methyljavanicin (271), methyl ether fusarubin (272) | B. subtilis | MICs, 50 μg/mL each. | |||||
| Indol-3-acetic acid (265), beauvericin (267), epicyclonerodiol oxide (269) | B. megaterium | MICs 50 μg/mL each | |||||
| Cyclonerodiol (268) | B. megaterium | MIC 12.50 μg/mL. | |||||
| epi-Cyclonerodiol oxide (269), methyl ether fusarubin (272) | E. coli | MIC 50 μg/mL | |||||
| 5-O-Methylsolaniol (270), 5-O-methyljavanicin (271), anhydrojavanicin (273) | E. coli | MIC 25 μg/mL | |||||
| epi-Cyclonerodiol oxide (269), 1,4-naphthoquinones, 5-O-methylsolaniol (270), 5-O-methyljavanicin (271), methyl ether fusarubin (272) | Clostridium perfringens | MICs 50, 50, 12.5 and 50 μg/mL | |||||
| Beauvericin (267), fusaproliferin (106), 5-O-methylsolaniol (270), 5-O-methyljavanicin (271), methyl ether fusarubin (272), anhydrojavanicin (273) | MRSA | MIC value of 50, 50, 12.5, 12.5, 12.5, and 25 μg/mL respectively. | |||||
| 5-O-Methyljavanicin (271), methyl ether fusarubin (272), anhydrojavanicin (273) | RN4220 | MIC value of 50 μg/mL each. | |||||
| Methyl ether fusarubin (272), anhydrojavanicin (273) | NewmanWT | MIC value of 50 μg/mL each. | |||||
| Bassiatin (266) | NewmanWT | MIC, 50 μg/mL | |||||
| 109 | Fusarium sp. TP-G1 | Dendrobium officinable | Chongqing Academy of Chinese Materia Medica in China | Trichosetin (274), beauvericin (267), beauvericin A (275), enniatin H (277), enniatin I (278), enniatin MK1688 (279) | S. aureus and MRSA | IC50 values in the range of 2–32 μg/mL | [127] |
| Enniatin B (276) | S. aureus and MRSA | IC50, 128 μg/mL each | |||||
| Fusaric acid (280), dehydrofusaric acid (281) | Acinetobacter baumannii | MIC, 64 and 128 μg/mL | |||||
| Fusarium sp. YD-2 | Santalum album | Dongguan, Guangdong Province, China | Fusariumin A (282) | S. aureus and P. aeruginosa | MIC, 6.3 μg/mL | [128] | |
| Asperterpenoid A (283) | Salmonella enteritidis and Micrococcus luteus | MIC, 25.2 and 6.3 μg/mL | |||||
| Agathic acid (284) | B. cereus and M. luteus | MIC, 12.5 and 25.4 μg/mL | |||||
| 110 | Fusarium chlamydosporium | Leaves of Anvillea garcinii | Al-Azhar University campus, Egypt | Fusarithioamide B (285) | E. coli, B. cereus, and S. aureus | MIC value of 3.7, 2.5 and 3.1 µg/mL | [129] |
| 111 | Fusarium solani A2 | Glycyrrhiza glabra | Kashmir Himalayas of Jammu and Kashmir State, India | 3,6,9-Trihydroxy-7-methoxy-4,4-dimethyl-3,4-dihydro-1H-benzo[g]-isochromene-5,10-dione (286), fusarubin (287), 3-O-methylfusarubin (288), javanicin (289) | S. aureus (MTCC 96), K. pneumonia (MTCC 109), S. pyogenes (MTCC 442), B. subtilis (MTCC 121), B. cereus (IIIM 25), Micrococcus luteus (MTCC 2470) and E. coli (MTCC 730) | MIC values in the range of <1 to 256 μg/mL. | [130] |
| Fusarubin (287) | Mycobacterium tuberculosis strain H37Rv | MIC, 8 μg/mL, | |||||
| 3,6,9-Trihydroxy-7-methoxy-4,4-dimethyl-3,4-dihydro-1H-benzo[g]-isochromene-5,10-dione (286), 3-O-methylfusarubin (288), javanicin (289) | MIC values of 256, 64, 32 μg/mL | ||||||
| 112 | Fusarium chlamydosporium | Anvillea garcinii | Al-Azhar University, Saudi Arabia | Fusarithioamide A (290) | B. cereus, S. aureus, and E. coli | MICs values of 3.1, 4.4, and 6.9 μg/mL | [131] |
| 113 | Fusarium sp. | Rhoeo spathacea | Pondok Cabe, Banten, Indonesia. | Javanicin (289) | M. tuberculosis and M. phlei | MIC 25 and 50 μg/mL | [132] |
| 114 | Fusarium sp. | Ficus carica | Qinling Mountain, Shaanxi Province, China | Helvolic acid Me ester (291) | B. subtilis, S. aureus, E. coli and P. aeruginosa | MIC, 6.25, 12.5, 6.25, and 3.13 μg/mL | [133] |
| Helvolic acid (100) | MICs 6.25, 6.25, 6.25, and 3.13 μg/mL | ||||||
| hydrohelvolic acid (292) | MICs 6.25, 12.5, 6.25, and 3.13 μg/mL | ||||||
| 115 | Fusarium sp. | - | - | Colletorin B (293), 4,5-dihydroascochlorin (294) | B. megaterium | 5 and 10 mm zone of inhibition at 10 μg/mL concentration of | [134] |
| 116 | Fusarium sp. | Opuntia dillenii | South-Eastern arid zone of Sri Lanka | Equisetin (295) | B. subtilis | MIC, 8 μg/mL | [135] |
| S. aureus and MRSA. | MIC, 16 μg/mL | ||||||
| 117 | Trichoderma harzianum | Zingiber officinale | Banyumas, Central Java, Indonesia | Pretrichodermamide A (296) | M. tuberculosis | MIC, 25 μg/mL (50 μM) | [136] |
| 118 | Trichoderma koningiopsis YIM PH30002 | Panax notoginseng | Koninginin W (297), koninginin D (298), 7-O- and koninginin A (301) | B. subtilis | MIC of 128 μg/mL. | [137] | |
| Koninginin W (297), 7-O-methylkoninginin D (299) | S. typhimurium | MIC, 64 and 128 μg/mL; | |||||
| Koninginin W (297), koninginin (300) | E. coli | MIC of 128 μg/mL. | |||||
| 119 | Trichoderma virens QA-8 | Artemisia argyi | Trichocarotins I–M (302–306), CAF-603 (307), 7β-hydroxy CAF-603 (308), trichocarotins E–H (309–312), and trichocarane A (313) | E. coli EMBLC-1, | MIC values ranging from 0.5 to 32 µg/mL MIC = 0.5 µg/mL | [138] | |
| 7β-Hydroxy CAF-603 (308) | Micrococcus luteus QDIO-3 | ||||||
| 120 | Trichoderma koningiopsis QA-3 | Artemisia argyi. | Trichodermaketone E (314), trichopyranone A (316), 3-hydroxyharziandione (317) and 10,11-dihydro-11-hydroxycyclonerodiol (318), harziandione (321) | E. coli | MIC values ranging from 0.5 to 64 μg/mL | [139] | |
| Trichopyranone A (316), 3-hydroxyharziandione (317), 10,11-dihydro-11-hydroxycyclonerodiol (318), cyclonerodiol (319), 6-(3-hydroxypent-1-en-1-yl)-2H-pyran-2-one (320), harziandione (321) | M. luteus | MIC values ranging from 1 to 16 μg/mL | |||||
| Trichodermaketone E (314), 4-epi-7-O-methylkoninginin D (315), 3-hydroxyharziandione (317), 10,11-dihydro-11-hydroxycyclonerodiol (318), cyclonerodiol (319), 6-(3-hydroxypent-1-en-1-yl)-2H-pyran-2-one (320), harziandione (321) | P. aeruginosa | with MIC values ranging from 4 to 16 μg/mL | |||||
| Trichodermaketone E (314), 10,11-dihydro-11-hydroxycyclonerodiol (318), cyclonerodiol (319), 6-(3-hydroxypent-1-en-1-yl)-2H-pyran-2-one (320), harziandione (321) | V. parahaemolyticus | MIC values ranging from 4 to 16 μg/mL. | |||||
| 3-Hydroxyharziandione (317) | E. coli | MIC value of 0.5 µg/mL | |||||
| 6-(3-Hydroxypent-1-en-1-yl)-2H-pyran-2-one (320) | M. luteus | MIC value of 1 µg/mL | |||||
| 121 | Trichoderma koningiopsis QA-3 | Artemisia argyi | Qichun of the Hubei Province, China | 15-Hydroxy-1,4,5,6-tetra-epi-koninginin G (322) | Vibrio alginolyticus | MIC, 1 μg/mL | [140] |
| Koninginin U (323), 14-ketokoninginin B (324) | Vibrio harveyi and Edwardsiella tarda | MICs 4 and 2 µg/mL | |||||
| 122 | Trichoderma atroviride B7 | Colquhounia coccinea var. mollis | Kunming Botanical Garden, Yunnan, China | Harzianol I (325) | S. aureus, B. subtilis, and M. luteus | EC50 7.7, 7.7, and 9.9 μg/mL | [141] |
| 123 | Trichoderma longibrachiatum MD33 | Dendrobium nobile | Jinshishi, Chishui, China | Dendrobine (326) | Bacillus mycoides, B. subtilis, and Staphylococcus | Zone of inhibition of 9, 12 and 8 mm | [142] |
| 124 | Trichoderma virens QA-8, | Artemisia argyi | Qichun of Hubei Province in central China | Trichocadinins B-D and G (327–330) | E. coli EMBLC-1, Aeromonas hydrophilia QDIO-1, Edwardsiella tarda QDIO-2, E. ictarda QDIO-10, Micrococcus luteus QDIO-3, P. aeruginosa QDIO-4, Vibrio alginolyticus QDIO-5, V. anguillarum QDIO-6, V. harveyi QDIO-7, V. parahemolyticus QDIO-8, and V. vulnificus QDIO-9 | MIC in the range of 8–64 μg/mL | [143] |
| Trichocadinin G (330) | Ed. tarda and V. anguillarum | MIC values of 1 and 2 μg/mL | |||||
| 125 | Trichoderma koningiopsis A729 | Morinda officinalis | Koninginols A-B (331–332) | B. subtilis | MIC values of 10 and 2 μg/mL | [144] | |
| 126 | Trichoderma koningiopsis QA-3 | Artemisia argyi | Qichun | Ent-koninginin A (333) | V. vulnificus | MIC, 4 μg/mL | [145] |
| Ent-koninginin A (333), trichoketide A (339) | E. coli, E. tarda, V. anguillarum, and V. parahemolyticus | MICs ranging from 8 to 64 μg/mL | |||||
| Ent-koninginin A (333), 1,6-di-epi-koninginin A (334), 15-hydroxykoninginin A (335), 10-deacetylkoningiopisin D (336), koninginin T (337), koninginin L (338), trichoketide A (339) | E. coli | MIC, 64 μg/mL each | |||||
| E. tarda, V. alginolyticus, and V. anguillarum | MIC values ranging from 4 to 64 μg/mL | ||||||
| Alternaria | |||||||
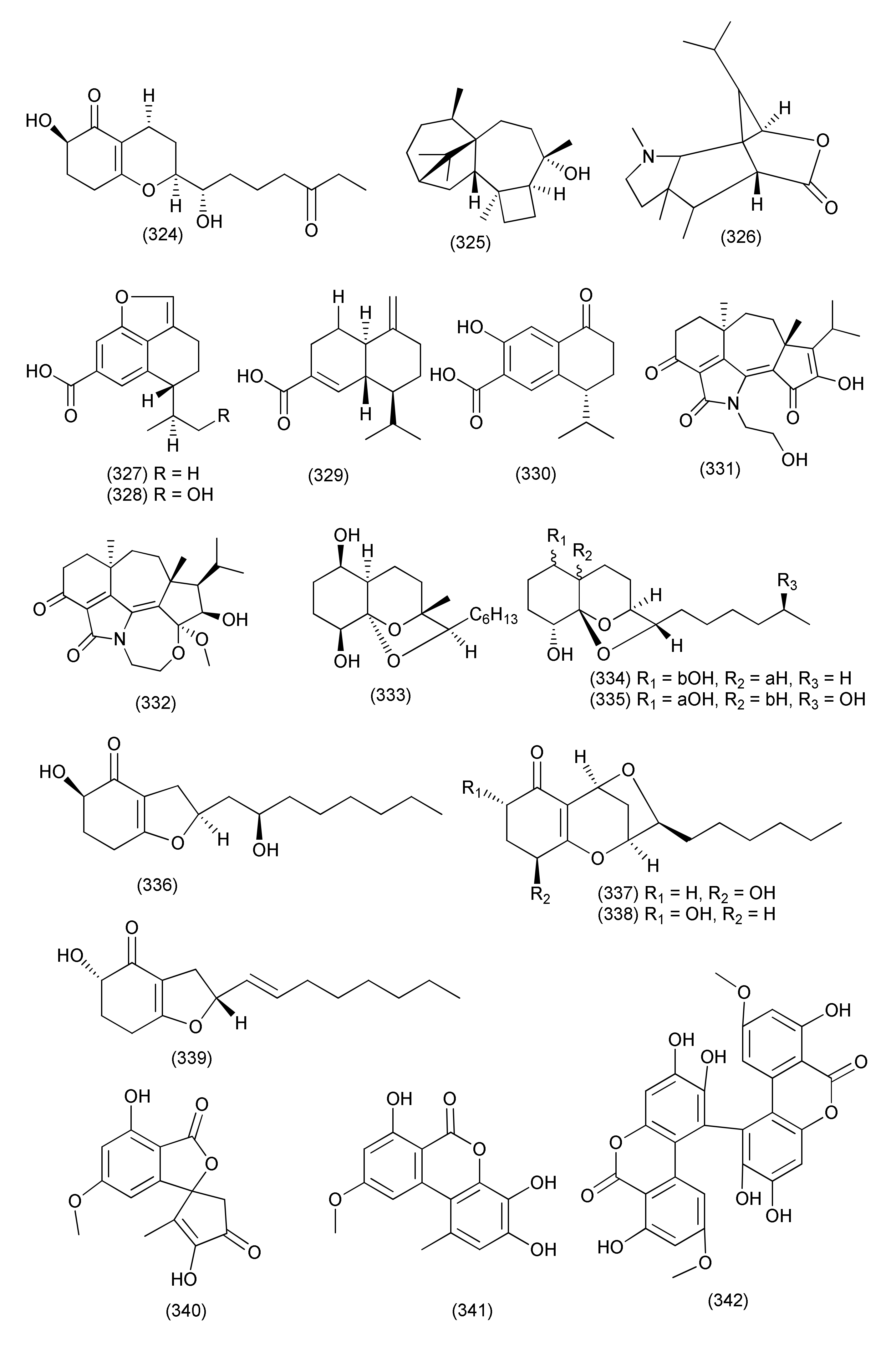
| 127 | Alternaria alternata ZHJG5 | Cercis chinensis | Isotalaroflavone (340), 4-hydroxyalternariol-9-methyl ether (341), verrulactone A (342) | Xanthomonas oryzae pv. Oryzae, Xanthomonas oryzae pv. oryzicola and Ralstonia solanacearum (Rs) | MIC ranging from 0.5 to 64 μg/mL. | [146] | |
| 128 | Alternaria sp. PfuH1 | Pogostemon cablin (Pacholi). | Alternariol (44), altertoxin VII (343), altenuisol (344) | S. agalactiae | MIC, 9.3, 17.3 and 85.3 μg/mL | [147] | |
| Altenuisol (344) | E. coli | MIC, 128 μg/mL | |||||
| 129 | Alternaria alternata ZHJG5 | Cercis chinensis | Alternariol (44), altenuisol (344), alterlactone (345), Dehydroaltenusin (346) | FabH of Xanthomonas oryzae pv. oryzae (Xoo) | IC50 values from 29.5 to 74.1 μM | [148] | |
| Xanthomonas oryzae pv. Oryzae | MIC values from 4 to 64 μg/mL. | ||||||
| Alternariol (44), alterlactone (345) | Rice bacterial leaf blight | a protective efficiency of 66.2 and 82.5% at the concentration of 200 μg/mL | |||||
| 130 | Alternaria alternata MGTMMP031 | Vitex negundo | Madurai, Tamil Nadu, India | Alternariol Me ether (347) | B. cereus, Klebsiella pneumoniae | MIC, 30 µM/L | [149] |
| E. coli, Salmonella typhi, Proteus mirabilis, S. aureus and S. epidermidis | MIC, 35 µM/L | ||||||
| 131 | Alternaria alternata | Grewia asiatica | 3,7-Dihydroxy-9-methoxy-2-methyl-6H-benzo[c]chromen-6-one (348) | S. aureus (ATCC 29213), VRE, and MRSA | MIC, 32, 32 and 8 μg/mL | [150] | |
| Alternariol (44) | S. aureus (ATCC 29213), VRE, and MRSA | MIC, 128, 128, and 64 μg/mL | |||||
| 132 | Alternaria sp. Samif01 | Salvia miltiorrhiza | Beijing Medicinal Plant Garden, Beijing, China | Altenuisol (344), 4-hydroxyalternariol-9-methyl ether (349) and alternariol (44) | A. tumefaciens, B. subtilis, Pseudomonas lachrymans, Ralstonia solanacearum, Staphylococcus hemolyticus and Xanthomonas vesicatorya | MIC values in the range of 86.7–364.7 μM | [151] |
| 133 | Alternaria sp. Samif01 | Salvia miltiorrhiza | Beijing, China | Alternariol 9-Me ether (347) | Bacillus subtilis ATCC 11562 and Staphylococcus haemolyticus ATCC 29970, A. tumefaciens ATCC 11158, Pseudomonas lachrymans ATCC 11921, Ralstonia solanacearum ATCC 11696, and Xanthomonas vesicatoria ATCC 11633 | IC50 values varying from 16.00 to 38.27 g/mL | [152] |
| 134 | Alternaria sp. and Pyrenochaeta sp., | Hydrastis canadensis | William Burch in Hendersonville, North Carolina | Altersetin (350), macrosphelide A (351) | S. aureus | MIC, 0.23, and 75 μg/mL | [153] |
| 135 | Simplicillium lanosoniveum | Hevea brasiliensis | Songkhla Province, Thailand | Simplicildones K (352) | S. aureus ATCC25923, MRSA | MIC, 128μg/mL | [154] |
| Botryorhodine C (353), simplicildones A (354) | S. aureus ATCC25923, MRSA | MIC, 32 μg/mL each | |||||
| 136 | Simplicillium sp. PSU-H41 | Hevea brasiliensis | Songkhla Province, Thailand | Botryorhodine C (353), simplicildone A (354) | S. aureus | MIC, 32 μg/mL each | [155] |
| Botryorhodine C (353) | MRSA | MIC, 32 μg/mL | |||||
| Cladosporium | |||||||
| 137 | Cladosporium cladosporioides | Zygophyllum mandavillei | Al-Ahsa, Saudi Arabia | Isocladosporin (355), 5′- hydroxyasperentin (356), 1-acetyl-17-methoxyaspidospermidin-20-ol (357), and 3-phenylpropionic acid (358) | Xanthomonas oryzae and Pseudomonas syringae | MIC values in the range of 7.81 to 125 µg/mL | [156] |
| 138 | Cladosporium sphaerospermum WBS017 | Fritillaria unibracteata var. wabuensis | Western Sichuan Plateau of China | Cladosin L (359) | S. aureus ATCC 29213 and S. aureus ATCC 700699 | MICs, 50 and 25 mM, | [157] |
| 139 | Cladosporium sp. | Rauwolfia serpentina | Me ether of fusarubin (360) | S. aureus, E. coli, P. aeruginosa and B. megaterium | Zone of inhibition of 27, 25, 24 and 22 mm (40μg/disk) | [158] | |
| Pestalotiopsis | |||||||
| 140 | Pestalotiopsis sp. M-23 | Leucosceptrum canum | Kunming Botanical Garden, China | 11-Dehydro-3a-hydroxyisodrimeninol (361) | B. subtilis | IC50, 280.27 µM | [159] |
| 141 | Pestalotiopsis sp. | Melaleuca quinquenervia | Toohey Forest, Queensland, Australia | (1S,3R)-austrocortirubin (362), (1S,3S)-austrocortirubin (363), 1-deoxyaustrocortirubin (364) | Gram-pos. | 100 μM | [160] |
| 142 | Neopestalotiopsis sp. | Neopestalotins B (365) | B. subtilis, S. aureus, S. pneumoniae | MIC, 10, 20, and 20 μg/mL | [161] | ||
| Phoma | |||||||
| 143 | Phoma cucurbitacearum | Glycyrrhiza glabra | Jammu (J&K). | Thiodiketopiperazine derivatives (366) and (367) | S. aureus and Streptococcus pyogenes | IC50, 10 μM | [162] |
| 144 | Phoma sp. JS752 | Phragmites communis | Seochun, South Korea | Barceloneic acid C (368) | Listeria monocytogenes and Staphylococcus pseudintermedius | MIC, 1.02 μg/mL each | [163] |
| 145 | Setophoma sp., | Psidium guajava fruits | Thielavins T (369), U (370) and V (371) | S. aureus ATCC 25923 | MIC, 6.25, 50, and 25 μg/mL | [164] | |
| Colletotrichum | |||||||
| 146 | Colletotrichum gloeosporioides B12 | Illigera rhodantha | Qionghai City, Hainan Province, China | Colletolides A (372) and B (373), and 3-methyleneisoindolinon (374) | Xanthomonas oryzae pv. oryzae, | MIC, 128 μg/mL each | [165] |
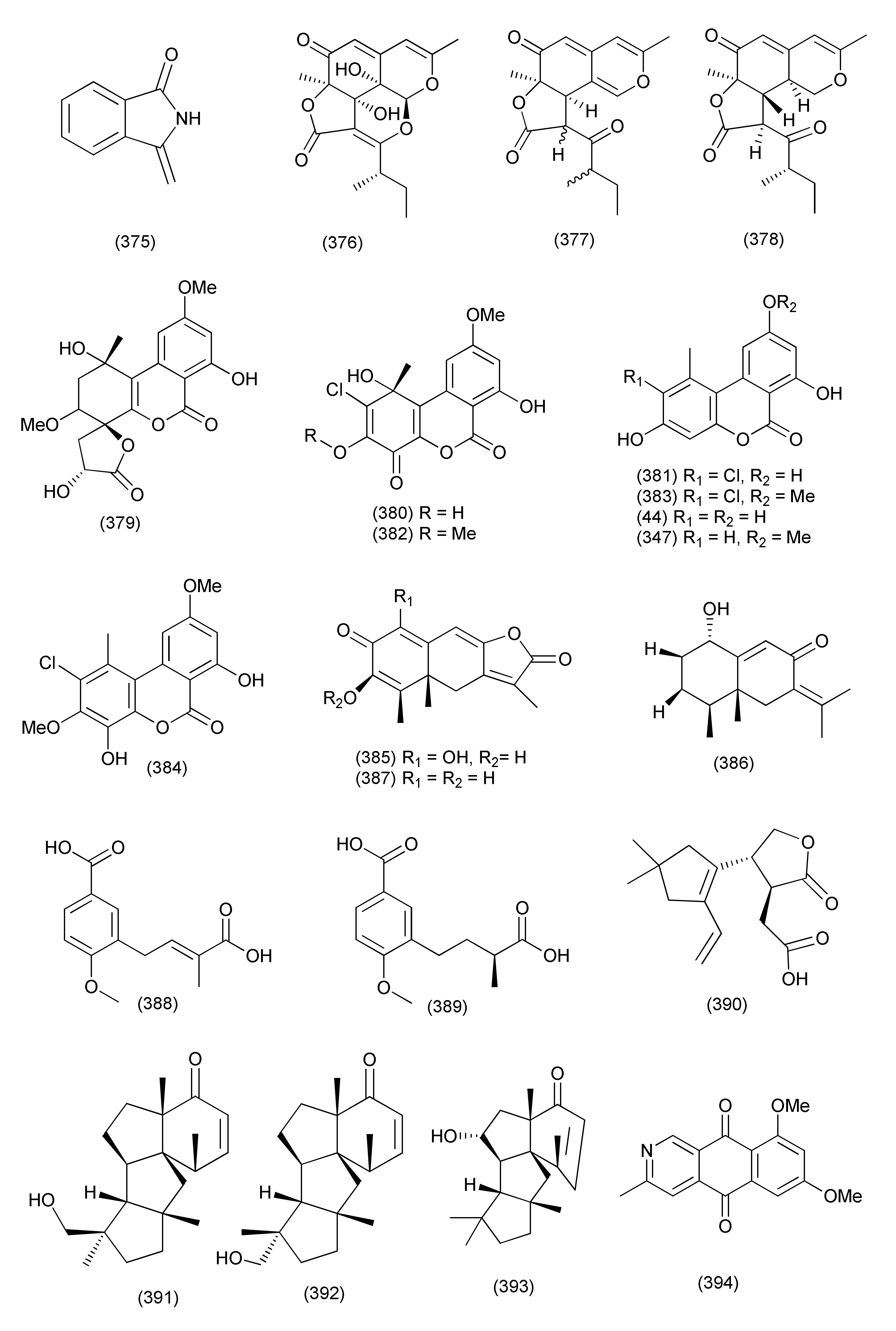
| Sclerone (375) | X. oryzae pv. oryzae | MIC, 64 μg/mL | |||||
| 147 | Colletotrichum sp. BS4 | Buxus sinica | Guangzhou, Guangdong Province, China | Colletotrichones A (376) | E. coli and B. subtilis | MIC, 1.0 and 0.1 μg/mL | [166] |
| Colletotrichone B (377) | S. aureus (DSM 799) | MIC, 5.0 μg/mL | |||||
| Colletotrichone C (378) | E. coli | MIC, 5.0 μg/mL | |||||
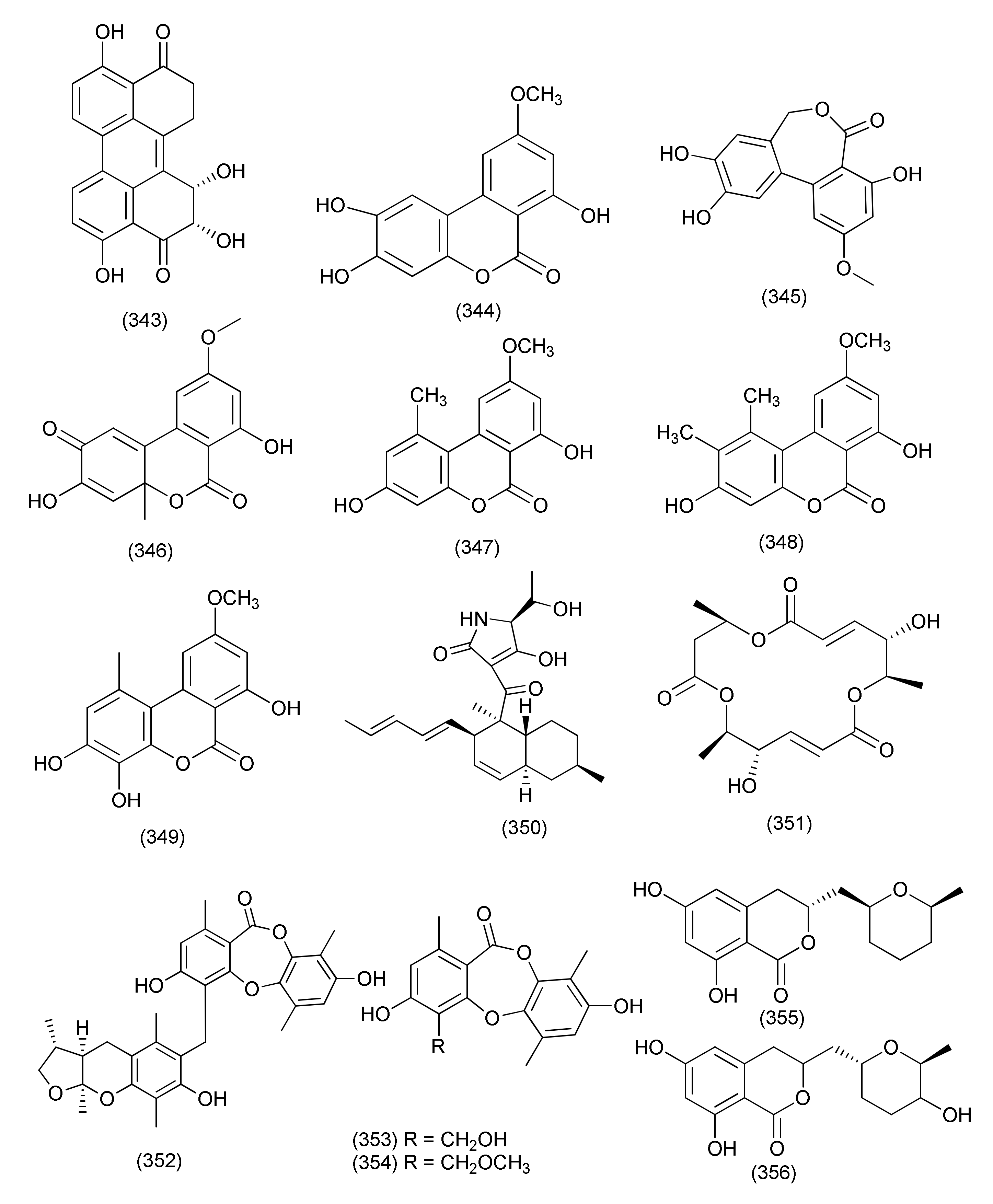
| Minor Taxa of Anamorphic Ascomycetes | |||||||
| 148 | Rhizopycnis vagum Nitaf22 (synonym Acrocalymma vagum) | Nicotiana tabacum | Agricultural University Beijing China | Rhizopycnolide A (379) | A. tumefaciens, B. subtilis, and P. lachrymans | MICs 100, 75, and 100 μg/mL | [167] |
| Rhizopycnin C (380), penicilliumolide D (384), alternariol (44) | A. tumefaciens, B. subtilis, Pseudomonas lachrymans, Ralstonia solanacearum, Staphylococcus hemolyticus, and Xanthomonas vesicatoria, | MICs in the range 25–100 μg/mL | |||||
| Rhizopycnin D (381) | A. tumefaciens, B. subtilis, and R. solanacearum, | MIC 50 μg/mL each, | |||||
| X. vesicatoria | MIC, 75 μg/mL. | ||||||
| Palmariol B (383), Alternariol 9-methyl ether (347) | A. tumefaciens, B. subtilis, P. lachrymans, R. solanacearum, and X. vesicatoria, | IC50 values in the range 16.7−34.3 μg/mL | |||||
| TMC-264 (382) | B. subtilis | MIC 50 μg/mL | |||||
| 149 | Rhizopycnis vagum Nitaf22 (synonym Acrocalymma vagum) | Nicotiana tabacum | China Agricultural University, Beijing | Rhizoperemophilane K (385), 1α-hydroxyhydroisofukinon (386), 2-oxo-3-hydroxyeremophila-1(10),3,7(11), 8-tetraen-8,12-olide (387) | A. tumefaciens, B. subtilis, P. lachrymans, R. solanacearum, S. haemolyticus, and X. vesicatoria, | MIC, 32~128 μg/mL | [168] |
| 150 | Rhizopycnis vagum Nitaf22 (synonym Acrocalymma vagum) | Nicotiana tabacum | China Agricultural University (CAU), Beijing 100101, China | Rhizopycnis acid A (388) | A. tumefaciens, B. subtilis, P. lachrymans, R. solanacearum, S. hemolyticus and X. vesicatoria | MICs, 20.82, 16.11, 23.48, 29.46, 21.11, and 24.31 µg/mL | [169] |
| Rhizopycnis acid B (389) | MICs, 70.89, 81.28, 21.23, 43.40, 67.61, and 34.86 µg/mL | ||||||
| 151 | Leptosphaeria sp. XL026 | Panax notoginseng | Shijiazhuang, Hebei province, China | Leptosphin B (390), conidiogenone C (391), conidiogenone D (392), conidiogenone G (393) | B. cereus | MICs 12.5–6.25 μg/mL | [170] |
| Conidiogenone D (392) | P. aeruginosa | MIC, 12.5 μg/mL | |||||
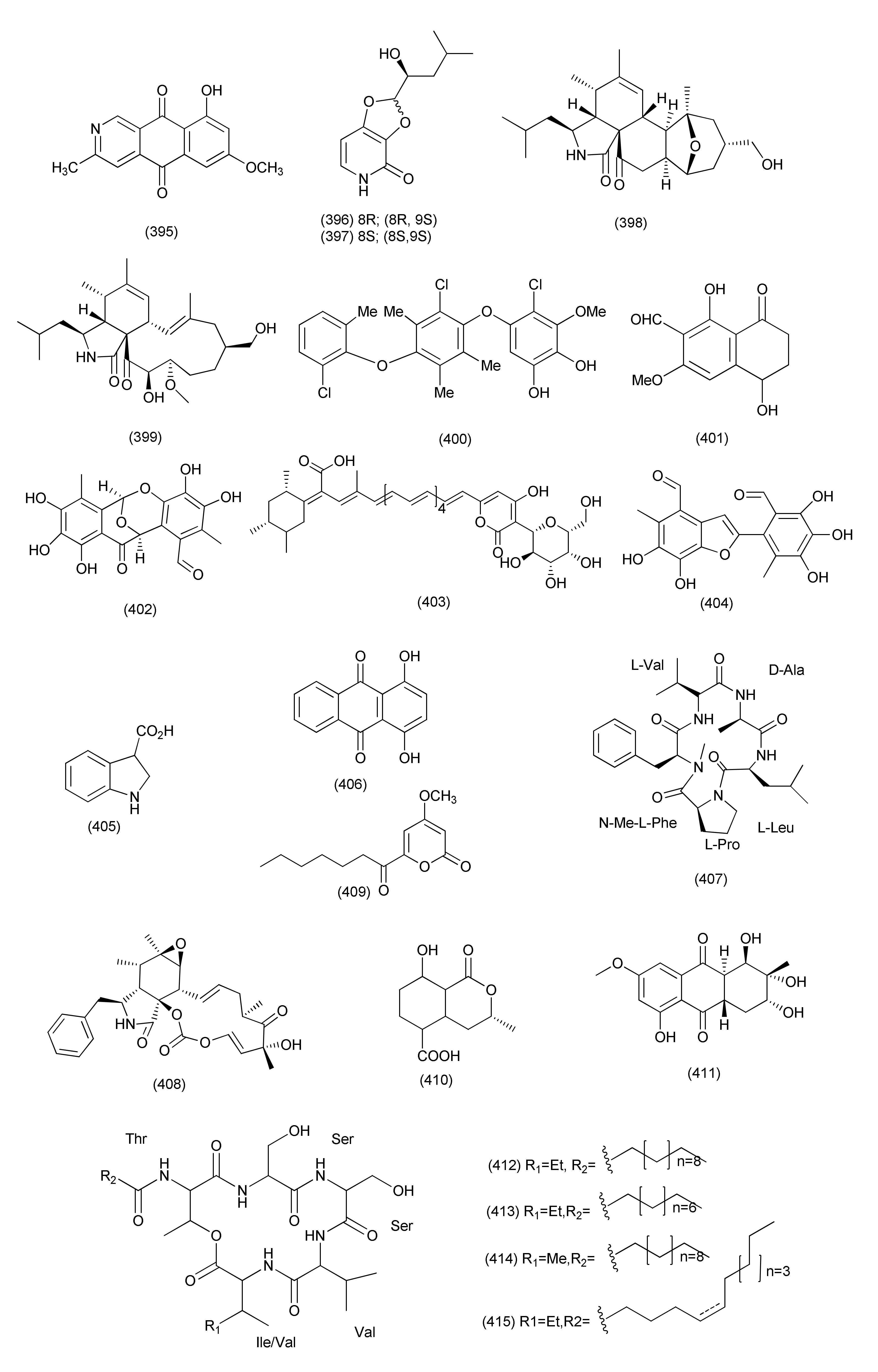
| 152 | Lophiostoma sp. Eef-7 | Eucalyptus exserta. | Scorpinone (394), 5-deoxybostrycoidin (395) | Ralstonia solanacearum | Zone of inhibition of 9.86 and 9.58 mm at 64 µg concentration | [171] | |
| Lophiostoma sp. Sigrf10 | Siraitia grosvenorii | Guangxi Province of China | (8R,9S)-dihydroisoflavipucine (396), (8S,9S)-dihydroisoflavipucine (397) | B. subtilis, A. tumefaciens, Ralstonia solanacearum, and Xanthomonas vesicatoria | IC50 in the range of 35.68–44.85 µM | [172] | |
| 153 | Cytospora chrysosperma | Hippophae rhamnoides | Cytochrysin A (398) | Enterococcus faecium | MIC, 25 μg/mL | [173] | |
| Cytochrysin C (399) | MRSA | MIC, 25 μg/mL | |||||
| 154 | Microsphaeropsis sp. Seimatosporium sp. | Salsola oppositifolia | Gomera, Spain | Microsphaerol (400) | B. megaterium and E. coli, | Zone of inhibition 8 and 9 mm at 0.05 mg concentration | [174] |
| Seimatorone (401) | B. megaterium and E. coli, | Zone of inhibition 3 and 7 (partial) mm at a 0.05 mg concentration | |||||
| 155 | Epicoccum nigrum MK214079 | Salix sp. | Caucasus mountains Lago-Naki, Russia | Epicocconigrone A (402), epipyrone A (403), and epicoccolide B (404) | S. aureus ATCC 29213 | MIC values ranging from 25 to 50 μM | [175] |
| 156 | Epicoccum nigrum | Entada abyssinica | Balatchi (Mbouda), in the West region of Cameroon | p-Hydroxybenzaldehyde (223) | S. aureus, B. cereus, P. aeruginosa, and E. coli | MICs 50, 25, 50, and 25 µg/mL | [176] |
| Beauvericin (267) | S. aureus, B. cereus, and Salmonella typhimurium | MICs 3.12, 12.5, and 12.5 µg/mL | |||||
| Indole-3-carboxylic acid (405) | S. aureus and E. faecalis | MIC values of 6.25 and 50 µg/mL | |||||
| Quinizarin (406) | S. aureus, B. cereus St | MIC values of 50 µg/mL each | |||||
| 157 | Stemphylium lycopersici | S. tonkinensis | Xylapeptide B (407) | B. subtilis, S. aureus and E. coli | MIC, 12.5, 25 and 25 μg/mL | [177] | |
| Cytochalasin E (408) | B. subtilis, S. aureus, B. anthracis, S. dysenteriae, and E. coli | MIC 12.5 to 25 μg/mL | |||||
| 6-Heptanoyl-4-methoxy-2H-pyran2-one (409) | S. paratyphi B | MIC, 12.5 μg/mL | |||||
| (–)-5-Carboxymellein (410) | B. subtilis, S. aureus, B. anthracis, S. dysenteriae, S. paratyphi, E. coli and S. paratyphi B | MIC values from 12.5 to 25 μg/mL | |||||
| 158 | Stemphylium globuliferum, | Juncus acutus | Egypt | Dihydroaltersolanol C (411) | S. aureus | MICs of 49.7 μM | [178] |
| 159 | Lecanicillium sp. (BSNB-SG3.7 Strain) | Sandwithia guyanensis | St Elie, France. | Stephensiolides I (412), D (413), G (414), stephensiolide F (415) | MRSA | MICs 4, 32, 16 and 32 μg/mL | [179] |
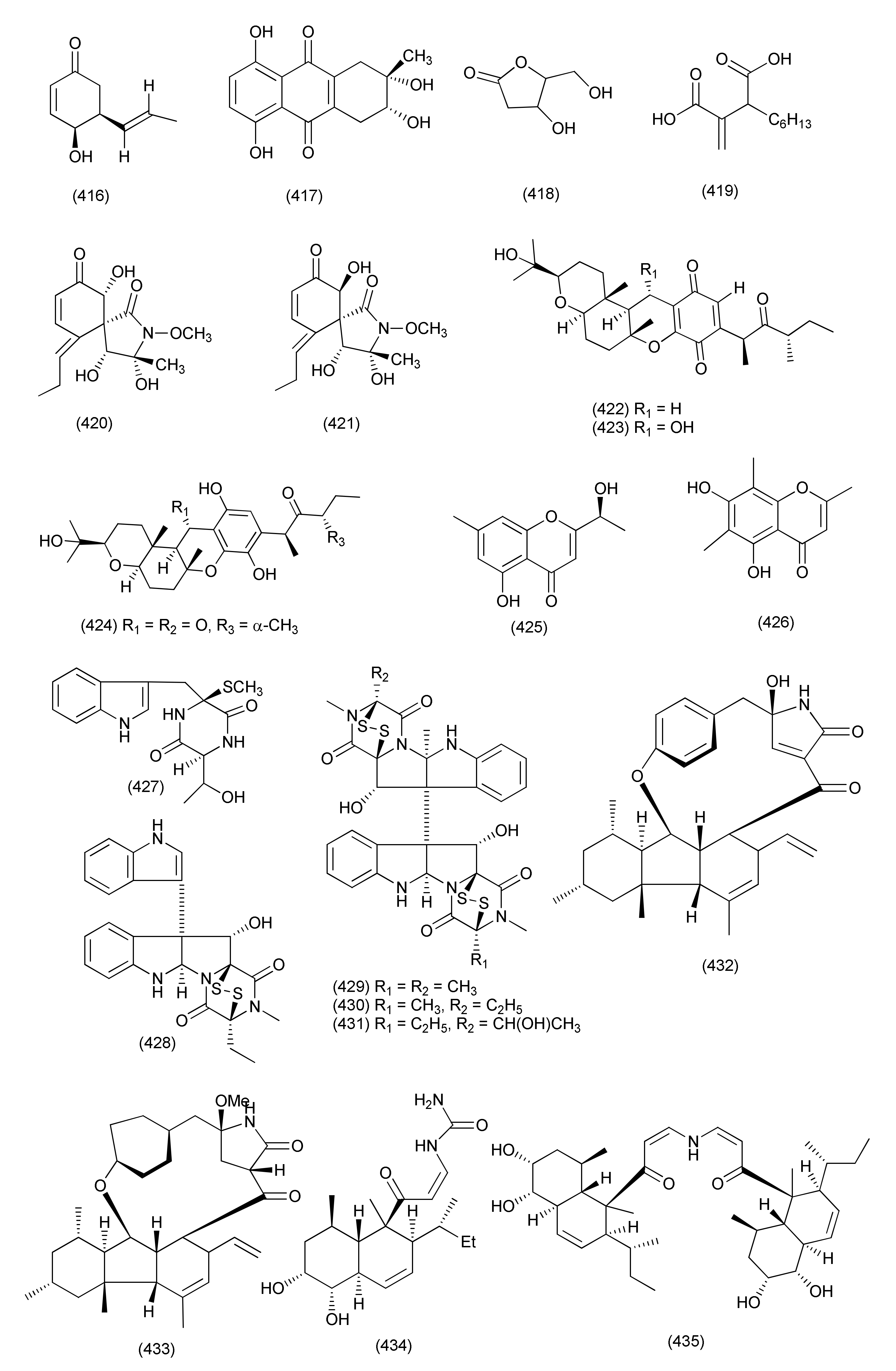
| 160 | Nigrospora sphaerica | Adiantum philippense | Western Ghats region near Virajpete, India | Phomalactone (416) | E. coli and X. campestris | MIC 3.12 μg/mL | [180] |
| S. typhi, B. subtilis, B. cereus, and K. pneumonia | MIC value of 6.25 μg/mL | ||||||
| S. aureus, S. epidermidis, and C. albicans | MIC of 12.5 μg/mL | ||||||
| 161 | Nigrospora sp. BCC 47789 | Choerospondias axillaris | Khao Yai National Park, Nakhon Ratchasima Province, Thailand | Nigrosporone B (417) | M. tuberculosis, B. cereus and E. faecium | MICs 172.25, 21.53 and 10.78 μM | [181] |
| 162 | Curvularia sorghina BRIP 15900) | Rauwolfia macrophylla | Mount Kalla in Cameroon | 2′-Deoxyribolactone (419), hexylitaconic acid (419) | E. coli, Micrococcus luteus, Pseudomonas agarici and Staphylococcus warneri | MIC ranging between 0.17 μg/mL and 0.58 μg/mL | [182] |
| 163 | Curvularia lunata | Paepalanthus chiquitensis | Serra do Cipó, in Minas Gerais State, Brazil | Triticones E (420), F (421) | E. coli, | MIC 62.5 μg/mL | [183] |
| 164 | Bipolaris sp. L1-2 | Lycium barbarum | Ningxia Province, China | Cochlioquinones B (422), C (423), isocochlioquinones (424) | B. subtilis, C. perfringens, and P. viridiflava | MICs 26 μM | [184] |
| 165 | Bipolaris eleusines | Potatoes | nursery of Yunnan Agricultural University, Kunming, Yunnan China | (S)-5-Hydroxy-2-(1-hydroxyethyl)-7-methylchromone (425), 5,7-dihydroxyl-2,6,8-trimethylchromone (426) | Staphylococcus aureus subsp. Aureus | inhibition rates of 56.3 and 32 %, at the concentration of 128 μg/mL | [185] |
| 166 | Bionectria sp. Y1085, | Huperzia serrata | Xichou County, Yunnan Province, China | Bionectin D (427), bionectin E (428), verticillin A (430), sch 52901 (429), gliocladicillin C (431) | E. coli, S. aureus, and S. typhimurium ATCC 6539, | MIC values ranging from 6.25–25 µg/mL | [186] |
| 167 | Cylindrocarpon sp., | Sapium ellipticum | Haut Plateaux region, Cameroon | Pyrrocidine A (432) | S. aureus, ATCC 25923, S. aueus ATCC 700699, S. aueus ATCC 700699, E. faecalis ATCC 29212, E. faecalis ATCC 51299, E. faecium ATCC 35667, E. faecium ATCC 700221 | MIC values ranging from 0.78 to 25 μM | [187] |
| 19-O-Methylpyrrocidine B (433) | S. aureus ATCC25923 and ATCC700699 | MIC, 50 and 25 μM, | |||||
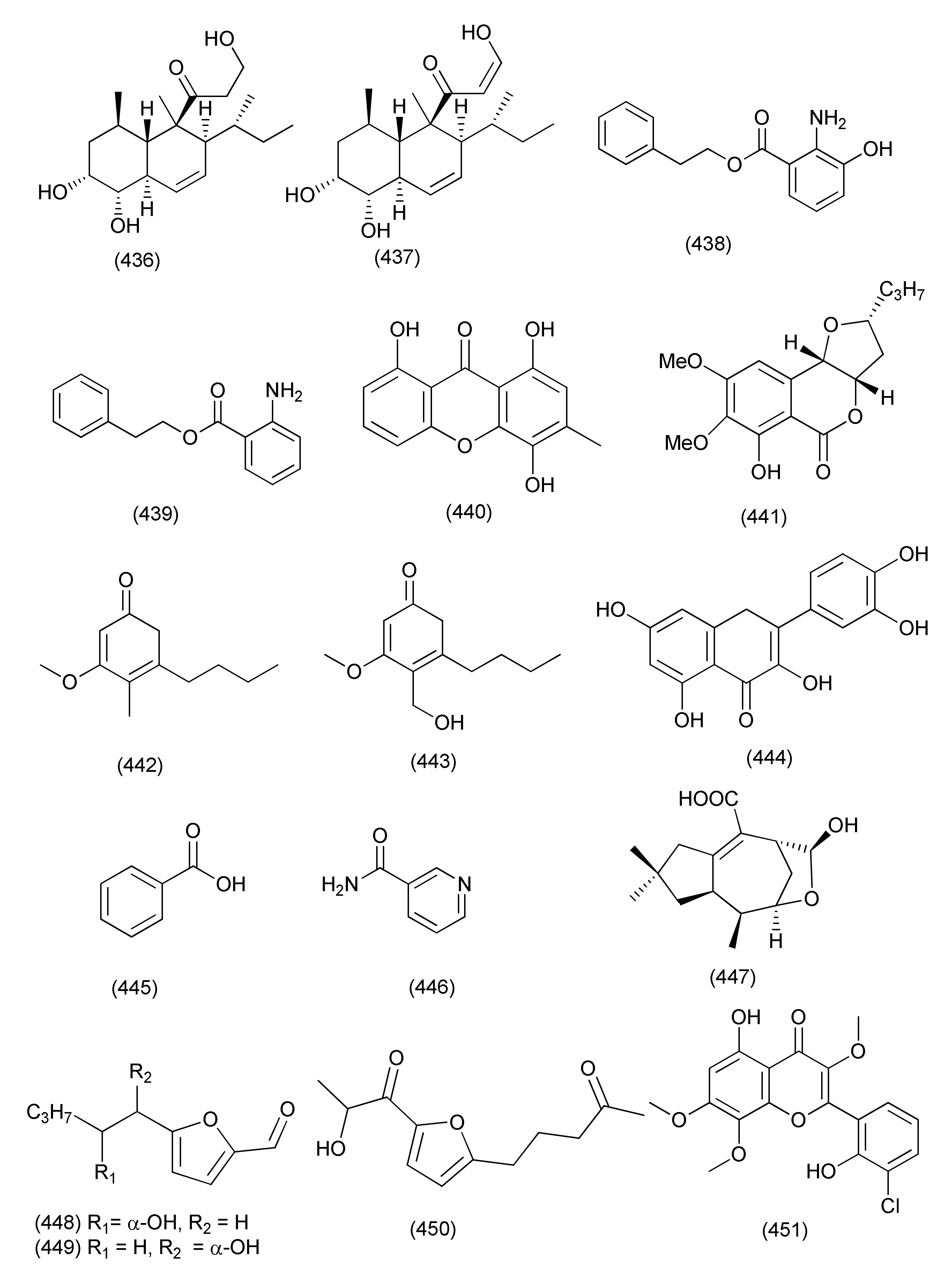
| 168 | Eupenicillium sp. LG41.9 treated with HDAC inhibitor, nicotinamide (15 mg/100 mL) | Xanthium sibiricum | Taian, Shandong Province, China | Eupenicinicol C (434) | [188] | ||
| Eupenicinicol D (435), | S. aureus | MIC 0.1 μg/mL, | |||||
| Eujavanicol A (436) | E. coli | MIC 5.0 μg/mL | |||||
| Eupenicinicol A (437) | |||||||
| 169 | Dendrothyrium variisporum | Globularia alypum | Ain Touta, Batna 05000, Algeria | 2-Phenylethyl 3-hydroxyanthranilate (438) | B. subtilis and M. luteus | MICs 8.33 and 16.66 μg/mL | [189] |
| 2-Phenylethyl anthranilate (439) | B. subtilis and M. luteus | 66.67 μg/mL each | |||||
| 170 | Exserohilum rostratum | Phanera splendens (Kunth) Vaz | Ravenelin (440) | Bacillus subtilis and Staphylococcus aureus | MICs, 7.5 and 484 μM | [190] | |
| 171 | Exserohilum rostratum | Bauhinia guianensis | Monocerin (441) | P. aeruginosa | MIC, 62.5 µg/mL | [191] | |
| Annularin I (442) | E. coli and B. subtilis | MIC, 62.50 and 31.25 µg/mL | |||||
| Annularin J (443) | E. coli and B. subtilis | MIC, 62.50 µg/mL each | |||||
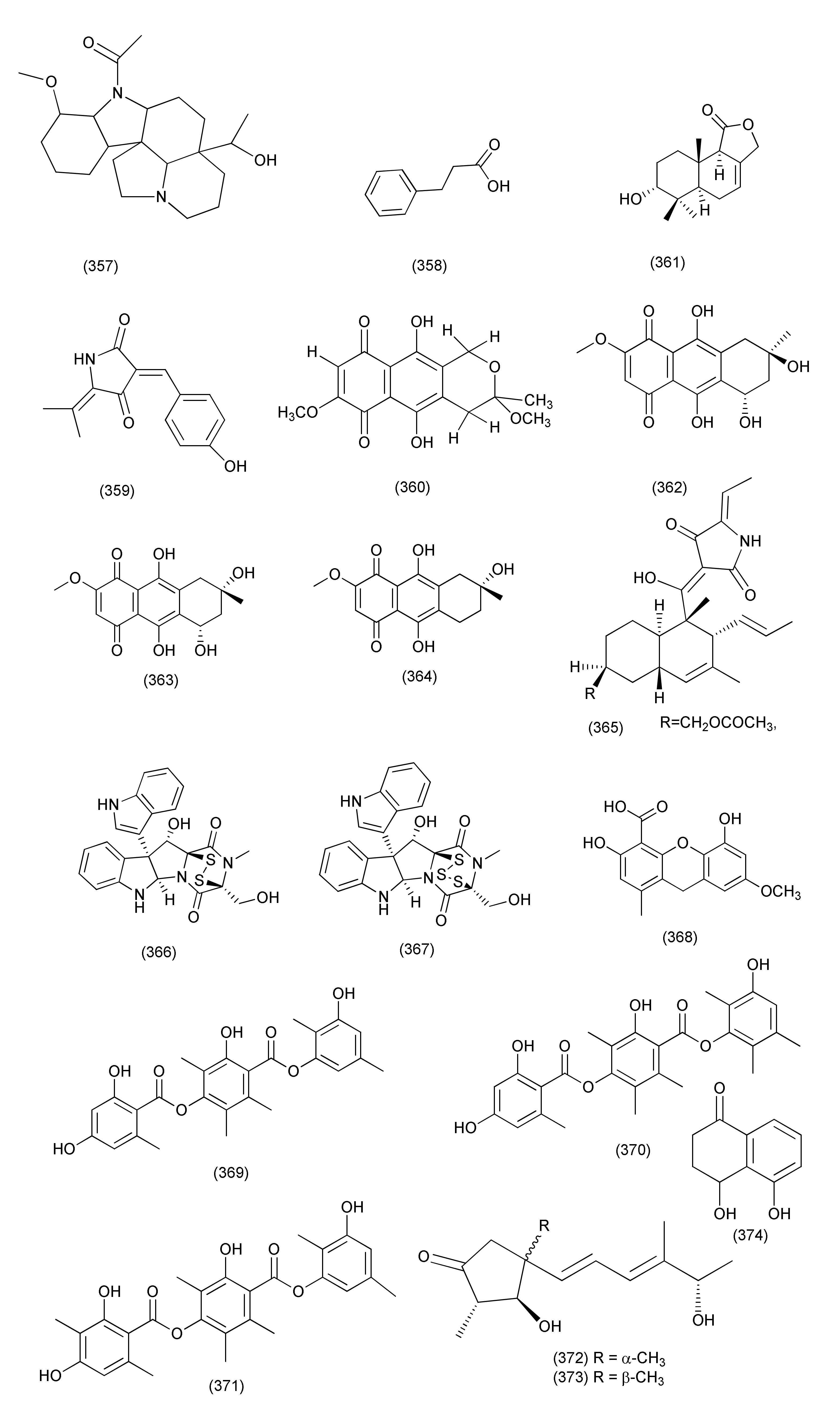
| Basidiomycete | |||||||
| 172 | Psathyrella candolleana | Ginkgo biloba | Quercetin (444), carboxybenzene (445), and nicotinamide (446) | S. aureus | MIC 0.3906, 0.7812 and 6.25 μg/mL | [192] | |
| 173 | Irpex lacteus DR10-1 | Distylium chinense | Banan district of Chongqing in the TGR area, China | Irpexlacte A (447), irpexlacte B-D (448–450) | P. aeruginosa | MIC values ranging from 23.8 to 35.4 μM | [193] |
| Zygomycetes | |||||||
| 174 | Mucor irregularis | Chlorflavonin (451) | [194] | ||||
4. Methods Used for Activation of Silent Biosynthetic Genes
4.1. Epigenetic Modification
4.2. The Co-Culture Strategy
4.3. OSMAC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S. Endophytic fungi: An untapped source for antibacterials. Front. Microbiol. 2015, 5, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, D.; Dussart, F. Selected Fungal Natural Products with Antimicrobial Properties. Molecules 2020, 25, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.C.; Lu, Y.H.; Wang, J.F.; Song, Z.Q.; Hou, Y.G.; Liu, S.S.; Liu, C.S.; Wu, S.H. Bioactive secondary metabolites of the genus Diaporthe and anamorph Phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 2021, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Choi, H.G.; Song, J.H.; Kang, K.S.; Shim, S.H. Bioactive secondary metabolites from an endophytic fungus Phoma sp. PF2 derived from Artemisia princeps Pamp. J. Antibiot. 2019, 72, 174–177. [Google Scholar] [CrossRef]
- El-hawary, S.S.; Moawad, A.S.; Bahr, H.S.; Abdelmohsen, U.R.; Mohammed, R. Natural product diversity from the endophytic fungi of the genus Aspergillus. RSC Adv. 2020, 10, 22058–22079. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Mishra, P.D.; Kulkarni-Almeida, A.; Verekar, S.A.; Sahoo, M.R.; Periyasamy, G.; Goswami, H.; Khanna, A.; Balakrishnan, A.; Vishwakarma, R. Anti-inflammatory and anti-cancer activity of ergoflavin isolated from an endophytic fungus. Chem. Biodivers. 2009, 6, 784–789. [Google Scholar] [CrossRef]
- Martínez-Luis, S.; Cherigo, L.; Arnold, E.; Spadafora, C.; Gerwick, W.H.; Cubilla-Rios, L. Antiparasitic and anticancer constituents of the endophytic fungus Aspergillus sp. strain F1544. Nat. Prod. Commun. 2012, 7, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, S.K.; Verekar, S.A.; Ganguli, B.N. Fungi: An Amazing and Hidden Source of Antimycobacterial compounds. In Fungi: Applications and Management Strategies; Deshmukh, S.K., Misra, J.K., Tiwari, J.P., Papp, T., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 32–60. [Google Scholar]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic Fungi: A Source of Potential Antifungal Compounds. J. Fungi 2018, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Reddy, M.S. Mangrove-associated fungi a novel source of potential anticancer Compounds. J. Fungi 2018, 4, 101. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, S.K.; Agrawala, S.; Gupta, M.K.; Patidar, R.K.; Ranjan, N. Recent advances in the discovery of antiviral metabolites from fungi. Curr. Pharm. Biotechnol. 2022, 23, 495–537. [Google Scholar] [CrossRef]
- Wang, W.X.; Cheng, G.G.; Li, Z.H.; Ai, H.L.; He, J.; Li, J.; Feng, T.; Liu, J.K. Curtachalasins, immunosuppressive agents from the endophytic fungus Xylaria cf. curta. Org. Biomol. Chem. 2019, 17, 7985–7994. [Google Scholar] [CrossRef]
- Bedi, A.; Gupta, M.K.; Conlan, X.A.; Cahill, D.M.; Deshmukh, S.K. Endophytic and marine fungi are potential source of antioxidants. In Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology; Sharma, V.K., Shah, M.P., Parmar, S., Kumar, A., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 23–89. [Google Scholar]
- Toghueo, R.M.K.; Boyom, F.F. Endophytic Penicillium species and their agricultural, biotechnological, and pharmaceutical applications. 3 Biotech 2020, 10, 1–35. [Google Scholar] [CrossRef]
- Toghueo, R.M.K. Bioprospecting endophytic fungi from Fusarium genus as sources of bioactive metabolites. Mycology 2020, 11, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Selvakumar, V.; Panneerselvam, A. Bioactive compounds from endophytic fungi. In Fungi and Their Role in Sustainable Development: Current Perspectives; Gehlot, P., Singh, J., Eds.; Springer: Singapore, 2018; pp. 699–717. [Google Scholar]
- Preethi, K.; Manon Mani, V.; Lavanya, N. Endophytic fungi: A potential source of bioactive compounds for commercial and therapeutic applications. In Endophytes; Patil, R.H., Maheshwari, V.L., Eds.; Springer: Singapore, 2021; pp. 247–272. [Google Scholar]
- Oktavia, L.; Krishna, V.S.; Rekha, E.M.; Fathoni, A.; Sriram, D.; Agusta, A. Anti-mycobacterial activity of two natural Bisanthraquinones:(+)-1, 1′-Bislunatin and (+)-2, 2′-Epicytoskyrin A. In IOP Conference Series: Earth and Environmental Science. IOP Publ. 2020, 591, 12025. [Google Scholar]
- Huang, X.; Zhou, D.; Liang, Y.; Liu, X.; Cao, F.; Qin, Y.; Mo, T.; Xu, Z.; Li, J.; Yang, R. Cytochalasins from endophytic Diaporthe sp. GDG-118. Nat. Prod. Res. 2021, 35, 3396–3403. [Google Scholar] [CrossRef]
- Li, X.M.; Mi, Q.L.; Gao, Q.; Li, J.; Song, C.M.; Zeng, W.L.; Xiang, H.Y.; Liu, X.; Chen, J.H.; Zhang, C.M.; et al. Antibacterial naphthalene derivatives from the fermentation products of the endophytic fungus Phomopsis fukushii. Chem. Nat. Compd. 2021, 57, 293–296. [Google Scholar] [CrossRef]
- Yang, H.Y.; Duan, Y.Q.; Yang, Y.K.; Li, J.; Liu, X.; Ye, L.; Mi, Q.L.; Kong, W.S.; Zhou, M.; Yang, G.Y.; et al. Three new naphthalene derivatives from the endophytic fungus Phomopsis fukushii. Phytochem. Lett. 2017, 22, 266–269. [Google Scholar] [CrossRef]
- Yang, H.Y.; Duan, Y.Q.; Yang, Y.K.; Liu, X.; Ye, L.; Mi, Q.L.; Kong, W.S.; Zhou, M.; Yang, G.Y.; Hu, Q.F.; et al. Two new diphenyl ether derivatives from the fermentation products of an endophytic fungus Phomopsis fukushii. Chem. Nat. Compd. 2019, 55, 428–431. [Google Scholar] [CrossRef]
- Gao, Y.H.; Zheng, R.; Li, J.; Kong, W.S.; Liu, X.; Ye, L.; Mi, Q.L.; Kong, W.S.; Zhou, M.; Yang, G.Y.; et al. Three new diphenyl ether derivatives from the fermentation products of an endophytic fungus Phomopsis fukushii. J. Asian Nat. Prod. Res. 2019, 21, 316–322. [Google Scholar] [CrossRef]
- Li, Z.J.; Yang, H.Y.; Li, J.; Liu, X.; Ye, L.; Kong, W.S.; Tang, S.Y.; Du, G.; Liu, Z.H.; Zhou, M.; et al. Isopentylated diphenyl ether derivatives from the fermentation products of an endophytic fungus Phomopsis fukushii. J. Antibiot. 2018, 71, 359–362. [Google Scholar] [CrossRef]
- Wu, F.; Zhu, Y.N.; Hou, Y.T.; Mi, Q.L.; Chen, J.H.; Zhang, C.M.; Miao, D.; Zhou, M.; Wang, W.G.; Hu, Q.F.; et al. Two new antibacterial anthraquinones from cultures of an endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2021, 57, 823–827. [Google Scholar] [CrossRef]
- Guo, L.; Niu, S.; Chen, S.; Liu, L. Diaporone A, a new antibacterial secondary metabolite from the plant endophytic fungus Diaporthe sp. J. Antibiot. 2020, 73, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.R.; Yang, W.W.; Zhang, X.Q.; Lu, Z.H.; Deng, Z.S.; Guo, Z.Y.; Cao, F.; Zou, K.; Proksch, P. Antibacterial bisabolane sesquiterpenoids and isocoumarin derivatives from the endophytic fungus Phomopsis prunorum. Phytochem. Lett. 2020, 37, 1–4. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Ponomareva, L.V.; Rodrigues, R.; Rohr, J.; Thorson, J.S.; Glienke, C.; Shaaban, K.A. Vochysiamides A and B: Two new bioactive carboxamides produced by the new species Diaporthe vochysiae. Fitoterapia 2019, 138, 104273. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.S.; Liang, M.J.; Mi, Q.L.; Chen, W.; Ling, J.; Chen, X.; Li, J.; Yang, G.Y.; Hu, Q.F.; Wang, W.G.; et al. Two new diphenyl ether derivatives from the fermentation products of the endophytic fungus Phomopsis asparagi. Chem. Nat. Compd. 2019, 55, 843–846. [Google Scholar] [CrossRef]
- Li, X.M.; Zeng, Y.C.; Chen, J.H.; Yang, Y.K.; Li, J.; Ye, L.; Du, G.; Zhou, M.; Hu, Q.F.; Yang, H.Y.; et al. Two new naphthalene derivatives from the fermentation products of an endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2019, 55, 618–621. [Google Scholar] [CrossRef]
- De Medeiros, A.G.; Savi, D.C.; Mitra, P.; Shaaban, K.A.; Jha, A.K.; Thorson, J.S.; Rohr, J.; Glienke, C. Bioprospecting of Diaporthe terebinthifolii LGMF907 for antimicrobial compounds. Folia Microbiol. 2018, 63, 499–505. [Google Scholar] [CrossRef]
- Jayanthi, G.; Arun Babu, R.; Ramachandran, R.; Karthikeyan, K.; Muthumary, J. Production, isolation and structural elucidation of a novel antimicrobial metabolite from the endophytic fungus, Phomopsis/Diaporthe theae. Int. J. Pharm. Biol. Sci. 2018, 8, 8–26. [Google Scholar]
- Kongprapan, T.; Xu, X.; Rukachaisirikul, V.; Phongpaichit, S.; Sakayaroj, J.; Chen, J.; Shen, X. Cytosporone derivatives from the endophytic fungus Phomopsis sp. PSU-H188. Phytochem. Lett. 2017, 22, 219–223. [Google Scholar] [CrossRef]
- Yedukondalu, N.; Arora, P.; Wadhwa, B.; Malik, F.A.; Vishwakarma, R.A.; Gupta, V.K.; Riyaz-Ul-Hassan, S.; Ali, A. Diapolic acid A-B from an endophytic fungus, Diaporthe terebinthifolii depicting antimicrobial and cytotoxic activity. J. Antibiot. 2017, 70, 212–215. [Google Scholar] [CrossRef]
- Sousa, J.P.B.; Aguilar-Pérez, M.M.; Arnold, A.E.; Rios, N.; Coley, P.D.; Kursar, T.A.; Cubilla-Rios, L. Chemical constituents and their antibacterial activity from the tropical endophytic fungus Diaporthe sp. F2934. J. Appl. Microbiol. 2016, 120, 1501–1508. [Google Scholar] [CrossRef] [Green Version]
- Jouda, J.B.; Mbazoa, C.D.; Douala-Meli, C.; Sarkar, P.; Bag, P.K.; Wandji, J. Antibacterial and cytotoxic cytochalasins from the endophytic fungus Phomopsis sp. harbored in Garcinia kola (Heckel) nut. BMC Complement Altern. Med. 2016, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Kusari, S.; Kusari, P.; Kayser, O.; Spiteller, M. Endophytic Diaporthe sp. LG23 produces a potent antibacterial tetracyclic triterpenoid. J. Nat. Prod. 2015, 78, 2128–2132. [Google Scholar] [CrossRef]
- Patridge, E.V.; Darnell, A.; Kucera, K.; Phillips, G.M.; Bokesch, H.R.; Gustafson, K.R.; Spakowicz, D.J.; Zhou, L.; Hungerford, W.M.; Plummer, M.; et al. Pyrrolocin a, a 3-decalinoyltetramic acid with selective biological activity, isolated from Amazonian cultures of the novel endophyte Diaporthales sp. E6927E. Nat. Prod. Commun. 2015, 10, 1649–1654. [Google Scholar] [CrossRef]
- Ibrahim, A.; Tanney, J.B.; Fei, F.; Seifert, K.A.; Cutler, G.C.; Capretta, A.; Miller, J.D.; Sumarah, M.W. Metabolomic-guided discovery of cyclic nonribosomal peptides from Xylaria ellisii sp. nov., a leaf and stem endophyte of Vaccinium angustifolium. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Xu, W.; Liu, C.; Zhou, D.; Liu, X.; Qin, Y.; Cao, F.; Li, J.; Yang, R.; Qin, J. Eremophilane sesquiterpenes from the endophytic fungus Xylaria sp. GDG-102. Nat. Prod. Res. 2019, 33, 1304–1309. [Google Scholar] [CrossRef]
- Zheng, N.; Yao, F.; Liang, X.; Liu, Q.; Xu, W.; Liang, Y.; Liu, X.; Li, J.; Yang, R. A new phthalide from the endophytic fungus Xylaria sp. GDG-102. Nat. Prod. Res. 2018, 32, 755–760. [Google Scholar] [CrossRef]
- Zheng, N.; Liu, Q.; He, D.L.; Liang, Y.; Li, J.; Yang, R.Y. A New compound from the endophytic fungus Xylaria sp. from Sophora tonkinensis. Chem. Nat. Compd. 2018, 54, 447–449. [Google Scholar] [CrossRef]
- Lin, X.; Yu, M.; Lin, T.; Zhang, L. Secondary metabolites of Xylaria sp., an endophytic fungus from Taxus mairei. Nat. Prod. Res. 2016, 30, 2442–2447. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, H.Q.; Zong, S.C.; Gao, J.M.; Zhang, A.L. Chemical and bioactive diversities of the genus Chaetomium secondary metabolites. Mini Rev. Med. Chem. 2012, 12, 127–148. [Google Scholar] [CrossRef]
- Tantapakul, C.; Promgool, T.; Kanokmedhakul, K.; Soytong, K.; Song, J.; Hadsadee, S.; Jungsuttiwong, S.; Kanokmedhakul, S. Bioactive xanthoquinodins and epipolythiodioxopiperazines from Chaetomium globosum 7s-1, an endophytic fungus isolated from Rhapis cochinchinensis (Lour.) Mart. Nat. Prod. Res. 2020, 34, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Hou, S.Y.; Zhang, T.Y.; Wu, Y.Y.; Zhang, M.Y.; Yan, X.M.; Xia, M.Y.; Zhang, Y.X. Cytotoxic and antimicrobial indole alkaloids from an endophytic fungus Chaetomium sp. SYP-F7950 of Panax notoginseng. RSC Adv. 2019, 9, 28754–28763. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhang, D.; Shi, R.; Yang, Z.; Zhao, F.; Tian, Y. Antimicrobial potential of endophytic fungi from Astragalus chinensis. 3 Biotech 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Li, G.; Qiao, Y.N.; Sun, Y.; Peng, X.P.; Lou, H.X. Chamiside A, a cytochalasan with a tricyclic core skeleton from the endophytic fungus Chaetomium nigricolor F5. Org. Lett. 2019, 21, 3319–3322. [Google Scholar] [CrossRef]
- Yang, S.X.; Zhao, W.T.; Chen, H.Y.; Zhang, L.; Liu, T.K.; Chen, H.P.; Yang, J.; Yang, X.L. Aureonitols A and B, Two New C13-Polyketides from Chaetomium globosum, an endophytic fungus in Salvia miltiorrhiza. Chem. Biodivers. 2019, 16, e1900364. [Google Scholar] [CrossRef]
- Ouyang, J.; Mao, Z.; Guo, H.; Xie, Y.; Cui, Z.; Sun, J.; Wu, H.; Wen, X.; Wang, J.; Shan, T. Mollicellins O–R, Four new depsidones isolated from the endophytic fungus Chaetomium sp. Eef-10. Molecules 2018, 23, 3218. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.X.; Chen, Y.; Yang, Y.H.; Li, G.H.; Zhao, P.J. A new epipolythiodioxopiperazine with antibacterial and cytotoxic activities from the endophytic fungus Chaetomium sp. M336. Nat. Prod Res. 2018, 32, 689–694. [Google Scholar] [CrossRef]
- Dissanayake, R.K.; Ratnaweera, P.B.; Williams, D.E.; Wijayarathne, C.D.; Wijesundera, R.L.; Andersen, R.J.; de Silva, E.D. Antimicrobial activities of endophytic fungi of the Sri Lankan aquatic plant Nymphaea nouchali and chaetoglobosin A and C, produced by the endophytic fungus Chaetomium globosum. Mycology 2016, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.T.; Shi, X.; Xian, P.J.; Feng, Z.; Yang, J.; Yang, X.L. A new fusicoccane diterpene and a new polyene from the plant endophytic fungus Talaromyces pinophilus and their antimicrobial activities. Nat. Prod. Res. 2021, 35, 124–130. [Google Scholar] [CrossRef]
- Feng, L.X.; Zhang, B.Y.; Zhu, H.J.; Pan, L.; Cao, F. Bioactive metabolites from Talaromyces purpureogenus, an endophytic fungus from Panax notoginseng. Chem. Nat. Compd. 2020, 56, 974–976. [Google Scholar] [CrossRef]
- Bingyang, Z.; Yangyang, M.; Hua, G.; Huajie, Z.; Wan, L. Absolute configuration determination of two drimane sesquiterpenoids from the endophytic fungi Talaromyces Purpureogenus of Panax notoginseng. Chem. J. Chin. Univ.-Chin. 2017, 38, 1046–1051. [Google Scholar]
- Guo, J.; Ran, H.; Zeng, J.; Liu, D.; Xin, Z. Tafuketide, a phylogeny-guided discovery of a new polyketide from Talaromyces funiculosus Salicorn 58. Appl. Microbiol. Biotechnol. 2016, 100, 5323–5338. [Google Scholar] [CrossRef]
- Zhao, Q.H.; Yang, Z.D.; Shu, Z.M.; Wang, Y.G.; Wang, M.G. Secondary metabolites and biological activities of Talaromyces sp. LGT-2, an endophytic fungus from Tripterygium wilfordii. Iran. J. Pharm Res. 2016, 15, 453–457. [Google Scholar]
- Zhang, S.; Chen, D.; Kuang, M.; Peng, W.; Chen, Y.; Tan, J.; Kang, F.; Xu, K.; Zou, Z. Rhytidhylides A and B, two new phthalide derivatives from the endophytic fungus Rhytidhysteron sp. BZM -9. Molecules 2021, 26, 6092. [Google Scholar] [CrossRef]
- Wang, J.T.; Li, H.Y.; Rao, R.; Yue, J.Y.; Wang, G.K.; Yu, Y. (±)-Stagonosporopsin A, stagonosporopsin B and stagonosporopsin C, antibacterial metabolites produced by endophytic fungus Stagonosporopsis oculihominis. Phytochem. Lett. 2021, 45, 157–160. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, X.; Wang, H.; Chen, Y.; Zhang, J.; Zou, Z.; Tan, H. Antibacterial secondary metabolites from the endophytic fungus Eutypella scoparia SCBG-8. Tetrahedron Lett. 2021, 79, 153314. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, X.; Huo, L.; Zhang, S.; Chen, Y.; Zou, Z.; Tan, H. Sesquiterpenes and steroids from an endophytic Eutypella scoparia. J. Nat. Prod. 2021, 84, 1715–1724. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Du, S.Y.; Ji, B.; Ji, C.J.; Xiao, Y.W.; Yan, R.M.; Zhu, D. New Helvolic Acid derivatives with antibacterial activities from Sarocladium oryzae DX-THL3, an endophytic fungus from Dongxiang wild rice (Oryza rufipogon Griff.). Molecules 2021, 26, 1828. [Google Scholar] [CrossRef]
- Carrieri, R.; Borriello, G.; Piccirillo, G.; Lahoz, E.; Sorrentino, R.; Cermola, M.; Bolletti Censi, S.; Grauso, L.; Mangoni, A.; Vinale, F. Antibiotic Activity of a Paraphaeosphaeria sporulosa -Produced diketopiperazine against Salmonella enterica. J. Fungi 2020, 6, 83. [Google Scholar] [CrossRef]
- Gao, Y.; Stuhldreier, F.; Schmitt, L.; Wesselborg, S.; Wang, L.; Müller, W.E.; Kalscheuer, R.; Guo, Z.; Zou, K.; Liu, Z.; et al. Sesterterpenes and macrolide derivatives from the endophytic fungus Aplosporella javeedii. Fitoterapia 2020, 146, 104652. [Google Scholar] [CrossRef]
- Lai, D.; Mao, Z.; Zhou, Z.; Zhao, S.; Xue, M.; Dai, J.; Zhou, L.; Li, D. New chlamydosporol derivatives from the endophytic fungus Pleosporales sp. Sigrf05 and their cytotoxic and antimicrobial activities. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, L.; Kalscheuer, R.; Liu, Z.; Proksch, P. Antifungal polyketide derivatives from the endophytic fungus Aplosporella javeedii. Bioorg. Med. Chem. 2020, 28, 115456. [Google Scholar] [CrossRef]
- Abbas, Z.; Siddiqui, B.S.; Shahzad, S.; Sattar, S.; Begum, S.; Batool, A.; Choudhary, M.I. Lawsozaheer, a new chromone produced by an endophytic fungus Paecilomyces variotii isolated from Lawsonia Alba Lam. inhibits the growth of Staphylococcus aureus. Nat. Prod. Res. 2021, 35, 4448–4453. [Google Scholar] [CrossRef]
- Chen, H.L.; Zhao, W.T.; Liu, Q.P.; Chen, H.Y.; Zhao, W.; Yang, D.F.; Yang, X.L. (±)-Preisomide: A new alkaloid featuring a rare naturally occurring tetrahydro-2H-1, 2-oxazin skeleton from an endophytic fungus Preussia isomera by using OSMAC strategy. Fitoterapia 2020, 141, 104475. [Google Scholar] [CrossRef]
- Xu, L.L.; Chen, H.L.; Hai, P.; Gao, Y.; Xie, C.D.; Yang, X.L.; Abe, I. (+)-and (−)-Preuisolactone A: A pair of caged norsesquiterpenoidal enantiomers with a tricyclo.4.4. 01, 6.02, 8. decane carbon skeleton from the endophytic fungus Preussia isomera. Org. Lett. 2019, 21, 1078–1081. [Google Scholar] [CrossRef]
- Macabeo, A.P.G.; Cruz, A.J.C.; Narmani, A.; Arzanlou, M.; Babai-Ahari, A.; Pilapil, L.A.E.; Garcia, K.Y.M.; Huch, V.; Stadler, M. Tetrasubstituted α-pyrone derivatives from the endophytic fungus, Neurospora udagawae. Phytochem. Lett. 2020, 35, 147–151. [Google Scholar] [CrossRef]
- Lai, D.; Li, J.; Zhao, S.; Gu, G.; Gong, X.; Proksch, P.; Zhou, L. Chromone and isocoumarin derivatives from the endophytic fungus Xylomelasma sp. Samif07, and their antibacterial and antioxidant activities. Nat. Prod. Res. 2021, 35, 4616–4620. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Kim, S.; Yu, N.H.; Park, A.R.; Yoon, H.; Bae, C.H.; Yeo, J.H.; Kim, I.S.; Kim, J.C. Antimicrobial activities of an oxygenated cyclohexanone derivative isolated from Amphirosellinia nigrospora JS-1675 against various plant pathogenic bacteria and fungi. J. Appl. Microbiol. 2019, 126, 894–904. [Google Scholar] [CrossRef]
- Wu, X.; Pang, X.J.; Xu, L.L.; Zhao, T.; Long, X.Y.; Zhang, Q.Y.; Qin, H.L.; Yang, D.F.; Yang, X.L. Two new alkylated furan derivatives with antifungal and antibacterial activities from the plant endophytic fungus Emericella sp. XL029. Nat. Prod. Res. 2018, 32, 2625–2631. [Google Scholar] [CrossRef]
- Wu, X.; Fang, L.Z.; Liu, F.L.; Pang, X.J.; Qin, H.L.; Zhao, T.; Xu, L.L.; Yang, D.F.; Yang, X.L. New prenylxanthones, polyketide hemiterpenoid pigments from the endophytic fungus Emericella sp. XL029 and their anti-agricultural pathogenic fungal and antibacterial activities. RSC Adv. 2017, 7, 31115–31122. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Z.; Zhang, H.W.; Sun, Z.H.; Dai, J.G.; Hu, Y.C.; Li, R.; Lin, P.C.; Xia, G.Y.; Wang, L.Y.; Qiu, B.L.; et al. Bysspectin A, an unusual octaketide dimer and the precursor derivatives from the endophytic fungus Byssochlamys spectabilis IMM0002 and their biological activities. Eur. J. Med. Chem. 2018, 145, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, D.; Hosoya, T.; Tomoda, H.; Kita, M.; Shigemori, H. Sydowianumols A, B, and C, Three new compounds from discomycete Poculum pseudosydowianum. Chem. Pharm. Bull. 2018, 66, 826–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Yuan, L.Y.; Guo, D.L.; Ye, Y.; Da-Wa, Z.M.; Wang, X.L.; Ma, F.W.; Chen, L.; Gu, Y.C.; Ding, L.S.; et al. Bioactive halogenated dihydroisocoumarins produced by the endophytic fungus Lachnum palmae isolated from Przewalskia tangutica. Phytochemistry 2018, 148, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Sørensen, D.; Jenkins, H.A.; Ejim, L.; Capretta, A.; Sumarah, M.W. Epoxynemanione A, nemanifuranones A–F, and nemanilactones A–C, from Nemania serpens, an endophytic fungus isolated from Riesling grapevines. Phytochemistry 2017, 140, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Amand, S.; Vallet, M.; Guedon, L.; Genta-Jouve, G.; Wien, F.; Mann, S.; Dupont, J.; Prado, S.; Nay, B. A reactive eremophilane and its antibacterial 2 (1 H)-naphthalenone rearrangement product, witnesses of a microbial chemical warfare. Org. Lett. 2017, 19, 4038–4041. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Li, C.; Luo, D.; Teng, P.; Guo, Z.; Tu, X.; Zou, K.; Gong, D. A new cinnamic acid derivative from plant-derived endophytic fungus Pyronema sp. Nat. Prod. Res. 2017, 31, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Wijeratne, E.K.; Xu, Y.; Arnold, A.E.; Gunatilaka, A.L. Pulvinulin A, graminin C, and cis-gregatin B–new natural furanones from Pulvinula sp. 11120, a fungal endophyte of Cupressus arizonica. Nat. Prod. Commun. 2015, 10, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Forcina, G.C.; Castro, A.; Bokesch, H.R.; Spakowicz, D.J.; Legaspi, M.E.; Kucera, K.; Villota, S.; Narva’ez-Trujillo, A.; McMahon, J.B.; Gustafson, K.R.; et al. Stelliosphaerols A and B, sesquiterpene–polyol conjugates from an ecuadorian fungal endophyte. J. Nat. Prod. 2015, 78, 3005–3010. [Google Scholar] [CrossRef]
- Hussain, H.; Jabeen, F.; Krohn, K.; Al-Harrasi, A.; Ahmad, M.; Mabood, F.; Shah, A.; Badshah, A.; Rehman, N.U.; Green, I.R.; et al. Antimicrobial activity of two mellein derivatives isolated from an endophytic fungus. Med. Chem. Res. 2015, 24, 2111–2114. [Google Scholar] [CrossRef]
- Qader, M.; Zaman, K.H.; Hu, Z.; Wang, C.; Wu, X.; Cao, S. Aspochalasin H1: A New Cyclic Aspochalasin from Hawaiian Plant-Associated Endophytic Fungus Aspergillus sp. T1307. Molecules 2021, 26, 4239. [Google Scholar] [CrossRef]
- Wang, M.L.; Chen, R.; Sun, F.J.; Cao, P.R.; Chen, X.R.; Yang, M.H. Three alkaloids and one polyketide from Aspergillus cristatus harbored in Pinellia ternate tubers. Tetrahedron Lett. 2021, 68, 152914. [Google Scholar] [CrossRef]
- Ebada, S.S.; Ebrahim, W. A new antibacterial quinolone derivative from the endophytic fungus Aspergillus versicolor strain Eich. 5.2.2. S. Afr. J. Bot. 2020, 134, 151–155. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.; Asfour, H.Z. Antimicrobial metabolites from the endophytic fungus Aspergillus versicolor. Phytochem. Lett. 2020, 35, 152–155. [Google Scholar] [CrossRef]
- Luo, P.; Shao, G.; Zhang, S.Q.; Zhu, L.; Ding, Z.T.; Cai, L. Secondary metabolites of endophytic fungus Aspergillus ochraceus SX-C7 from Selaginella stauntoniana. Zhongcaoyao 2020, 51, 17–23. [Google Scholar]
- Fathallah, N.; Raafat, M.M.; Issa, M.Y.; Abdel-Aziz, M.M.; Bishr, M.; Abdelkawy, M.A.; Salama, O. Bio-guided fractionation of prenylated benzaldehyde derivatives as potent antimicrobial and antibiofilm from Ammi majus L. fruits-associated Aspergillus amstelodami. Molecules 2019, 24, 4118. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhang, X.; Anbari, W.H.A.; Zhou, Q.; Zhou, P.; Zhang, M.; Zeng, F.; Chen, C.; Tong, Q.; Wang, J.; et al. Cysteine Residue Containing Merocytochalasans and 17, 18-seco-Aspochalasins from Aspergillus micronesiensis. J. Nat. Prod. 2019, 82, 2653–2658. [Google Scholar] [CrossRef]
- Mawabo, I.K.; Nkenfou, C.; Notedji, A.; Jouda, J.B.; Lunga, P.K.; Eke, P.; Fokou, V.T.; Kuiate, J.R. Antimicrobial activities of two secondary metabolites isolated from Aspergillus niger, endophytic fungus harbouring stems of Acanthus montanus. Issues Biol. Sci. Pharm. Res. 2019, 7, 7–15. [Google Scholar]
- Yang, X.F.; Wang, N.N.; Kang, Y.F.; Ma, Y.M. A new furan derivative from an endophytic Aspergillus tubingensis of Decaisnea insignis (Griff.) Hook. f. & Thomson. Nat. Prod. Res. 2019, 33, 2777–2783. [Google Scholar]
- Akhter, N.; Pan, C.; Liu, Y.; Shi, Y.; Wu, B. Isolation and structure determination of a new indene derivative from endophytic fungus Aspergillus flavipes Y-62. Nat. Prod. Res. 2019, 33, 2939–2944. [Google Scholar] [CrossRef]
- Zhang, H.; Ruan, C.; Bai, X.; Chen, J.; Wang, H. Heterocyclic alkaloids as antimicrobial agents of Aspergillus fumigatus D endophytic on Edgeworthia chrysantha. Chem. Nat. Compd. 2018, 54, 411–414. [Google Scholar] [CrossRef]
- Liu, R.; Li, H.; Yang, J.; An, Z. Quinazolinones isolated from Aspergillus sp., an endophytic fungus of Astragalus membranaceus. Chem. Nat. Compd. 2018, 54, 808–810. [Google Scholar] [CrossRef]
- Ola, A.R.; Tawo, B.D.; Belli, H.L.L.; Proksch, P.; Tommy, D.; Hakim, E.H. A new antibacterial polyketide from the endophytic fungi Aspergillus fumigatiaffinis. Nat. Prod. Commun. 2018, 13, 1573–1574. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhang, X.; He, Y.; Sun, W.; Feng, W.; Liu, J.; Hu, Z.; Xu, Q.; Zhu, H.; Zhang, J.; et al. Aspermerodione, a novel fungal metabolite with an unusual 2, 6-dioxabicyclo.2.2. 1. heptane skeleton, as an inhibitor of penicillin-binding protein 2a. Sci. Rep. 2018, 8, 1–11. [Google Scholar]
- Yan, W.; Li, S.J.; Guo, Z.K.; Zhang, W.J.; Wei, W.; Tan, R.X.; Jiao, R.H. New p-terphenyls from the endophytic fungus Aspergillus sp. YXf3. Bioorg. Med. Chem. Lett. 2017, 27, 51–54. [Google Scholar] [CrossRef]
- Gombodorj, S.; Yang, M.H.; Shang, Z.C.; Liu, R.H.; Li, T.X.; Yin, G.P.; Kong, L.Y. New phenalenone derivatives from Pinellia ternata tubers derived Aspergillus sp. Fitoterapia 2017, 120, 72–78. [Google Scholar] [CrossRef]
- Goutam, J.; Sharma, G.; Tiwari, V.K.; Mishra, A.; Kharwar, R.N.; Ramaraj, V.; Koch, B. Isolation and characterization of “terrein” an antimicrobial and antitumor compound from endophytic fungus Aspergillus terreus (JAS-2) associated from Achyranthus aspera Varanasi, India. Front. Microbiol. 2017, 8, 1334. [Google Scholar] [CrossRef] [Green Version]
- Elkhayat, E.S.; Ibrahim, S.R.; Mohamed, G.A.; Ross, S.A. Terrenolide S, a new antileishmanial butenolide from the endophytic fungus Aspergillus terreus. Nat. Prod. Res. 2016, 30, 814–820. [Google Scholar] [CrossRef]
- Ma, Y.M.; Ma, C.C.; Li, T.; Wang, J. A new furan derivative from an endophytic Aspergillus flavus of Cephalotaxus fortunei. Nat. Prod. Res. 2016, 30, 79–84. [Google Scholar] [CrossRef]
- Sadorn, K.; Saepua, S.; Boonyuen, N.; Laksanacharoen, P.; Rachtawee, P.; Prabpai, S.; Kongsaeree, P.; Pittayakhajonwut, P. Allahabadolactones A and B from the endophytic fungus, Aspergillus allahabadii BCC45335. Tetrahedron 2016, 72, 489–495. [Google Scholar] [CrossRef]
- Ma, Y.M.; Li, T.; Ma, C.C. A new pyrone derivative from an endophytic Aspergillus tubingensis of Lycium ruthenicum. Nat. Prod. Res. 2016, 30, 1499–1503. [Google Scholar] [CrossRef]
- Ma, Y.M.; Liang, X.A.; Zhang, H.C.; Liu, R. Cytotoxic and antibiotic cyclic pentapeptide from an endophytic Aspergillus tamarii of Ficus carica. J. Agric. Food Chem. 2016, 64, 3789–3793. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.; Khedr, A.I.M.; Fouad, M.A.; Kotb, M.H.R.; Ross, S.A. Aspernolides F and G, new butyrolactones from the endophytic fungus Aspergillus terreus. Phytochem. Lett. 2015, 14, 84–90. [Google Scholar] [CrossRef]
- Elfita, E.; Munawar, M.; Muharni, M.; Ivantri, I. Chemical constituen from an endophytic fungus Aspergillus sp. (SbD5) isolated from Sambiloto (Andrographis paniculata Nees). Microbiol. Indones. 2015, 9, 6. [Google Scholar]
- Zhang, W.; Wei, W.; Shi, J.; Chen, C.; Zhao, G.; Jiao, R.; Tan, R. Natural phenolic metabolites from endophytic Aspergillus sp. IFB-YXS with antimicrobial activity. Bioorg. Med. Chem. Lett. 2015, 25, 2698–2701. [Google Scholar] [CrossRef]
- Song, H.C.; Qin, D.; Liu, H.Y.; Dong, J.Y.; You, C.; Wang, Y.M. Resorcylic acid lactones produced by an endophytic Penicillium ochrochloron strain from Kadsura angustifolia. Planta Med. 2021, 87, 225–235. [Google Scholar] [CrossRef]
- Syarifah, S.; Elfita, E.; Widjajanti, H.; Setiawan, A.; Kurniawati, A.R. Diversity of endophytic fungi from the root bark of Syzygium zeylanicum, and the antibacterial activity of fungal extracts, and secondary metabolite. Biodivers. J. 2021, 22, 4572–4582. [Google Scholar] [CrossRef]
- Qin, Y.Y.; Huang, X.S.; Liu, X.B.; Mo, T.X.; Xu, Z.L.; Li, B.C.; Qin, X.Y.; Li, J.; Schӓberle, T.F.; Yang, R.Y. Three new andrastin derivatives from the endophytic fungus Penicillium vulpinum. Nat. Prod. Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Peng, C.; Ding, W.; Hu, J.F.; Li, J. Chromenopyridin A, a new N-methoxy-1-pyridone alkaloid from the endophytic fungus Penicillium nothofagi P-6 isolated from the critically endangered conifer Abies beshanzuensis. Nat. Prod. Res. 2020, 1–7. [Google Scholar] [CrossRef]
- Graf, T.N.; Kao, D.; Rivera-Chávez, J.; Gallagher, J.M.; Raja, H.A.; Oberlies, N.H. Drug leads from endophytic fungi: Lessons learned via scaled production. Planta Med. 2020, 86, 988–996. [Google Scholar] [CrossRef]
- Qin, Y.; Liu, X.; Lin, J.; Huang, J.; Jiang, X.; Mo, T.; Xu, Z.; Li, J.; Yang, R. Two new phthalide derivatives from the endophytic fungus Penicillium vulpinum isolated from Sophora tonkinensis. Nat. Prod. Res. 2021, 35, 421–427. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, L.; Zhu, G.; Zuo, M.; Gong, Q.; He, W.; Li, M.; Yuan, C.; Hao, X.; Zhu, W. New phenylpyridone derivatives from the Penicillium sumatrense GZWMJZ-313, a fungal endophyte of Garcinia multiflora. Chin. Chem. Lett. 2019, 30, 431–434. [Google Scholar] [CrossRef]
- Zhao, T.; Xu, L.L.; Zhang, Y.; Lin, Z.H.; Xia, T.; Yang, D.F.; Chen, Y.M.; Yang, X.L. Three new α-pyrone derivatives from the plant endophytic fungus Penicillium ochrochloronthe and their antibacterial, antifungal, and cytotoxic activities. J. Asian Nat. Prod. Res. 2019, 21, 851–858. [Google Scholar] [CrossRef]
- Xie, J.; Wu, Y.Y.; Zhang, T.Y.; Zhang, M.Y.; Peng, F.; Lin, B.; Zhang, Y.X. New antimicrobial compounds produced by endophytic Penicillium janthinellum isolated from Panax notoginseng as potential inhibitors of FtsZ. Fitoterapia 2018, 131, 35–43. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Zhang, T.Y.; Zhang, M.Y.; Cheng, J.; Zhang, Y.X. An endophytic Fungi of Ginkgo biloba L. produces antimicrobial metabolites as potential inhibitors of FtsZ of Staphylococcus aureus. Fitoterapia 2018, 128, 265–271. [Google Scholar] [CrossRef]
- Yang, M.H.; Li, T.X.; Wang, Y.; Liu, R.H.; Luo, J.; Kong, L.Y. Antimicrobial metabolites from the plant endophytic fungus Penicillium sp. Fitoterapia 2017, 116, 72–76. [Google Scholar] [CrossRef]
- Ma, Y.M.; Qiao, K.; Kong, Y.; Li, M.Y.; Guo, L.X.; Miao, Z.; Fan, C. A new isoquinolone alkaloid from an endophytic fungus R22 of Nerium indicum. Nat. Prod. Res. 2017, 31, 951–958. [Google Scholar] [CrossRef]
- Feng, Z.W.; Lv, M.M.; Li, X.S.; Zhang, L.; Liu, C.X.; Guo, Z.Y.; Deng, Z.S.; Zou, K.; Proksch, P. Penicitroamide, an antimicrobial metabolite with high carbonylization from the endophytic fungus Penicillium sp. (No. 24). Molecules 2016, 21, 1438. [Google Scholar] [CrossRef] [Green Version]
- Jouda, J.B.; Mbazoa, C.D.; Sarkar, P.; Bag, P.K.; Wandji, J. Anticancer and antibacterial secondary metabolites from the endophytic fungus Penicillium sp. CAM64 against multi-drug resistant Gram-negative bacteria. Afr. Health Sci. 2016, 16, 734–743. [Google Scholar] [CrossRef]
- Lenta, B.N.; Ngatchou, J.; Frese, M.; Ladoh-Yemeda, F.; Voundi, S.; Nardella, F.; Michalek, C.; Wibberg, D.; Ngouela, S.; Tsamo, E.; et al. Purpureone, an antileishmanial ergochrome from the endophytic fungus Purpureocillium lilacinum. Z. Naturforsch. B. 2016, 71, 1159–1167. [Google Scholar] [CrossRef]
- Klomchit, A.; Calderin, J.D.; Jaidee, W.; Watla-Iad, K.; Brooks, S. Napthoquinones from Neocosmospora sp.—Antibiotic Activity against Acidovorax citrulli, the Causative Agent of Bacterial Fruit Blotch in Watermelon and Melon. J. Fungi 2021, 7, 370. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khayat, M.T.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F. A new antifungal aminobenzamide derivative from the endophytic fungus Fusarium sp. Pharmacogn. Mag. 2019, 15, 204–207. [Google Scholar] [CrossRef]
- Jiang, C.X.; Li, J.; Zhang, J.M.; Jin, X.J.; Yu, B.; Fang, J.G.; Wu, Q.X. Isolation, identification, and activity evaluation of chemical constituents from soil fungus Fusarium avenaceum SF-1502 and endophytic fungus Fusarium proliferatum AF-04. J. Agric. Food Chem. 2019, 67, 1839–1846. [Google Scholar] [CrossRef]
- Shi, S.; Li, Y.; Ming, Y.; Li, C.; Li, Z.; Chen, J.; Luo, M. Biological activity and chemical composition of the endophytic fungus Fusarium sp. TP-G1 obtained from the root of Dendrobium officinale Kimura et Migo. Rec. Nat. Prod. 2018, 12, 549–556. [Google Scholar] [CrossRef]
- Yan, C.; Liu, W.; Li, J.; Deng, Y.; Chen, S.; Liu, H. Bioactive terpenoids from Santalum album derived endophytic fungus Fusarium sp. YD-2. RSC Adv. 2018, 8, 14823–14828. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.R.; Mohamed, G.A.; Al Haidari, R.A.; Zayed, M.F.; El-Kholy, A.A.; Elkhayat, E.S.; Ross, S.A. Fusarithioamide B, a new benzamide derivative from the endophytic fungus Fusarium chlamydosporium with potent cytotoxic and antimicrobial activities. Bioorg. Med. Chem. 2018, 26, 786–790. [Google Scholar] [CrossRef]
- Shah, A.; Rather, M.A.; Hassan, Q.P.; Aga, M.A.; Mushtaq, S.; Shah, A.M.; Hussain, A.; Baba, S.A.; Ahmad, Z. Discovery of anti-microbial and anti-tubercular molecules from Fusarium solani: An endophyte of Glycyrrhiza glabra. J. Appl. Microbiol. 2017, 122, 1168–1176. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.A.; Fat’hi, S.M.; Ross, S.A. Fusarithioamide A, a new antimicrobial and cytotoxic benzamide derivative from the endophytic fungus Fusarium chlamydosporium. Biochem. Biophys Res. Commun. 2016, 479, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Alvin, A.; Kalaitzis, J.A.; Sasia, B.; Neilan, B.A. Combined genetic and bioactivity-based prioritization leads to the isolation of an endophyte-derived antimycobacterial compound. J. Appl. Microbiol. 2016, 120, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.A.; Ma, Y.M.; Zhang, H.C.; Liu, R. A new helvolic acid derivative from an endophytic Fusarium sp. of Ficus carica. Nat. Prod. Res. 2016, 30, 2407–2412. [Google Scholar] [CrossRef]
- Hussain, H.; Drogies, K.H.; Al-Harrasi, A.; Hassan, Z.; Shah, A.; Rana, U.A.; Green, I.R.; Draeger, S.; Schulz, B.; Krohn, K. Antimicrobial constituents from endophytic fungus Fusarium sp. Asian Pac. J. Trop. Dis. 2015, 5, 186–189. [Google Scholar] [CrossRef]
- Ratnaweera, P.B.; de Silva, E.D.; Williams, D.E.; Andersen, R.J. Antimicrobial activities of endophytic fungi obtained from the arid zone invasive plant Opuntia dillenii and the isolation of equisetin, from endophytic Fusarium sp. BMC Complement. Altern. Med. 2015, 15, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Harwoko, H.; Daletos, G.; Stuhldreier, F.; Lee, J.; Wesselborg, S.; Feldbrügge, M.; Müller, W.E.; Kalscheuer, R.; Ancheeva, E.; Proksch, P. Dithiodiketopiperazine derivatives from endophytic fungi Trichoderma harzianum and Epicoccum nigrum. Nat. Prod. Res. 2021, 35, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Hu, B.Y.; Qian, M.A.; Wang, Z.H.; Zou, J.M.; Sang, X.Y.; Li, L.; Luo, X.D.; Zhao, L.X. Koninginin W, a new polyketide from the endophytic fungus Trichoderma koningiopsis YIM PH30002. Chem. Biodivers. 2021, 18, e2100460. [Google Scholar] [CrossRef]
- Shi, X.S.; Song, Y.P.; Meng, L.H.; Yang, S.Q.; Wang, D.J.; Zhou, X.W.; Ji, N.Y.; Wang, B.G.; Li, X.M. Isolation and Characterization of antibacterial carotane sesquiterpenes from Artemisia argyi associated endophytic Trichoderma virens QA-8. Antibiotics 2021, 10, 213. [Google Scholar] [CrossRef]
- Shi, X.S.; Meng, L.H.; Li, X.; Wang, D.J.; Zhou, X.W.; Du, F.Y.; Wang, B.G.; Li, X.M. Polyketides and Terpenoids with Potent Antibacterial Activities from the Artemisia argyi-Derived Fungus Trichoderma koningiopsis QA-3. Chem. Biodivers. 2020, 17, e2000566. [Google Scholar] [CrossRef]
- Shi, X.S.; Li, H.L.; Li, X.M.; Wang, D.J.; Li, X.; Meng, L.H.; Zhou, X.W.; Wang, B.G. Highly oxygenated polyketides produced by Trichoderma koningiopsis QA-3, an endophytic fungus obtained from the fresh roots of the medicinal plant Artemisia argyi. Bioorg. Chem. 2020, 94, 103448. [Google Scholar] [CrossRef]
- Li, W.Y.; Liu, Y.; Lin, Y.T.; Liu, Y.C.; Guo, K.; Li, X.N.; Luo, S.H.; Li, S.H. Antibacterial harziane diterpenoids from a fungal symbiont Trichoderma atroviride isolated from Colquhounia coccinea var. mollis. Phytochemistry 2020, 170, 112198. [Google Scholar] [CrossRef]
- Sarsaiya, S.; Jain, A.; Fan, X.; Jia, Q.; Xu, Q.; Shu, F.; Zhou, Q.; Shi, J.; Chen, J. New insights into detection of Front Microbiol, 11a dendrobine compound from a novel endophytic Trichoderma longibrachiatum strain and its toxicity against phytopathogenic bacteria. Front. Microbiol. 2020, 11, 337. [Google Scholar] [CrossRef]
- Shi, X.S.; Meng, L.H.; Li, X.M.; Li, X.; Wang, D.J.; Li, H.L.; Zhou, X.W.; Wang, B.G. Trichocadinins B–G: Antimicrobial cadinane sesquiterpenes from Trichoderma virens QA-8, an endophytic fungus obtained from the medicinal plant Artemisia argyi. J. Nat. Prod. 2019, 82, 2470–2476. [Google Scholar] [CrossRef]
- Chen, S.; Li, H.; Chen, Y.; Li, S.; Xu, J.; Guo, H.; Liu, Z.; Zhu, S.; Liu, H.; Zhang, W. Three new diterpenes and two new sesquiterpenoids from the endophytic fungus Trichoderma koningiopsis A729. Bioorg. Chem. 2019, 86, 368–374. [Google Scholar] [CrossRef]
- Shi, X.S.; Wang, D.J.; Li, X.M.; Li, H.L.; Meng, L.H.; Li, X.; Pi, Y.; Zhou, X.W.; Wang, B.G. Antimicrobial polyketides from Trichoderma koningiopsis QA-3, an endophytic fungus obtained from the medicinal plant Artemisia argyi. Rsc. Adv. 2017, 7, 51335–51342. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wang, B.; Tian, K.; Ji, W.; Zhang, T.; Ping, C.; Yan, W.; Ye, Y. Novel metabolites from the Cercis chinensis derived endophytic fungus Alternaria alternata ZHJG5 and their antibacterial activities. Pest Manag. Sci. 2021, 77, 2264–2271. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.D.; Yi, T.F.; Ma, Q.Y.; Xie, Q.Y.; Zhou, L.M.; Chen, J.P.; Dai, H.F.; Wu, Y.G.; Zhao, Y.X. Biphenyl metabolites from the patchouli endophytic fungus Alternaria sp. PfuH1. Fitoterapia 2020, 146, 104708. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xiao, C.; Wang, J.; Tian, K.; Ji, W.; Yang, T.; Khan, B.; Qian, G.; Yan, W.; Ye, Y. Discovery of natural FabH inhibitors using an immobilized enzyme column and their antibacterial activity against Xanthomonas oryzae pv. oryzae. J. Agric. Food Chem. 2020, 68, 14204–14211. [Google Scholar] [CrossRef]
- Palanichamy, P.; Kannan, S.; Murugan, D.; Alagusundaram, P.; Marudhamuthu, M. Purification, crystallization and anticancer activity evaluation of the compound alternariol methyl ether from endophytic fungi Alternaria alternata. J. Appl. Microbiol. 2019, 127, 1468–1478. [Google Scholar] [CrossRef]
- Deshidi, R.; Devari, S.; Kushwaha, M.; Gupta, A.P.; Sharma, R.; Chib, R.; Khan, I.A.; Jaglan, S.; Shah, B.A. Isolation and quantification of alternariols from endophytic fungus, Alternaria alternata: LC-ESI-MS/MS analysis. ChemistrySelect 2017, 2, 364–368. [Google Scholar] [CrossRef]
- Tian, J.; Fu, L.; Zhang, Z.; Dong, X.; Xu, D.; Mao, Z.; Liu, Y.; Lai, D.; Zhou, L. Dibenzo-α-pyrones from the endophytic fungus Alternaria sp. Samif01: Isolation, structure elucidation, and their antibacterial and antioxidant activities. Nat. Prod. Res. 2017, 31, 387–396. [Google Scholar] [CrossRef]
- Lou, J.; Yu, R.; Wang, X.; Mao, Z.; Fu, L.; Liu, Y.; Zhou, L. Alternariol 9-methyl ether from the endophytic fungus Alternaria sp. Samif01 and its bioactivities. Braz. J. Microbiol. 2016, 47, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, J.J.; Todd, D.A.; Egan, J.M.; Raja, H.A.; Oberlies, N.H.; Kvalheim, O.M.; Cech, N.B. Biochemometrics for natural products research: Comparison of data analysis approaches and application to identification of bioactive compounds. J. Nat. Prod. 2016, 79, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Rukachaisirikul, V.; Chinpha, S.; Saetang, P.; Phongpaichit, S.; Jungsuttiwong, S.; Hadsadee, S.; Sakayaroj, J.; Preedanon, S.; Temkitthawon, P.; Ingkaninan, K. Depsidones and a dihydroxanthenone from the endophytic fungi Simplicillium lanosoniveum (JFH Beyma) Zare & W. Gams PSU-H168 and PSU-H261. Fitoterapia 2019, 138, 104286. [Google Scholar]
- Saetang, P.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J.; Borwornpinyo, S.; Seemakhan, S.; Muanprasat, C. Depsidones and an α-pyrone derivative from Simpilcillium sp. PSU-H41, an endophytic fungus from Hevea brasiliensis leaf. Phytochemistry 2017, 143, 115–123. [Google Scholar] [CrossRef]
- Yehia, R.S.; Osman, G.H.; Assaggaf, H.; Salem, R.; Mohamed, M.S. Isolation of potential antimicrobial metabolites from endophytic fungus Cladosporium cladosporioides from endemic plant Zygophyllum mandavillei. S. Afr. J. Bot. 2020, 134, 296–302. [Google Scholar] [CrossRef]
- Pan, F.; El-Kashef, D.H.; Kalscheuer, R.; Müller, W.E.; Lee, J.; Feldbrügge, M.; Mándi, A.; Kurtán, T.; Liu, Z.; Wu, W.; et al. Cladosins LO, new hybrid polyketides from the endophytic fungus Cladosporium sphaerospermum WBS017. Eur. J. Med. Chem. 2020, 191, 112159. [Google Scholar] [CrossRef]
- Khan, M.I.H.; Sohrab, M.H.; Rony, S.R.; Tareq, F.S.; Hasan, C.M.; Mazid, M.A. Cytotoxic and antibacterial naphthoquinones from an endophytic fungus, Cladosporium sp. Toxicol. Rep. 2016, 3, 861–865. [Google Scholar] [CrossRef]
- Kuang, C.; Jing, S.X.; Liu, Y.; Luo, S.H.; Li, S.H. Drimane sesquiterpenoids and isochromone derivative from the endophytic fungus Pestalotiopsis sp. M-23. Nat. Prod. Bioprospect. 2016, 6, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Beattie, K.D.; Ellwood, N.; Kumar, R.; Yang, X.; Healy, P.C.; Choomuenwai, V.; Quinn, R.J.; Elliott, A.G.; Huang, J.X.; Chitty, J.L.; et al. Antibacterial and antifungal screening of natural products sourced from Australian fungi and characterisation of pestalactams D–F. Phytochemistry 2016, 124, 79–85. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, S.; Wang, B.; Niu, S.; Wu, W.; Guo, L.; Che, Y. Four new tetramic acid and one new furanone derivatives from the plant endophytic fungus Neopestalotiopsis sp. Fitoterapia 2015, 103, 106–112. [Google Scholar] [CrossRef]
- Arora, P.; Wani, Z.A.; Nalli, Y.; Ali, A.; Riyaz-Ul-Hassan, S. Antimicrobial potential of thiodiketopiperazine derivatives produced by Phoma sp., an endophyte of Glycyrrhiza glabra Linn. Microb. Ecol. 2016, 72, 802–812. [Google Scholar] [CrossRef]
- Xia, X.; Kim, S.; Bang, S.; Lee, H.J.; Liu, C.; Park, C.I.; Shim, S.H. Barceloneic acid C, a new polyketide from an endophytic fungus Phoma sp. JS752 and its antibacterial activities. J. Antibiot. 2015, 68, 139–141. [Google Scholar] [CrossRef]
- De Medeiros, L.S.; Abreu, L.M.; Nielsen, A.; Ingmer, H.; Larsen, T.O.; Nielsen, K.F.; Rodrigues-Filho, E. Dereplication-guided isolation of depsides thielavins S–T and lecanorins D–F from the endophytic fungus Setophoma sp. Phytochemistry 2015, 111, 154–162. [Google Scholar] [CrossRef]
- Li, Y.; Wei, W.; Wang, R.L.; Liu, F.; Wang, Y.K.; Li, R.; Khan, B.; Lin, J.; Yan, W.; Ye, Y.H. Colletolides A and B, two new γ-butyrolactone derivatives from the endophytic fungus Colletotrichum gloeosporioides. Phytochem. Lett. 2019, 33, 90–93. [Google Scholar] [CrossRef]
- Wang, W.X.; Kusari, S.; Laatsch, H.; Golz, C.; Kusari, P.; Strohmann, C.; Kayser, O.; Spiteller, M. Antibacterial azaphilones from an endophytic fungus, Colletotrichum sp. BS4. J. Nat. Prod. 2016, 79, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Yin, R.; Zhou, Z.; Gu, G.; Dai, J.; Lai, D.; Zhou, L. Eremophilane-type sesquiterpenoids from the endophytic fungus Rhizopycnis vagum and their antibacterial, cytotoxic, and phytotoxic activities. Front. Chem. 2020, 8, 980. [Google Scholar] [CrossRef]
- Wang, A.; Li, P.; Zhang, X.; Han, P.; Lai, D.; Zhou, L. Two new anisic acid derivatives from endophytic fungus Rhizopycnis vagum Nitaf22 and their antibacterial activity. Molecules 2018, 23, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, D.; Wang, A.; Cao, Y.; Zhou, K.; Mao, Z.; Dong, X.; Tian, J.; Xu, D.; Dai, J.; Peng, Y.; et al. Bioactive dibenzo-α-pyrone derivatives from the endophytic fungus Rhizopycnis vagum Nitaf22. J. Nat. Prod. 2016, 79, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Liu, T.K.; Shi, Q.; Yang, X.L. Sesquiterpenoids and diterpenes with antimicrobial activity from Leptosphaeria sp. XL026, an endophytic fungus in Panax notoginseng. Fitoterapia 2019, 137, 104243. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W.; Wu, C.; Feng, H.; Peng, Y.; Shahid, H.; Cui, Z.; Ding, P.; Shan, T. Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta. BMC Microbiol. 2021, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mou, Q.L.; Yang, S.X.; Xiang, T.; Liu, W.W.; Yang, J.; Guo, L.P.; Wang, W.J.; Yang, X.L. New cytochalasan alkaloids and cyclobutane dimer from an endophytic fungus Cytospora chrysosperma in Hippophae rhamnoides and their antimicrobial activities. Tetrahedron Lett. 2021, 87, 153207. [Google Scholar] [CrossRef]
- Mao, Z.; Xue, M.; Gu, G.; Wang, W.; Li, D.; Lai, D.; Zhou, L. Lophiostomin A–D: New 3, 4-dihydroisocoumarin derivatives from the endophytic fungus Lophiostoma sp. Sigrf10. RSC Adv. 2020, 10, 6985–6991. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harrasi, A.; Ahmad, M.; Mabood, F.; Hassan, Z.; Shah, A.; Green, I.R.; Schulz, B.; et al. Microsphaerol and seimatorone: Two new compounds isolated from the endophytic fungi, Microsphaeropsis sp. and Seimatosporium sp. Chem. Biodivers. 2015, 12, 289–294. [Google Scholar] [CrossRef]
- Harwoko, H.; Lee, J.; Hartmann, R.; Mándi, A.; Kurtán, T.; Müller, W.E.; Feldbrügge, M.; Kalscheuer, R.; Ancheeva, E.; Daletos, G.; et al. Azacoccones FH, new flavipin-derived alkaloids from an endophytic fungus Epicoccum nigrum MK214079. Fitoterapia 2020, 146, 104698. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; Melong, R.; Tsamo, A.T.; Maffo, T.; Kapche, D.G.; Ngadjui, B.T.; McGaw, L.J.; Eloff, J.N. Cytotoxicity, antioxidant and antibacterial activity of four compounds produced by an endophytic fungus Epicoccum nigrum associated with Entada abyssinica. Rev. Bras. Farmacogn. 2017, 27, 251–253. [Google Scholar] [CrossRef]
- Xu, Z.L.; Zheng, N.; Cao, S.M.; Li, S.T.; Mo, T.X.; Qin, Y.Y.; Li, J.; Yang, R.Y. Secondary metabolites from the endophytic fungus Stemphylium lycopersici and their antibacterial activities. Chem. Nat. Compd. 2020, 56, 1162–1165. [Google Scholar] [CrossRef]
- Liu, Y.; Marmann, A.; Abdel-Aziz, M.S.; Wang, C.Y.; Müller, W.E.; Lin, W.H.; Mándi, A.; Kurtán, T.; Daletos, G.; Proksch, P. Tetrahydroanthraquinone derivatives from the endophytic fungus Stemphylium globuliferum. Eur. J. Org. Chem. 2015, 2015, 2646–2653. [Google Scholar] [CrossRef]
- Mai, P.Y.; Levasseur, M.; Buisson, D.; Touboul, D.; Eparvier, V. Identification of antimicrobial compounds from Sandwithia guyanensis-associated endophyte using molecular network approach. Plants 2020, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Ramesha, K.P.; Mohana, N.C.; Nuthan, B.R.; Rakshith, D.; Satish, S. Antimicrobial metabolite profiling of Nigrospora sphaerica from Adiantum philippense L. J. Genet. Eng. Biotechnol. 2020, 18, 1–9. [Google Scholar] [CrossRef]
- Kornsakulkarn, J.; Choowong, W.; Rachtawee, P.; Boonyuen, N.; Kongthong, S.; Isaka, M.; Thongpanchang, C. Bioactive hydroanthraquinones from endophytic fungus Nigrospora sp. BCC 47789. Phytochem. Lett. 2018, 24, 46–50. [Google Scholar] [CrossRef]
- Kaaniche, F.; Hamed, A.; Abdel-Razek, A.S.; Wibberg, D.; Abdissa, N.; El Euch, I.Z.; Allouche, N.; Mellouli, L.; Shaaban, M.; Sewald, N. Bioactive secondary metabolites from new endophytic fungus Curvularia sp. isolated from Rauwolfia macrophylla. PLoS ONE 2019, 14, e0217627. [Google Scholar] [CrossRef] [Green Version]
- Hilario, F.; Polinário, G.; de Amorim, M.R.; de Sousa Batista, V.; do Nascimento Júnior, N.M.; Araújo, A.R.; Bauab, T.M.; Dos Santos, L.C. Spirocyclic lactams and curvulinic acid derivatives from the endophytic fungus Curvularia lunata and their antibacterial and antifungal activities. Fitoterapia 2020, 141, 104466. [Google Scholar] [CrossRef]
- Long, Y.; Tang, T.; Wang, L.Y.; He, B.; Gao, K. Absolute configuration and biological activities of meroterpenoids from an endophytic fungus of Lycium barbarum. J. Nat. Prod. 2019, 82, 2229–2237. [Google Scholar] [CrossRef]
- He, J.; Li, Z.H.; Ai, H.L.; Feng, T.; Liu, J.K. Anti-bacterial chromones from cultures of the endophytic fungus Bipolaris eleusines. Nat. Prod. Res. 2019, 33, 3515–3520. [Google Scholar] [CrossRef]
- Yang, Y.H.; Yang, D.S.; Li, G.H.; Pu, X.J.; Mo, M.H.; Zhao, P.J. Antibacterial diketopiperazines from an endophytic fungus Bionectria sp. Y1085. J. Antibiot. 2019, 72, 752–758. [Google Scholar] [CrossRef]
- Kamdem, R.S.; Pascal, W.; Rehberg, N.; van Geelen, L.; Höfert, S.P.; Knedel, T.O.; Janiak, C.; Sureechatchaiyan, P.; Kassack, M.U.; Lin, W.; et al. Metabolites from the endophytic fungus Cylindrocarpon sp. isolated from tropical plant Sapium ellipticum. Fitoterapia 2018, 128, 175–179. [Google Scholar] [CrossRef]
- Li, G.; Kusari, S.; Golz, C.; Laatsch, H.; Strohmann, C.; Spiteller, M. Epigenetic modulation of endophytic Eupenicillium sp. LG41 by a histone deacetylase inhibitor for production of decalin-containing compounds. J. Nat. Prod. 2017, 80, 983–988. [Google Scholar] [CrossRef]
- Teponno, R.B.; Noumeur, S.R.; Helaly, S.E.; Hüttel, S.; Harzallah, D.; Stadler, M. Furanones and anthranilic acid derivatives from the endophytic fungus Dendrothyrium variisporum. Molecules 2017, 22, 1674. [Google Scholar] [CrossRef] [Green Version]
- Pina, J.R.S.; Silva-Silva, J.V.; Carvalho, J.M.; Bitencourt, H.R.; Watanabe, L.A.; Fernandes, J.M.P.; Souza, G.E.D.; Aguiar, A.C.C.; Guido, R.V.C.; Almeida-Souza, F.; et al. Antiprotozoal and antibacterial activity of ravenelin, a xanthone isolated from the endophytic fungus Exserohilum rostratum. Molecules 2021, 26, 3339. [Google Scholar] [CrossRef]
- Pinheiro, E.A.; Borges, F.C.; Pina, J.R.; Ferreira, L.R.; Cordeiro, J.S.; Carvalho, J.M.; Feitosa, A.O.; Campos, F.R.; Barison, A.; Souza, A.D.; et al. Annularins I and J: New metabolites isolated from endophytic fungus Exserohilum rostratum. J. Braz. Chem. Soc. 2016, 27, 1432–1436. [Google Scholar]
- Pan, Y.; Zheng, W.; Yang, S. Chemical and activity investigation on metabolites produced by an endophytic fungi Psathyrella candolleana from the seed of Ginkgo biloba. Nat. Prod. Res. 2020, 34, 3130–3133. [Google Scholar] [CrossRef]
- Duan, X.X.; Qin, D.; Song, H.C.; Gao, T.C.; Zuo, S.H.; Yan, X.; Wang, J.Q.; Ding, X.; Di, Y.T.; Dong, J.Y. Irpexlacte A-D, four new bioactive metabolites of endophytic fungus Irpex lacteus DR10-1 from the waterlogging tolerant plant Distylium chinense. Phytochem. Lett. 2019, 32, 151–156. [Google Scholar] [CrossRef]
- Rehberg, N.; Akone, H.S.; Ioerger, T.R.; Erlenkamp, G.; Daletos, G.; Gohlke, H.; Proksch, P.; Kalscheuer, R. Chlorflavonin targets acetohydroxyacid synthase catalytic subunit IlvB1 for synergistic killing of Mycobacterium tuberculosis. ACS Infect. Dis. 2018, 4, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef] [PubMed]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Guo, Y.; Jud, W.; Weikl, F.; Ghirardo, A.; Junker, R.R.; Polle, A.; Benz, J.P.; Pritsch, K.; Schnitzler, J.P.; Rosenkranz, M. Volatile organic compound patterns predict fungal trophic mode and lifestyle. Commun. Biol. 2021, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Weisskopf, L.; Schulz, S.; Garbeva, P. Microbial volatile organic compounds in intra-kingdom and inter-kingdom interactions. Nat. Rev. Microbiol. 2021, 19, 391–404. [Google Scholar] [CrossRef]
- Chen, J.J.; Feng, X.; Xia, C.Y.; Kong, D.; Qi, Z.Y.; Liu, F.; Chen, D.; Lin, F.; Zhang, C. Confirming the phylogenetic position of the genus Muscodor and the description of a new Muscodor species. Mycosphere 2019, 10, 187–201. [Google Scholar] [CrossRef]
- Saxena, S.; Strobel, G.A. Marvellous Muscodor spp.: Update on Their Biology and Applications. Microb. Ecol. 2020, 82, 5–20. [Google Scholar] [CrossRef]
- Ezra, D.; Hess, W.; Strobel, G. Unique wild type endophytic isolates of Muscodor albus, a volatile antibiotic producing fungus. Microbiology 2004, 150, 4023–4031. [Google Scholar] [CrossRef] [Green Version]
- Atmosukarto, I.; Castillo, U.; Hess, W.M.; Sears, J.; Strobel, G. Isolation and characterization of M. albus I-41.3 s, a volatile antibiotic producing fungus. Plant Sci. 2010, 169, 854–861. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Strobel, G.A.; Moore, E.; Robison, R.; Sears, J. Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 2010, 156, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.L.; Wang, G.P.; Mao, L.J.; Komon-Zelazowska, M.; Yuan, Z.L.; Lin, F.C.; Druzhinina, I.S.; Kubicek, C.P. Muscodor fengyangensis sp. nov. from southeast China: Morphology, physiology and production of volatile compounds. Fungal Biol. 2010, 114, 797–808. [Google Scholar] [CrossRef]
- Meshram, V.; Kapoor, N.; Saxena, S. Muscodor kashayum sp. nov.–a new volatile anti-microbial producing endophytic fungus. Mycology 2013, 4, 196–204. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Bussaban, B.; Hyde, K.D.; Matsui, K.; Lumyong, S. Molecular and morphological evidence support four new species in the genus Muscodor from northern Thailand. Ann. Microbiol. 2013, 63, 1341–1351. [Google Scholar] [CrossRef]
- Strobel, G.A.; Blatt, B. Volatile Organic Compound Formulations Having Antimicrobial Activity. U.S. Patent Application No. 16/179,370, 5 September 2019. [Google Scholar]
- Suwannarach, N.; Kaewyana, C.; Yodmeeklin, A.; Kumla, J.; Matsui, K.; Lumyong, S. Evaluation of Muscodor cinnamomi as an egg biofumigant for the reduction of microorganisms on the eggshell surface and its effect on egg quality. Int. J. Food Microbiol. 2017, 244, 52–61. [Google Scholar] [CrossRef]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium- chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Levison, M. Effect of colon flora and short chain fatty acids on in vitro growth of Pseudomonas aeruginosa and Enterobacteriaceae. Infect. Immun. 1973, 8, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Moo, C.L.; Yang, S.K.; Osman, M.A.; Yuswam, M.H.; Loh, J.Y.; Lim, W.M.; Lim, S.H.E.; Lai, K.S. Antibacterial activity and mode of action of β- caryophyllene on Bacillus cereus. Pol. J. Microbiol. 2020, 68, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Bakun, P.; Czarczynska-Goslinka, B.; Goslinka, T.; Lijewiski, S. In vitro and in vivo biological activities of azulene derivatives with potential applications in medicine. Med. Chem. Res. 2021, 30, 834–846. [Google Scholar] [CrossRef]
- Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic mapping of fungal secondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef] [Green Version]
- Wasil, Z.; Pahirulzaman, K.A.K.; Butts, C.; Simpson, T.J.; Lazarus, C.M.; Cox, R.J. One pathway, many compounds: Heterologous expression of a fungal biosynthetic pathway reveals its intrinsic potential for diversity. Chem. Sci. 2013, 4, 3845–3856. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef]
- Bharatiya, P.; Rathod, P.; Hiray, A.; Kate, A.S. Multifarious elicitors: Invoking biosynthesis of various bioactive secondary metabolite in fungi. Appl. Biochem. Biotechnol. 2021, 193, 668–686. [Google Scholar] [CrossRef]
- Schneider, P.; Misiek, M.; Hoffmeister, D. In vivo and in vitro production o ptions for fungal secondary metabolites. Mol. Pharm. 2008, 5, 234–242. [Google Scholar] [CrossRef]
- Yu, M.; Li, Y.; Banakar, S.P.; Liu, L.; Shao, C.; Li, Z.; Wang, C. New metabolites from the co-culture of marine derived actinomycete Streptomyces rochei MB037 and fungus Rhinocladiella similis 35. Front. Microbiol. 2019, 10, 915. [Google Scholar] [CrossRef] [Green Version]
- Huo, L.; Hug, J.J.; Fu, C.; Bian, X.; Zhang, Y.; Müller, R. Heterologous expression of bacterial natural product biosynthetic pathways. Nat. Prod. Rep. 2019, 36, 1412–1436. [Google Scholar] [CrossRef]
- Zhang, W.; Shao, C.L.; Chen, M.; Liu, Q.A.; Wang, C.Y. Brominated resorcylic acid lactones from the marine-derived fungus Cochliobolus lunatus induced by histone deacetylase inhibitors. Tetrahedron Lett. 2014, 55, 4888–4891. [Google Scholar] [CrossRef]
- Wang, Z.R.; Li, G.; Ji, L.X.; Wang, H.H.; Gao, H.; Peng, X.P.; Lou, H.X. Induced production of steroids by co-cultivation of two endophytes from Mahonia fortunei. Steroids 2019, 145, 1–4. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; Shao, C.L.; Liu, Y.; Zhao, D.L.; Cao, F.; Fu, X.M.; Yu, J.Y.; Wu, J.S.; Zhang, Z.K.; Wang, C.Y. Terpenoids from the coral-derived fungus Trichoderma harzianum (XS-20090075) induced by chemical epigenetic manipulation. Front. Microbiol. 2020, 11, 572. [Google Scholar] [CrossRef]
- Feng, X.; He, C.; Jiao, L.; Liang, X.; Zhao, R.; Guo, Y. Analysis of differential expression proteins reveals the key pathway in response to heat stress in Alicyclobacillus acidoterrestris DSM 3922T. Food Microbiol. 2019, 80, 77–84. [Google Scholar] [CrossRef]
- Yushchuk, O.; Ostash, I.; Mösker, E.; Vlasiuk, I.; Deneka, M.; Rückert, C.; Busche, T.; Fedorenko, V.; Kalinowski, J.; Süssmuth, R.D.; et al. Eliciting the silent lucensomycin biosynthetic pathway in Streptomyces cyanogenus S136 via manipulation of the global regulatory gene adpA. Sci. Rep. 2021, 11, 3507. [Google Scholar] [CrossRef]
- Libis, V.; Antonovsky, N.; Zhang, M.; Shang, Z.; Montiel, D.; Maniko, J.; Ternei, M.A.; Calle, P.Y.; Lemetre, C.; Owen, J.G.; et al. Uncovering the biosynthetic potential of rare metagenomic DNA using co-occurrence network analysis of targeted sequences. Nat. Commun. 2019, 10, 3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, F.; Leng, D.J.; Wilkening, I.; Song, L.; Tosin, M.; Corre, C. Triggering the expression of a silent gene cluster from genetically intractable bacteria results in scleric acid discovery. Chem. Sci. 2019, 10, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, W.; Chen, L.; Zhao, C.; Wu, J.; Yan, D.; Deng, Z.; Sun, Y. In vitro packaging mediated one-step targeted cloning of natural product pathway. ACS Synth. Biol. 2019, 8, 1991–1997. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Huang, C.; Luo, Y. Recent Advances in Silent Gene Cluster Activation in Streptomyces. Front. Bioeng. Biotechnol. 2021, 9, 632230. [Google Scholar] [CrossRef]
- Li, L.; Wei, K.; Zheng, G.; Liu, X.; Chen, S.; Jiang, W.; Lu, Y. CRISPR-Cpf1-assisted multiplex genome editing and transcriptional repression in Streptomyces. Appl. Environ. Microbiol. 2018, 84, e00827-18. [Google Scholar] [CrossRef] [Green Version]
- Poças-Fonseca, M.J.; Cabral, C.G.; Manfrão-Netto, J.H.C. Epigenetic manipulation of filamentous fungi for biotechnological applications: A systematic review. Biotechnol. Lett. 2020, 42, 885–904. [Google Scholar] [CrossRef]
- Mao, X.M.; Xu, W.; Li, D.; Yin, W.B.; Chooi, Y.H.; Li, Y.Q.; Tang, Y.; Hu, Y. Epigenetic genome mining of an endophytic fungus leads to the pleiotropic bio- synthesis of natural products. Angew. Chem. Int. Ed. 2015, 54, 7592–7596. [Google Scholar] [CrossRef] [Green Version]
- Strauss, J.; Reyes-Dominguez, Y. Regulation of secondary metabolism by chromatin structure and epigenetic codes. Fungal Genet. Biol. 2011, 48, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Gacek, A.; Strauss, J. The chromatin code of fungal secondary metabolite gene clusters. Appl. Microbiol. Biotechnol. 2012, 95, 1389–1404. [Google Scholar] [CrossRef] [Green Version]
- Aghcheh, R.K.; Kubicek, C.P. Epigenetics as an emerging tool for improvement of fungal strains used in biotechnology. Appl. Microbiol. Biotechnol. 2015, 99, 6167–6181. [Google Scholar] [CrossRef]
- Li, C.Y.; Chung, Y.M.; Wu, Y.C.; Hunyadi, A.; Wang, C.C.; Chang, F.R. Natural products development under epigenetic modulation in fungi. Phytochem. Rev. 2020, 19, 1323–1340. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, N.; Hwang, S.; Kim, W.; Lee, Y.; Cho, S.; Palsson, B.O.; Cho, B.K. Discovery of novel secondary metabolites encoded in actinomycete genomes through coculture. J. Ind. Microbiol. Biotechnol. 2021, 48, kuaa001. [Google Scholar] [CrossRef]
- Tomm, H.A.; Ucciferri, L.; Ross, A.C. Advances in microbial culturing conditions to activate silent biosynthetic gene clusters for novel metabolite production. J. Ind. Microbiol. Biotechnol. 2019, 46, 1381–1400. [Google Scholar] [CrossRef]
- Gonciarz, J.; Bizukojc, M. Adding talc microparticles to Aspergillus terreus ATCC 20542 preculture decreases fungal pellet size and improves lovastatin production. Eng. Life Sci. 2014, 14, 190–200. [Google Scholar] [CrossRef]
- Timmermans, M.L.; Picott, K.J.; Ucciferri, L.; Ross, A.C. Culturing marine bacteria from the genus Pseudoalteromonas on a cotton scaffold alters secondary metabolite production. Microbiologyopen 2019, 8, e00724. [Google Scholar] [CrossRef] [Green Version]
- Boruta, T.; Bizukojc, M. Application of aluminum oxide nanoparticles in Aspergillus terreus cultivations: Evaluating the effects on lovastatin production and fungal morphology. Biomed. Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bode, H.B.; Bethe, B.; Hofs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochemistry 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Discovery of aspoquinolones A–D, prenylated quinoline-2-one alkaloids from Aspergillus nidulans, motivated by genome mining. Org. Biomol. Chem. 2006, 4, 3517–3520. [Google Scholar] [CrossRef]
- Scherlach, K.; Schuemann, J.; Dahse, H.M.; Hertweck, C. Aspernidine A and B, prenylated isoindolinone alkaloids from the model fungus Aspergillus nidulans. J. Antibiot. 2010, 63, 375–377. [Google Scholar] [CrossRef]
























Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deshmukh, S.K.; Dufossé, L.; Chhipa, H.; Saxena, S.; Mahajan, G.B.; Gupta, M.K. Fungal Endophytes: A Potential Source of Antibacterial Compounds. J. Fungi 2022, 8, 164. https://doi.org/10.3390/jof8020164
Deshmukh SK, Dufossé L, Chhipa H, Saxena S, Mahajan GB, Gupta MK. Fungal Endophytes: A Potential Source of Antibacterial Compounds. Journal of Fungi. 2022; 8(2):164. https://doi.org/10.3390/jof8020164
Chicago/Turabian StyleDeshmukh, Sunil K., Laurent Dufossé, Hemraj Chhipa, Sanjai Saxena, Girish B. Mahajan, and Manish Kumar Gupta. 2022. "Fungal Endophytes: A Potential Source of Antibacterial Compounds" Journal of Fungi 8, no. 2: 164. https://doi.org/10.3390/jof8020164
APA StyleDeshmukh, S. K., Dufossé, L., Chhipa, H., Saxena, S., Mahajan, G. B., & Gupta, M. K. (2022). Fungal Endophytes: A Potential Source of Antibacterial Compounds. Journal of Fungi, 8(2), 164. https://doi.org/10.3390/jof8020164