Suction Flows Generated by the Carnivorous Bladderwort Utricularia—Comparing Experiments with Mechanical and Mathematical Models


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Flow Data from Biological Suction Feeders
2.1.1. Material
2.1.2. Flow Visualization Using Bladderwort Traps
2.2. Flow Data From Mechanical Suction Feeders
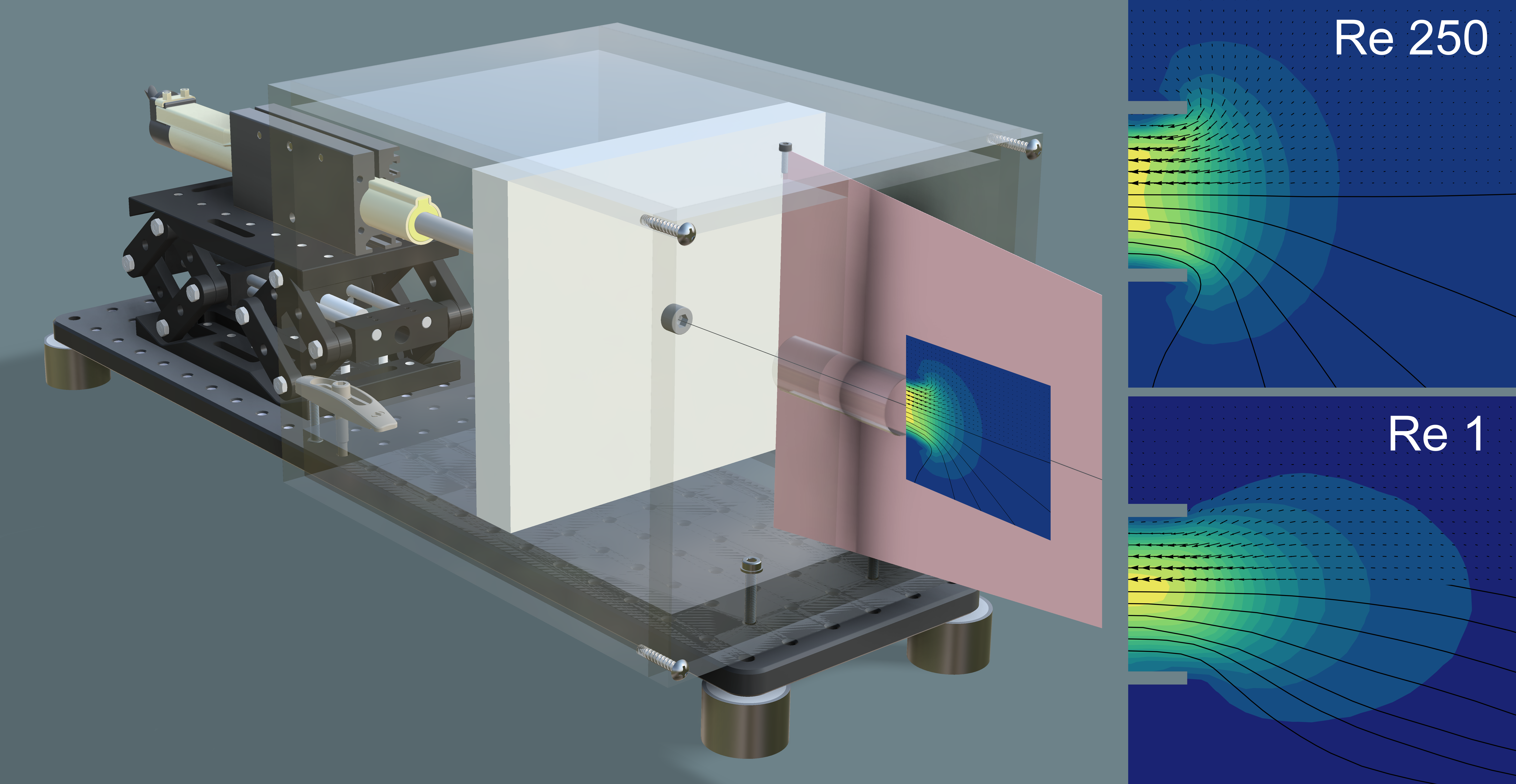
2.2.1. Designing a Dynamically Scaled Model of a Suction Feeder
2.2.2. Flow Visualization Using a Dynamically Scaled Model
3. Results
3.1. Suction Flows of Bladderwort Traps
3.2. Suction Flows of a Dynamically Scaled Model
4. Discussion
4.1. Comparing Biological Suction Flows With Theoretical Models
4.1.1. Inviscid-Flow Model
4.1.2. Creeping-Flow Model
4.1.3. Comparing Theoretical Predictions and Experimental Data
4.2. Comparing Suction Flows Across Re
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recording 1 | Gape (mm) | Flow Speed at Half Gape (m/s) | Reynolds Number 2 |
|---|---|---|---|
| 1 | 0.37 | 0.080 | 29 |
| 2 | 0.35 | 0.063 | 22 |
| 3 | 0.32 | 0.108 | 34 |
| 4 | 0.41 | 0.098 | 41 |
| 5 | 0.27 | 0.158 | 43 |
| 6 | 0.28 | 0.116 | 32 |
| 7 | 0.41 | 0.076 | 31 |
| 8 | 0.29 | 0.075 | 22 |
| 9 | 0.31 | 0.095 | 30 |
| 10 | 0.33 | 0.108 | 36 |
| 11 | 0.32 | 0.042 | 14 |
| 12 | 0.36 | 0.042 | 15 |
| 13 | 0.37 | 0.064 | 23 |
| 14 | 0.42 | 0.060 | 25 |
| 15 | 0.33 | 0.114 | 38 |
Appendix B. Velocity Field of Sampson Flow
Appendix B.1. Axial Transect
Appendix B.2. Velocity Field

References
- Wainwright, P.C.; McGee, M.D.; Longo, S.J.; Patricia Hernandez, L. Origins, innovations, and diversification of suction feeding in vertebrates. Integr. Comp. Biol. 2015, 55, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Longo, S.J.; McGee, M.D.; Oufiero, C.E.; Waltzek, T.B.; Wainwright, P.C. Body ram, not suction, is the primary axis of suction-feeding diversity in spiny-rayed fishes. J. Exp. Biol. 2016, 219, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westneat, M.W. Skull biomechanics and suction feeding in fishes. Fish Physiol. 2005, 23, 29–75. [Google Scholar] [CrossRef]
- Wainwright, P.C.; Ferry-Graham, L.A.; Waltzek, T.B.; Carroll, A.M.; Hulsey, C.D.; Grubich, J.R. Evaluating the use of ram and suction during prey capture by cichlid fishes. J. Exp. Biol. 2001, 204, 3039–3051. [Google Scholar] [PubMed]
- Holzman, R.; Day, S.W.; Mehta, R.S.; Wainwright, P.C. Jaw protrusion enhances forces exerted on prey by suction feeding fishes. J. R. Soc. Interface 2008, 5, 1445–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferry-Graham, L.A.; Wainwright, P.C.; Lauder, G.V. Quantification of flow during suction feeding in bluegill sunfish. Zoology 2003, 106, 159–168. [Google Scholar] [CrossRef]
- Holzman, R.; Day, S.W.; Mehta, R.S.; Wainwright, P.C. Integrating the determinants of suction feeding performance in centrarchid fishes. J. Exp. Biol. 2008, 211, 3296–3305. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, P.; Carroll, A.M.; Collar, D.C.; Day, S.W.; Higham, T.E.; Holzman, R.A. Suction feeding mechanics, performance, and diversity in fishes. Integr. Comp. Biol. 2007, 47, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Deban, S.M.; Olson, W.M. Biomechanics: Suction feeding by a tiny predatory tadpole. Nature 2002, 420, 41. [Google Scholar] [CrossRef]
- Stinson, C.M.; Deban, S.M. Functional trade-offs in the aquatic feeding performance of salamanders. Zoology 2017, 125, 69–78. [Google Scholar] [CrossRef]
- Marshall, C.D.; Kovacs, K.M.; Lydersen, C. Feeding kinematics, suction and hydraulic jetting capabilities in bearded seals (Erignathus barbatus). J. Exp. Biol. 2008, 211, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motta, P.J.; Hueter, R.E.; Tricas, T.C.; Summers, A.P.; Huber, D.R.; Lowry, D.; Mara, K.R.; Matott, M.P.; Whitenack, L.B.; Wintzer, A.P. Functional morphology of the feeding apparatus, feeding constraints, and suction performance in the nurse shark Ginglymostoma cirratum. J. Morphol. 2008, 269, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Gordon, E.; Pacheco, S. Prey composition in the carnivorous plants Utricularia inflata and U. gibba (Lentibulariaceae) from Paria Peninsula, Venezuela. Rev. De Biol. Trop. 2007, 55, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, S. Prey selection in three species of the carnivorous aquatic plant Utricularia (bladderwort). Arch. Für Hydrobiol. 1999, 146, 449–470. [Google Scholar] [CrossRef]
- Day, S.W.; Higham, T.E.; Cheer, A.Y.; Wainwright, P.C. Spatial and temporal patterns of water flow generated by suction-feeding bluegill sunfish Lepomis macrochirus resolved by particle image velocimetry. J. Exp. Biol. 2005, 208, 2661–2671. [Google Scholar] [CrossRef] [Green Version]
- Leeuwen, J.v.; Muller, M. Optimum sucking techniques for predatory fish. J. Zool. 1984, 37, 137–169. [Google Scholar] [CrossRef]
- Carroll, A.M.; Wainwright, P.C.; Huskey, S.H.; Collar, D.C.; Turingan, R.G. Morphology predicts suction feeding performance in centrarchid fishes. J. Exp. Biol. 2004, 207, 3873–3881. [Google Scholar] [CrossRef] [Green Version]
- Higham, T.E.; Day, S.W.; Wainwright, P.C. The pressures of suction feeding: The relation between buccal pressure and induced fluid speed in centrarchid fishes. J. Exp. Biol. 2006, 209, 3281–3287. [Google Scholar] [CrossRef] [Green Version]
- Van Wassenbergh, S.; Aerts, P.; Herrel, A. Scaling of suction-feeding kinematics and dynamics in the African catfish, Clarias gariepinus. J. Exp. Biol. 2005, 208, 2103–2114. [Google Scholar] [CrossRef] [Green Version]
- Norton, S.F.; Brainerd, E.L. Convergence in the feeding mechanics of ecomorphologically similar species in the Centrarchidae and Cichlidae. J. Exp. Biol. 1993, 176, 11–29. [Google Scholar]
- Higham, T.E.; Day, S.W.; Wainwright, P.C. Multidimensional analysis of suction feeding performance in fishes: Fluid speed, acceleration, strike accuracy and the ingested volume of water. J. Exp. Biol. 2006, 209, 2713–2725. [Google Scholar] [CrossRef] [Green Version]
- Roos, G.; Van Wassenbergh, S.; Herrel, A.; Aerts, P. Kinematics of suction feeding in the seahorse Hippocampus reidi. J. Exp. Biol. 2009, 212, 3490–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzman, R.; Collar, D.C.; Day, S.W.; Bishop, K.L.; Wainwright, P.C. Scaling of suction-induced flows in bluegill: Morphological and kinematic predictors for the ontogeny of feeding performance. J. Exp. Biol. 2008, 211, 2658–2668. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Osse, J.; Verhagen, J. A quantitative hydrodynamical model of suction feeding in fish. J. Theor. Biol. 1982, 95, 49–79. [Google Scholar] [CrossRef]
- Aerts, P.; Osse, J.; Verraes, W. Model of jaw depression during feeding in Astatotilapia elegans (Teleostei: Cichlidae): Mechanisms for energy storage and triggering. J. Morphol. 1987, 194, 85–109. [Google Scholar] [CrossRef] [PubMed]
- Camp, A.L.; Roberts, T.J.; Brainerd, E.L. Swimming muscles power suction feeding in largemouth bass. Proc. Natl. Acad. Sci. USA 2015, 112, 8690–8695. [Google Scholar] [CrossRef] [Green Version]
- Camp, A.L.; Roberts, T.J.; Brainerd, E.L. Bluegill sunfish use high power outputs from axial muscles to generate powerful suction-feeding strikes. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Van Wassenbergh, S.; Strother, J.A.; Flammang, B.E.; Ferry-Graham, L.A.; Aerts, P. Extremely fast prey capture in pipefish is powered by elastic recoil. J. R. Soc. Interface 2008, 5, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Poppinga, S.; Weisskopf, C.; Westermeier, A.S.; Masselter, T.; Speck, T. Fastest predators in the plant kingdom: Functional morphology and biomechanics of suction traps found in the largest genus of carnivorous plants. Aob Plants 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Prabhakar, S.; Sane, S.P. The biomechanics of fast prey capture in aquatic bladderworts. Biol. Lett. 2011. [Google Scholar] [CrossRef]
- Vincent, O.; Weißkopf, C.; Poppinga, S.; Masselter, T.; Speck, T.; Joyeux, M.; Quilliet, C.; Marmottant, P. Ultra-fast underwater suction traps. Proc. R. Soc. Lond. B Biol. Sci. 2011, 278, 2909–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermeier, A.S.; Fleischmann, A.; Müller, K.; Schäferhoff, B.; Rubach, C.; Speck, T.; Poppinga, S. Trap diversity and character evolution in carnivorous bladderworts (Utricularia, Lentibulariaceae). Sci. Rep. 2017, 7, 12052. [Google Scholar] [CrossRef] [PubMed]
- Sasago, A.; Sibaoka, T. Water extrusion in the trap bladders of Utricularia vulgaris I. A possible pathway of water across the bladder wall. Bot. Mag. Tokyo 1985, 98, 55–66. [Google Scholar] [CrossRef]
- Sasago, A.; Sibaoka, T. Water extrustion in the trap bladders of Utricularia vulgaris II. A possible mechanism of water outflow. Bot. Mag. Tokyo 1985, 98, 113–124. [Google Scholar] [CrossRef]
- Adamec, L.; Poppinga, S. Measurement of the critical negative pressure inside traps of aquatic carnivorous Utricularia species. Aquat. Bot. 2016, 133, 10–16. [Google Scholar] [CrossRef]
- Sydenham, P.; Findlay, G. The rapid movement of the bladder of Utricularia sp. Aust. J. Biol. Sci. 1973, 26, 1115–1126. [Google Scholar] [CrossRef] [Green Version]
- Poppinga, S.; Daber, L.E.; Westermeier, A.S.; Kruppert, S.; Horstmann, M.; Tollrian, R.; Speck, T. Biomechanical analysis of prey capture in the carnivorous Southern bladderwort (Utricularia australis). Sci. Rep. 2017, 7, 1776. [Google Scholar] [CrossRef]
- Van Wassenbergh, S.; Roos, G.; Genbrugge, A.; Leysen, H.; Aerts, P.; Adriaens, D.; Herrel, A. Suction is kid’s play: Extremely fast suction in newborn seahorses. Biol. Lett. 2009, 5, 200–203. [Google Scholar] [CrossRef]
- Sanford, C.P.; Wainwright, P.C. Use of sonomicrometry demonstrates the link between prey capture kinematics and suction pressure in largemouth bass. J. Exp. Biol. 2002, 205, 3445–3457. [Google Scholar]
- Friday, L. The size and shape of traps of Utricularia vulgaris L. Funct. Ecol. 1991, 5, 602–607. [Google Scholar] [CrossRef]
- Reifenrath, K.; Theisen, I.; Schnitzler, J.; Porembski, S.; Barthlott, W. Trap architecture in carnivorous Utricularia (Lentibulariaceae). Flora-Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 597–605. [Google Scholar] [CrossRef]
- Anto, J.; Majoris, J.; Turingan, R. Prey selection and functional morphology through ontogeny of Amphiprion clarkii with a congeneric comparison. J. Fish Biol. 2009, 75, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Bremigan, M.T.; Stein, R.A. Gape-dependent larval foraging and zooplankton size: Implications for fish recruitment across systems. Can. J. Fish. Aquat. Sci. 1994, 51, 913–922. [Google Scholar] [CrossRef]
- Hernandez, L. Intraspecific scaling of feeding mechanics in an ontogenetic series of zebrafish, Danio rerio. J. Exp. Biol. 2000, 203, 3033–3043. [Google Scholar] [PubMed]
- Krebs, J.M.; Turingan, R.G. Intraspecific variation in gape–prey size relationships and feeding success during early ontogeny in red drum, Sciaenops ocellatus. Environ. Biol. Fishes 2003, 66, 75–84. [Google Scholar] [CrossRef]
- China, V.; Holzman, R. Hydrodynamic starvation in first-feeding larval fishes. Proc. Natl. Acad. Sci. USA 2014, 111, 8083–8088. [Google Scholar] [CrossRef] [Green Version]
- Pekkan, K.; Chang, B.; Uslu, F.; Mani, K.; Chen, C.-Y.; Holzman, R. Characterization of zebrafish larvae suction feeding flow using μPIV and optical coherence tomography. Exp. Fluids 2016, 57, 112. [Google Scholar] [CrossRef] [Green Version]
- Yaniv, S.; Elad, D.; Holzman, R. Suction feeding across fish life stages: Flow dynamics from larvae to adults and implications for prey capture. J. Exp. Biol. 2014, 217, 3748–3757. [Google Scholar] [CrossRef] [Green Version]
- True, A.C.; Crimaldi, J.P. Hydrodynamics of viscous inhalant flows. Phys. Rev. E 2017, 95, 053107. [Google Scholar] [CrossRef] [Green Version]
- Jumars, P.A. Boundary-trapped, inhalant siphon and drain flows: Pipe entry revisited numerically. Limnol. Oceanogr. Fluids Environ. 2013, 3, 21–39. [Google Scholar] [CrossRef]
- Szymanski, P. Quelques Solutions Exactes des Equations de 1′Hydrodynamique de Fluide Visqueux dans le Cas d’un Tube Cylindrique. J. Des Math. Pures Et Appl. 1932, 11, 67–107. [Google Scholar]
- Vrentas, J.; Duda, J. Flow of a Newtonian fluid through a sudden contraction. Appl. Sci. Res. 1973, 28, 241–260. [Google Scholar] [CrossRef]
- Berg, O.; Brown, M.D.; Schwaner, M.J.; Hall, M.R.; Müller, U.K. Hydrodynamics of the bladderwort feeding strike. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Berg, O.; Singh, K.; Hall, M.R.; Schwaner, M.J.; Müller, U.K. Thermodynamics of the bladderwort feeding strike—Suction power from elastic energy storage. Integr. Comp. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Voesenek, C.J.; Muijres, F.T.; Van Leeuwen, J.L. Biomechanics of swimming in developing larval fish. J. Exp. Biol. 2018, 221, jeb149583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, L.A.; Peskin, C.S. When vortices stick: An aerodynamic transition in tiny insect flight. J. Exp. Biol. 2004, 207, 3073–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birch, J.M.; Dickson, W.B.; Dickinson, M.H. Force production and flow structure of the leading edge vortex on flapping wings at high and low Reynolds numbers. J. Exp. Biol. 2004, 207, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- China, V.; Levy, L.; Liberzon, A.; Elmaliach, T.; Holzman, R. Hydrodynamic regime determines the feeding success of larval fish through the modulation of strike kinematics. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170235. [Google Scholar] [CrossRef]
- Drost, M.; Muller, M.; Osse, J. A quantitative hydrodynamical model of suction feeding in larval fishes: The role of frictional forces. Proc. R. Soc. B: Biol. Sci. 1988, 234, 263–281. [Google Scholar]
- D’Amato, P. The Savage Garden, Revised: Cultivating Carnivorous Plants; Ten Speed Press: Berkeley, CA, USA, 2013. [Google Scholar]
- Thielicke, W.; Stamhuis, E. PIVlab–towards user-friendly, affordable and accurate digital particle image velocimetry in MATLAB. J. Open Res. Softw. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Raffel, M.; Willert, C.E.; Wereley, S.T.; Kompenhans, J. Particle Image Velocimetry: A Practical Guide; Springer: Berlin/Heidelberg, Germany, 1998; p. 255. [Google Scholar]
- Westerweel, J. Fundamentals of digital particle image velocimetry. Meas. Sci. Technol. 1997, 8, 1379. [Google Scholar] [CrossRef] [Green Version]
- Sampson, R.A., XII. On Stokes’s current function. Philos. Trans. R. Soc. London 1891, 449–518. [Google Scholar] [CrossRef] [Green Version]
- Dagan, Z.; Weinbaum, S.; Pfeffer, R. An infinite-series solution for the creeping motion through an orifice of finite length. J. Fluid Mech. 1982, 115, 505–523. [Google Scholar] [CrossRef] [Green Version]
- Weissberg, H.L. End correction for slow viscous flow through long tubes. Phys. Fluids 1962, 5, 1033–1036. [Google Scholar] [CrossRef]
- Thompson, B.; Maynes, D.; Webb, B. Characterization of the hydrodynamically developing flow in a microtube using MTV. J. Fluids Eng. 2005, 127, 1003–1012. [Google Scholar] [CrossRef]
- Durst, F.; Loy, T. Investigations of laminar flow in a pipe with sudden contraction of cross sectional area. Comput. Fluids 1985, 13, 15–36. [Google Scholar] [CrossRef]





| Characteristic | Bladderwort Trap 1 | Larval Fish 2 | Adult Fish 3 |
|---|---|---|---|
| peak flow speed (m/s) 4 | 0.04 to 0.16 (2 to 3) | 0.0012 (0.003) | 1 to 7 |
| peak suction pressure (kPa) | 10 to 17 | 0.25 | 31 to 37 |
| gape diameter (mm) 5 | 0.27 to 0.42 (0.1) | 0.12 to 0.30 | 26 |
| time to peak flow (ms) | 0.1 | 10 | 25 |
| Reynolds number 6 | 14 to 43 (250) | 0.1 to 0.4 (0.9) | 7 × 104 to 23 × 104 |
| Womersley number | 4 to 20 | 3 to 4 | 200 to 400 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, K.; Reyes, R.C.; Campa, G., Jr.; Brown, M.D.; Hidalgo, F.; Berg, O.; Müller, U.K. Suction Flows Generated by the Carnivorous Bladderwort Utricularia—Comparing Experiments with Mechanical and Mathematical Models. Fluids 2020, 5, 33. https://doi.org/10.3390/fluids5010033
Singh K, Reyes RC, Campa G Jr., Brown MD, Hidalgo F, Berg O, Müller UK. Suction Flows Generated by the Carnivorous Bladderwort Utricularia—Comparing Experiments with Mechanical and Mathematical Models. Fluids. 2020; 5(1):33. https://doi.org/10.3390/fluids5010033
Chicago/Turabian StyleSingh, Krizma, Roberto C. Reyes, Gabriel Campa, Jr., Matthew D. Brown, Fatima Hidalgo, Otto Berg, and Ulrike K. Müller. 2020. "Suction Flows Generated by the Carnivorous Bladderwort Utricularia—Comparing Experiments with Mechanical and Mathematical Models" Fluids 5, no. 1: 33. https://doi.org/10.3390/fluids5010033
APA StyleSingh, K., Reyes, R. C., Campa, G., Jr., Brown, M. D., Hidalgo, F., Berg, O., & Müller, U. K. (2020). Suction Flows Generated by the Carnivorous Bladderwort Utricularia—Comparing Experiments with Mechanical and Mathematical Models. Fluids, 5(1), 33. https://doi.org/10.3390/fluids5010033