
Transparent Exopolymer Particles in Deep Oceans: Synthesis and Future Challenges


Abstract
:1. Introduction
2. Overview of the Data on TEP Distribution in Deep Waters
2.1. Data Obtained by the Colorimetry
2.1.1. Data Collected in Coastal, Slope Region, and Marginal Seas
2.1.2. Data Collected in Open Oceans
2.2. Data Obtained by the Microscopy
2.3. Summary of the Observed Data
- In the mesopelagic layer, the variability range of the TEP concentrations is on the order of 100-fold (Table 1). In the marginal sea and slope region, a vertical (depth-dependent decrease [18,19]) and a lateral (offshoreward decrease [17]) gradient in the TEP distribution pattern has been documented. Microscopic observations have revealed that TEP are colonized by prokaryotes in the mesopelagic layer [17,24].
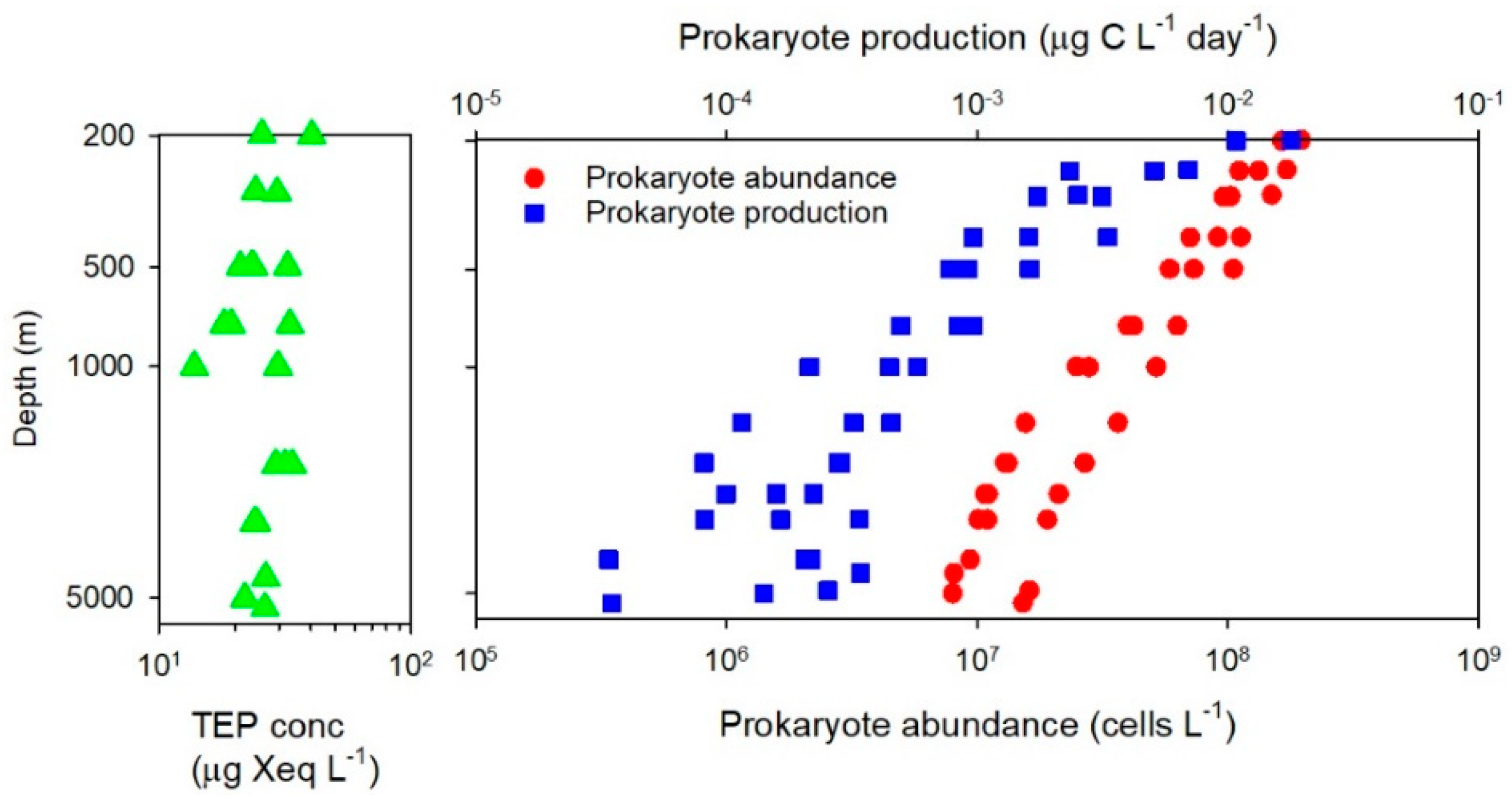
- Examination of the full-depth distribution of TEP in open oceans has revealed that TEP concentrations are less variable (<3 fold) throughout the meso- and bathypelagic water columns down to the depths of 4000–5400 m (Table 1), although some anomalous features have been noted [22,25]. This vertical distribution of TEP is largely decoupled from the distribution of prokaryote abundance and production [19] (Figure 1). One study using the microscopic method has found a remarkably high TEP abundance in the bathypelagic layer of the coastal upwelling region [25]. Microscopic observations have also found that TEP were colonized by prokaryotes in the bathypelagic layer. High relative contributions of TEP-associated prokaryotes to the total prokaryote abundance (up to 20%) were observed at the depths >2000 m in the Arctic Ocean [24].
3. Potential Factors Affecting TEP Distribution in the Deep Oceans
3.1. Sources of TEP
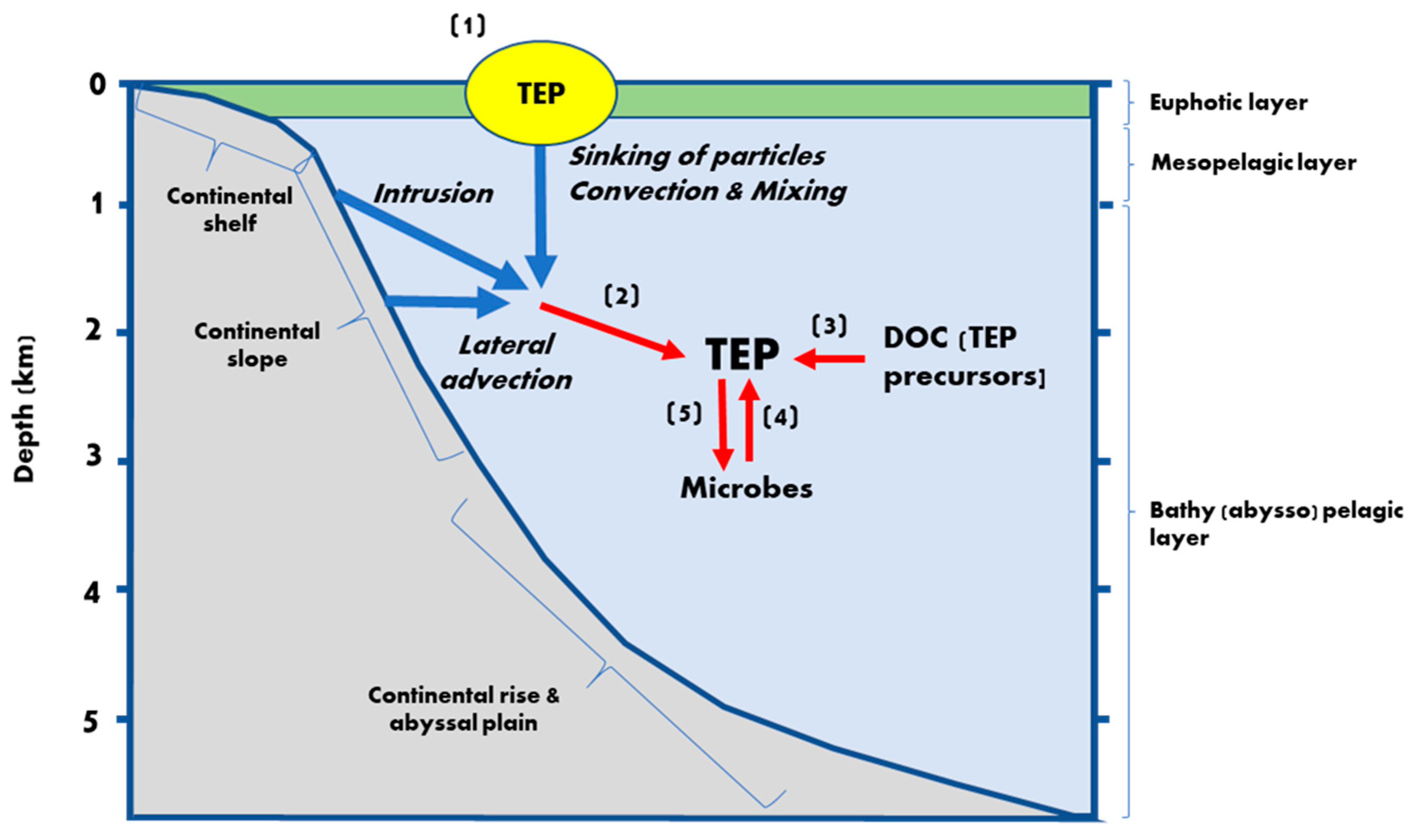
3.1.1. Transport
3.1.2. Autochthonous Production of TEP
3.2. Sinks of TEP
3.2.1. Prokaryotes
3.2.2. Grazers
4. Organic Carbon Inventory
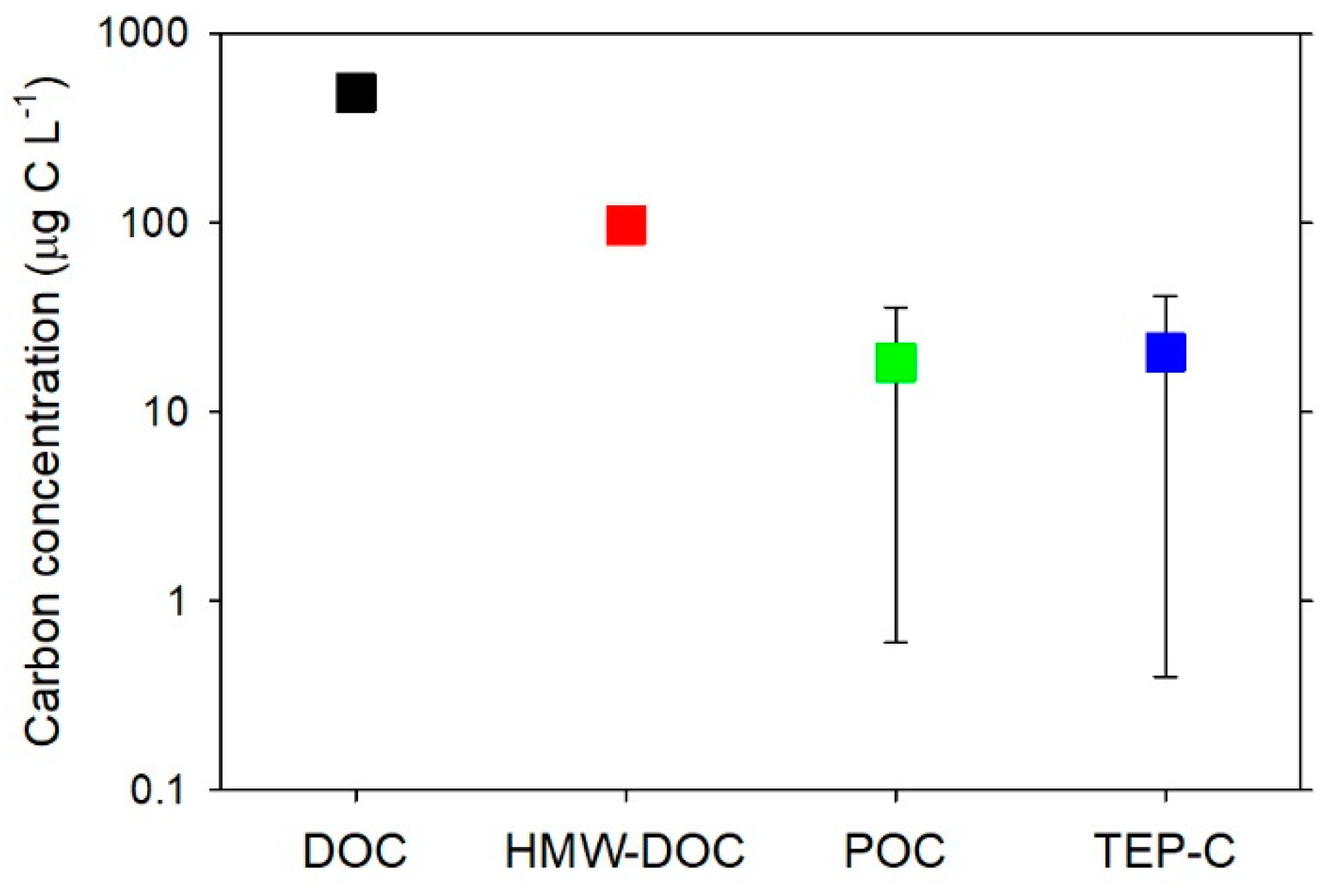
- To estimate TEP-C, studies have used a conversion factor derived from laboratory experiments using diatom cultures [15]. However, the validity of this conversion factor in deep waters has yet to be tested. If the organic carbon yield relative to the Alcian blue-reactive residues (sulfate and carboxyl groups) of TEP is systematically lower in deeper than shallower waters, the TEP-C values estimated from the conversion factor for the shallower water (diatom-derived fresh TEP) may be too high.
- TEP-C concentration may exceed POC concentration due to the use of different pore-size-filters for the determination of TEP (0.4-μm-pore-size polycarbonate filter) and POC (0.7-μm-pore-size GF/F filter). If large quantities of organic carbon associated with TEP pass through the GF/F filters, but are retained on 0.4-μm polycarbonate filters, this would explain the high TEP-C concentration relative to POC.
5. Knowledge Gaps and Future Challenges
- Theories have been proposed to explain the spontaneous assembly of gels [2,3,4] and the coagulation of particles [26] in seawater. Self-assembled gels have been identified in the deep oceanic water column [35]; however, a rigorous validation of TEP quantification methods are required to evaluate TEP formation via the spontaneous assembly of DOC. Coagulation theory is generally formulated to describe the coagulation rate as a product of particle number, collision rate, and stickiness, whereby the dominant mechanism by which the collision rate is controlled differs, depending on the particle size. Data on TEP size distributions in deep oceans are scarce [25], and we lack information about the abundance of TEP or TEP precursors in the sub-micrometer size range. Previous work has revealed that submicron particles and colloids are present in meso- and bathypelagic oceans [47,48,49], yet it remains to be seen if TEP are produced via the coagulation of submicron particles under deep water physical conditions. Disaggregation, the converse process of aggregation, may also affect TEP distribution at certain depths. Further studies are required to evaluate the extent of TEP delivery via the disaggregation of sinking particles.
- To date, only a few studies have used the microscopic method to examine prokaryote colonization on TEP at depth [17,24]. These studies have provided valuable information regarding the potential role of TEP in the food webs of deep waters. Given that deep water microbial communities are dominated by organisms with surface-associated lifestyles, as evidenced by the presence of genes encoding pilus, polysaccharide, and antibiotics synthesis [36], it is likely that TEP in deep waters represent a hot spot of microbes, including prokaryotes, protists, and viruses [13]. They can also serve as important food resources for metazoan grazers that thrive throughout the oceanic water columns [50]. Despite the extensive data collected over the past two decades concerning prokaryote, protist, and virus distributions in deep water columns [12,13], further research is needed to incorporate TEP and other gel-like particles into the food web models of deep oceans.
- To incorporate TEP dynamics into ocean carbon cycle models, it is necessary to collect quantitative data on TEP in terms of carbon. In this regard, further testing and refinement of methodologies are required to reduce large uncertainties associated with the estimation of TEP-C. It is also important to clarify the mechanisms by which TEP dynamics are regulated and to evaluate the turnover time of TEP. Currently, TEP turnover time and their lability in deep waters is poorly understood, suggesting a need to develop new methods to tackle this issue. Efforts to determine the dynamics (production and decay) of detrital polysaccharides in marine waters are inherently complicated by numerous analytical challenges [51]. We clearly need to know more about the chemical compositions, physical structures, and microbial processing of TEP and other gels in deep waters.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlson, C.A.; Hansell, D.A. DOM Sources, Sinks, Reactivity, and Budgets. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 65–126. [Google Scholar]
- Verdugo, P. Marine Microgels. Annu. Rev. Mar. Sci. 2012, 4, 375–400. [Google Scholar] [CrossRef]
- Chin, W.-C.; Orellana, M.V.; Verdugo, P. Spontaneous Assembly of Marine Dissolved Organic Matter into Polymer Gels. Nature 1998, 391, 568–572. [Google Scholar] [CrossRef]
- Verdugo, P.; Alldredge, A.L.; Azam, F.; Kirchman, D.L.; Passow, U.; Santschi, P.H. The Oceanic Gel Phase: A Bridge in the Dom–Pom Continuum. Mar. Chem. 2004, 92, 67–85. [Google Scholar] [CrossRef]
- Mari, X.; Passow, U.; Migon, C.; Burd, A.B.; Legendre, L. Transparent Exopolymer Particles: Effects on Carbon Cycling in the Ocean. Prog. Oceanogr. 2017, 151, 13–37. [Google Scholar] [CrossRef] [Green Version]
- Passow, U. Transparent Exopolymer Particles (TEP) in Aquatic Environments. Prog. Oceanogr. 2002, 55, 287–333. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; Carlson, C.A. The Biological Pump in a High CO2 World. Mar. Ecol. Prog. Ser. 2012, 470, 249–271. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; Alldredge, A.L. Distribution, Size and Bacerial Colonizaion of Transparent Exopolymer Particles in the Ocean. Mar. Ecol. Prog. Ser. 1994, 113, 185–198. [Google Scholar] [CrossRef]
- Zäncker, B.; Engel, A.; Cunliffe, M. Bacterial Communities Associated with Individual Transparent Exopolymer Particles (TEP). J. Plankton Res. 2019, 41, 561–565. [Google Scholar] [CrossRef]
- Ling, S.C.; Alldredge, A.L. Does the Marine Copepod Calanus Pacificus Consume Transparent Exopolymer Particles (TEP)? J. Plankton Res. 2003, 25, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Arístegui, J.; Gasol, J.M.; Duarte, C.M.; Herndld, G.J. Microbial Oceanography of the Dark Ocean’s Pelagic Realm. Limnol. Oceanogr. 2009, 54, 1501–1529. [Google Scholar] [CrossRef] [Green Version]
- Herndl, G.J.; Reinthaler, T. Microbial Control of the Dark End of the Biological Pump. Nat. Geosci. 2013, 6, 718–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, T.; Tamburini, C.; Arístegui, J.; Baltar, F.; Bochdansky, A.B.; Fonda-Umani, S.; Fukuda, H.; Gogou, A.; Hansell, D.A.; Hansman, R.L.; et al. Emerging Concepts on Microbial Processes in the Bathypelagic Ocean—Ecology, Biogeochemistry, and Genomics. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1519–1536. [Google Scholar] [CrossRef]
- Robinson, C.; Steinberg, D.K.; Anderson, T.R.; Arístegui, J.; Carlson, C.A.; Frost, J.R.; Ghiglione, J.-F.; Hernández-León, S.; Jackson, G.A.; Koppelmann, R.; et al. Mesopelagic Zone Ecology and Biogeochemistry—A Synthesis. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1504–1518. [Google Scholar] [CrossRef]
- Passow, U.; Alldredge, A.L. A Dye-Binding Assay for the Spectrophotometric Measurement of Transparent Exopolymer Particles (TEP). Limnol. Oceanogr. 1995, 40, 1326–1335. [Google Scholar] [CrossRef]
- Engel, A.; Passow, U. Carbon and Nitrogen Content of Transparent Exopolymer Particles (TEP) in Relation to Their Alcian Blue Adsorption. Mar. Ecol. Prog. Ser. 2001, 219, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bar-Zeev, E.; Berman, T.; Rahav, E.; Dishon, G.; Herut, B.; Berman-Frank, I. Transparent Exopolymer Particle (TEP) Dynamics in the Eastern Mediterranean Sea. Mar. Ecol. Prog. Ser. 2011, 431, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Retuerta, E.; Mazuecos, I.P.; Reche, I.; Gasol, J.M.; Álvarez-Salgado, X.A.; Álvarez, M.; Montero, M.F.; Arístegui, J. Transparent Exopolymer Particle (TEP) Distribution and in Situ Prokaryotic Generation across the Deep Mediterranean Sea and Nearby North East Atlantic Ocean. Prog. Oceanogr. 2019, 173, 180–191. [Google Scholar] [CrossRef]
- Yamada, Y.; Yokokawa, T.; Uchimiya, M.; Nishino, S.; Fukuda, H.; Ogawa, H.; Nagata, T. Transparent Exopolymer Particles (TEP) in the Deep Ocean: Full-Depth Distribution Patterns and Contribution to the Organic Carbon Pool. Mar. Ecol. Prog. Ser. 2017, 583, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Fukuda, H.; Uchimiya, M.; Motegi, C.; Nishino, S.; Kikuchi, T.; Nagata, T. Localized Accumulation and a Shelf-Basin Gradient of Particles in the Chukchi Sea and Canada Basin, Western Arctic. J. Geophys. Res. Ocean. 2015, 120, 4638–4653. [Google Scholar] [CrossRef]
- Annane, S.; St-Amand, L.; Starr, M.; Pelletier, E.; Ferreyra, G.A. Contribution of Transparent Exopolymeric Particles (TEP) to Estuarine Particulate Organic Carbon Pool. Mar. Ecol. Prog. Ser. 2015, 529, 17–34. [Google Scholar] [CrossRef]
- Cisternas-Novoa, C.; Lee, C.; Engel, A. Transparent Exopolymer Particles (TEP) and Coomassie Stainable Particles (CSP): Differences between Their Origin and Vertical Distributions in the Ocean. Mar. Chem. 2015, 175, 56–71. [Google Scholar] [CrossRef]
- Gardner, W.D.; Jo Richardson, M.; Mishonov, A.V.; Biscaye, P.E. Global Comparison of Benthic Nepheloid Layers Based on 52 Years of Nephelometer and Transmissometer Measurements. Prog. Oceanogr. 2018, 168, 100–111. [Google Scholar] [CrossRef]
- Busch, K.; Endres, S.; Iversen, M.H.; Michels, J.; Nöthig, E.-M.; Engel, A. Bacterial Colonization and Vertical Distribution of Marine Gel Particles (TEP and CSP) in the Arctic Fram Strait. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.; Endres, S.; Galgani, L.; Schartau, M. Marvelous Marine Microgels: On the Distribution and Impact of Gel-Like Particles in the Oceanic Water-Column. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Burd, A.B.; Jackson, G.A. Particle Aggregation. Ann. Rev. Mar. Sci. 2009, 1, 65–90. [Google Scholar] [CrossRef] [PubMed]
- Weinbauer, M.G.; Liu, J.; Motegi, C.; Maier, C.; Pedrotti, M.L.; Dai, M.; Gattuso, J.P. Seasonal Variability of Microbial Respiration and Bacterial and Archaeal Community Composition in the Upper Twilight Zone. Aquat. Microb. Ecol. 2013, 71, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Azam, F.; Long, R.A. Sea Snow Microcosms. Nature 2001, 414, 497–498. [Google Scholar] [CrossRef]
- Dilling, L.; Alldredge, A.L. Fragmentation of Marine Snow by Swimming Macrozooplankton: A New Process Impacting Carbon Cycling in the Sea. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2000, 47, 1227–1245. [Google Scholar] [CrossRef]
- Martin, J.H.; Knauer, G.A.; Karl, D.M.; Broenkow, W.W. Vertex—Carbon Cycling in the Northeast Pacific. Deep. Sea Res. 1987, 34, 267–285. [Google Scholar] [CrossRef]
- Karl, D.M.; Knauer, G.A.; Martin, J.H. Downward Flux of Particulate Organic Matter in the Ocean: A Particle Decomposition Paradox. Nature 1988, 332, 438–441. [Google Scholar] [CrossRef]
- Passow, U.; Shipe, R.F.; Murray, A.; Pak, D.K.; Brzezinski, M.A.; Alldredge, A.L. The Origin of Transparent Exopolymer Particles (TEP) and Their Role in the Sedimentation of Particulate Matter. Cont. Shelf Res. 2001, 21, 327–346. [Google Scholar] [CrossRef] [Green Version]
- Boyd, P.W.; Claustre, H.; Levy, M.; Siegel, D.A.; Weber, T. Multi-Faceted Particle Pumps Drive Carbon Sequestration in the Ocean. Nature 2019, 568, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Benner, R.; Amon, R.M. The Size-Reactivity Continuum of Major Bioelements in the Ocean. Ann. Rev. Mar. Sci. 2015, 7, 185–205. [Google Scholar] [CrossRef]
- Ding, Y.-X.; Chin, W.-C.; Verdugo, P. Development of a Fluorescence Quenching Assay to Measure the Fraction of Organic Carbon Present in Self-Assembled Gels in Seawater. Mar. Chem. 2007, 106, 456–462. [Google Scholar] [CrossRef]
- DeLong, E.F.; Preston, C.M.; Mincer, T.; Rich, V.; Hallam, S.J.; Frigaard, N.U.; Martinez, A.; Sullivan, M.B.; Edwards, R.; Brito, B.R.; et al. Community Genomics among Stratified Microbial Assemblages in the Ocean’s Interior. Science 2006, 311, 496–503. [Google Scholar] [CrossRef]
- Vidal-Melgosa, S.; Sichert, A.; Francis, T.B.; Bartosik, D.; Niggemann, J.; Wichels, A.; Willats, W.G.T.; Fuchs, B.M.; Teeling, H.; Becher, D.; et al. Diatom Fucan Polysaccharide Precipitates Carbon During Algal Blooms. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- Hansell, D.A. Recalcitrant Dissolved Organic Carbon Fractions. Annu. Rev. Mar. Sci. 2013, 5, 421–445. [Google Scholar] [CrossRef]
- Bercovici, S.K.; Arroyo, M.C.; De Corte, D.; Yokokawa, T.; Hansell, D.A. Limited Utilization of Extracted Dissolved Organic Matter by Prokaryotic Communities from the Subtropical North Atlantic. Limnol. Oceanogr. 2021. [Google Scholar] [CrossRef]
- Shen, Y.; Benner, R. Molecular Properties Are a Primary Control on the Microbial Utilization of Dissolved Organic Matter in the Ocean. Limnol. Oceanogr. 2019, 65, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Fukuda, H.; Fukuda, R.; Koike, I. Bacterioplankton Distribution and Production in Deep Pacific Waters: Large-Scale Geographic Variations and Possible Coupling with Sinking Particle Fluxes. Limnol. Oceanogr. 2000, 45, 426–435. [Google Scholar] [CrossRef]
- Stief, P.; Elvert, M.; Glud, R.N. Respiration by “Marine Snow” at High Hydrostatic Pressure: Insights from Continuous Oxygen Measurements in a Rotating Pressure Tank. Limnol. Oceanogr. 2021. [Google Scholar] [CrossRef]
- Tamburini, C.; Boutrif, M.; Garel, M.; Colwell, R.R.; Deming, J.W. Prokaryotic Responses to Hydrostatic Pressure in the Ocean-—A Review. Environ. Microbiol 2013, 15, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, R.; Ogawa, H.; Nagata, T.; Koike, I.I. Direct Determination of Carbon and Nitrogen Contents of Natural Bacterial Assemblages in Marine Environments. Appl. Environ. Microbiol 1998, 64, 3352–3358. [Google Scholar] [CrossRef] [Green Version]
- Tanet, L.; Martini, S.; Casalot, L.; Tamburini, C. Reviews and Syntheses: Bacterial Bioluminescence—Ecology and Impact in the Biological Carbon Pump. Biogeosciences 2020, 17, 3757–3778. [Google Scholar] [CrossRef]
- Turnewitsch, R.; Springer, B.M.; Kiriakoulakis, K.; Vilas, J.C.; Arístegui, J.; Wolff, G.; Peine, F.; Werk, S.; Graf, G.; Waniek, J.J. Determination of Particulate Organic Carbon (POC) in Seawater: The Relative Methodological Importance of Artificial Gains and Losses in Two Glass-Fiber-Filter-Based Techniques. Mar. Chem. 2007, 105, 208–228. [Google Scholar] [CrossRef]
- Koike, I.; Hara, S.; Terauchi, K.; Kogure, K. Role of Sub-Micrometer Particles in the Ocean. Nature 1990, 345, 242–244. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Koike, I. Marine Colloids: Thier Roles in Food Webs and Biogeochemical Fluxes. In Biogeochemical Processes and Ocean Flux in the Western Pacific; Sakai, H., Nozaki, Y., Eds.; Terra Scientific Publishing Company (TERRAPUB): Tokyo, Japan, 1995; pp. 275–292. [Google Scholar]
- Wells, M.L.; Goldberg, E.D. The Distribution of Colloids in the North-Atlantic and Southern Oceans. Limnol. Oceanogr. 1994, 39, 286–302. [Google Scholar] [CrossRef]
- Hernandez-Leon, S.; Koppelmann, R.; Fraile-Nuez, E.; Bode, A.; Mompean, C.; Irigoien, X.; Olivar, M.P.; Echevarria, F.; Fernandez de Puelles, M.L.; Gonzalez-Gordillo, J.I.; et al. Large Deep-Sea Zooplankton Biomass Mirrors Primary Production in the Global Ocean. Nat. Commun. 2020, 11, 6048. [Google Scholar] [CrossRef] [PubMed]
- Arnosti, C.; Wietz, M.; Brinkhoff, T.; Hehemann, J.H.; Probandt, D.; Zeugner, L.; Amann, R. The Biogeochemistry of Marine Polysaccharides: Sources, Inventories, and Bacterial Drivers of the Carbohydrate Cycle. Ann. Rev. Mar. Sci. 2021, 13, 81–108. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region | Mesopelagic | Bathypelagic | References | ||
|---|---|---|---|---|---|
| Coastal and slope region, estuary and marginal sea | |||||
| Santa Barbara Chanell (eastern Pacific) | 20 | (200–1400) | [15] | ||
| Eastern Mediterranean Sea | 200 | (300–1000) | [17] | ||
| Mediterranean Sea and Atlantic | 1.2–35 | (200–1000) | 0.6–16 | (1000–3900) | [18] |
| Western Arctic (slope region) 1 | 37–129 | (200–1000) | 39–52 | (1230–1960) | [19] |
| St Lawrence Estuary | 15–200 | (130–320) | [21] | ||
| Open oceans | |||||
| North Atlantic Ocean (subtropical) 2 | 18–33 | (200–1000) | 16–48 | (1250–4580) | [22] |
| Central Pacific (subtropical and equatorial) 1 | 12–40 | (200–1000) | 14–34 | (1000–5370) | [19] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagata, T.; Yamada, Y.; Fukuda, H. Transparent Exopolymer Particles in Deep Oceans: Synthesis and Future Challenges. Gels 2021, 7, 75. https://doi.org/10.3390/gels7030075
Nagata T, Yamada Y, Fukuda H. Transparent Exopolymer Particles in Deep Oceans: Synthesis and Future Challenges. Gels. 2021; 7(3):75. https://doi.org/10.3390/gels7030075
Chicago/Turabian StyleNagata, Toshi, Yosuke Yamada, and Hideki Fukuda. 2021. "Transparent Exopolymer Particles in Deep Oceans: Synthesis and Future Challenges" Gels 7, no. 3: 75. https://doi.org/10.3390/gels7030075
APA StyleNagata, T., Yamada, Y., & Fukuda, H. (2021). Transparent Exopolymer Particles in Deep Oceans: Synthesis and Future Challenges. Gels, 7(3), 75. https://doi.org/10.3390/gels7030075