
Long Non-Coding RNAs in the Control of Gametogenesis: Lessons from Fission Yeast


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
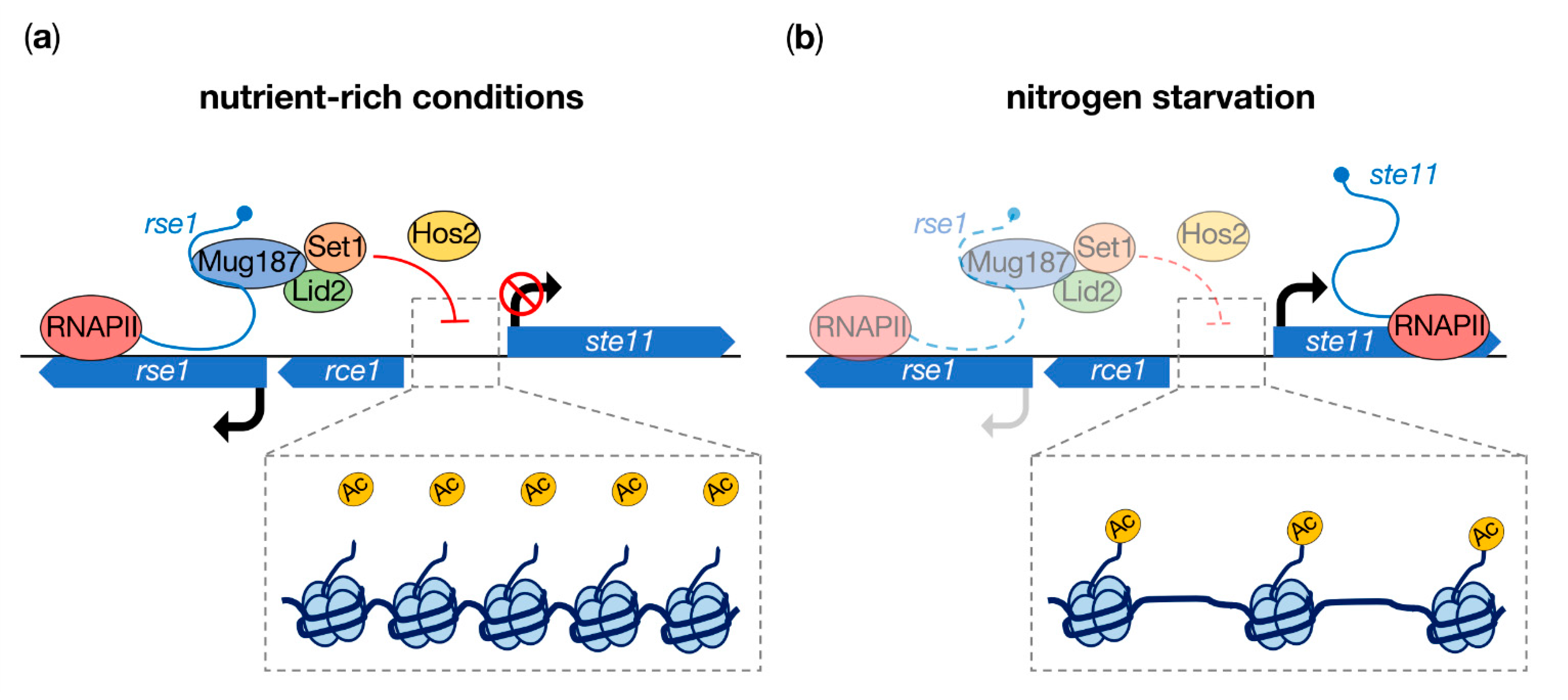
2. The lncRNA rse1 Guides Chromatin Modifiers to Repress in cis the Expression of the Meiosis-Specific Transcription Factor Ste11 in Mitotic Cells
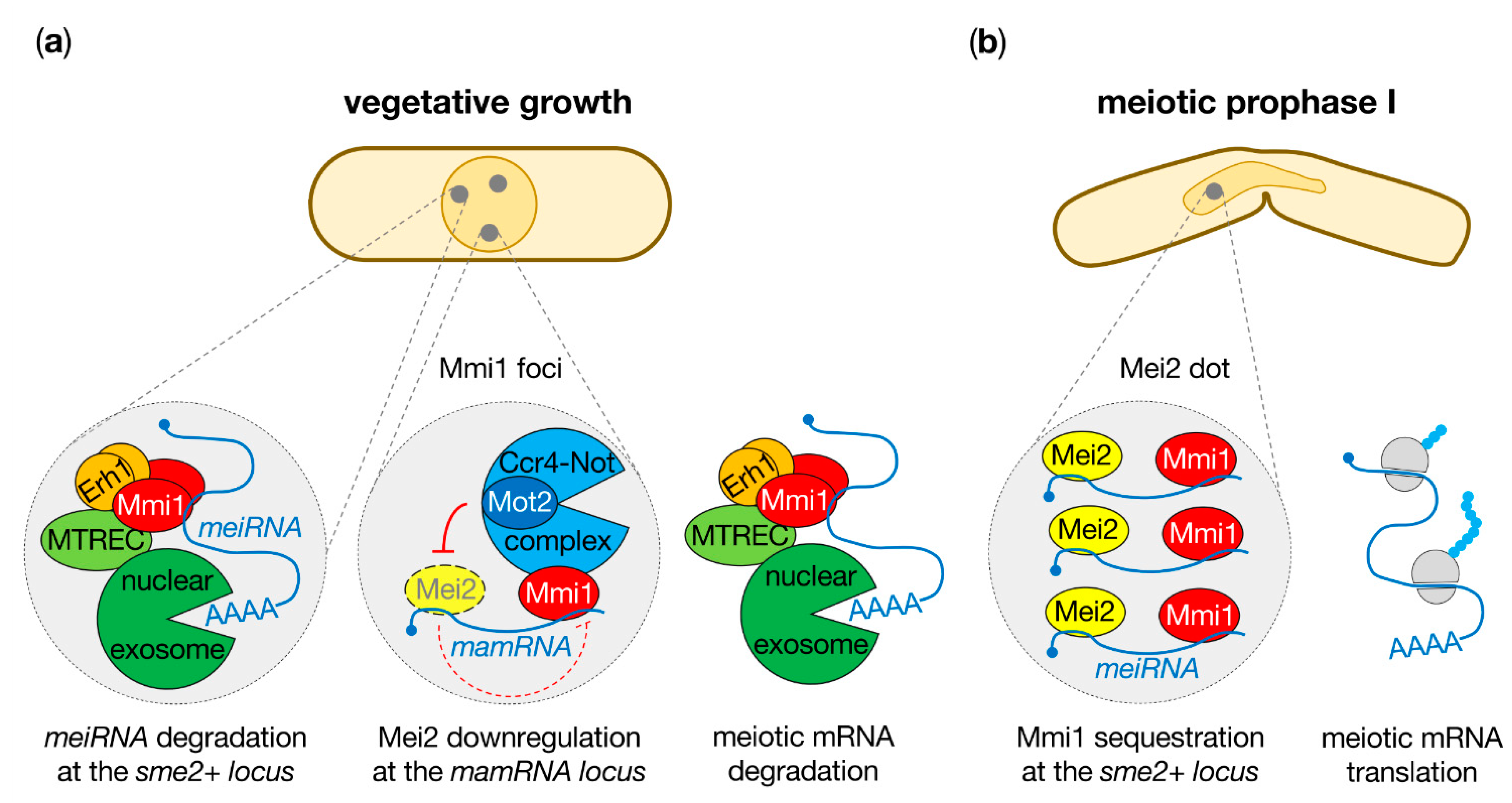
3. The lncRNA meiRNA Lures the YTH-Family RNA-Binding Protein Mmi1 to Promote Expression of Gametogenic Transcripts upon Meiosis Onset
4. The lncRNA mamRNA Scaffolds the Antagonistic RNA-Binding Proteins Mmi1 and Mei2 to Exert Their Mutual Inhibition during Mitosis
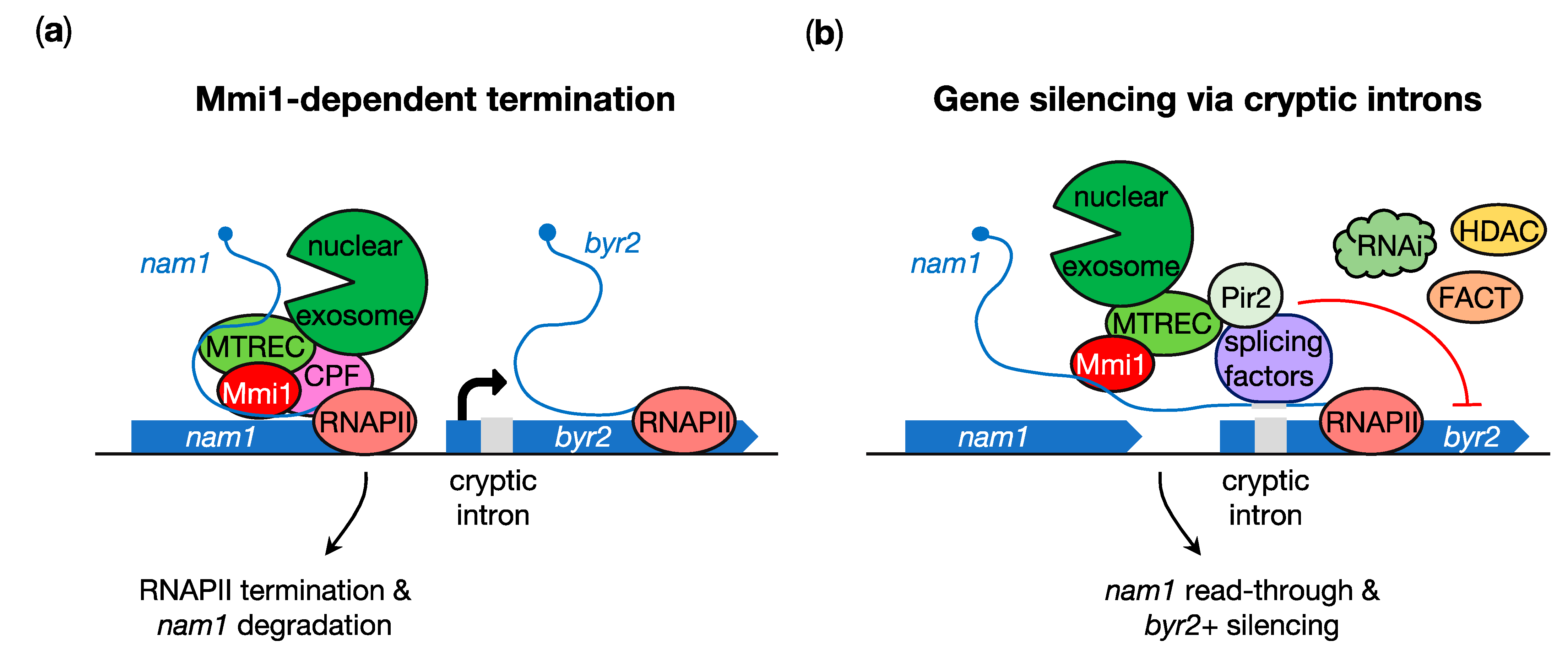
5. Transcription Termination of the lncRNA nam1 Protects Expression of the Downstream Gametogenic byr2+ Gene
6. Homologous Chromosomes Pairing during Meiosis Requires the lncRNAs meiRNA and omt3
7. Widespread Antisense lncRNAs Underlie Global Genome Reprogramming during Meiosis
8. Conclusions and Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Rinn, J.L.; Chang, H.Y. Genome Regulation by Long Noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Wang, Y.; Chen, L.-L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-W.; Kim, H.-W.; Nam, J.-W. The small peptide world in long noncoding RNAs. Brief. Bioinform. 2019, 20, 1853–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, D.; Oberdoerffer, P.; Batista, P.J.; Nussenzweig, A. RNA: A double-edged sword in genome maintenance. Nat. Rev. Genet. 2020, 21, 651–670. [Google Scholar] [CrossRef] [PubMed]
- Michelini, F.; Jalihal, A.P.; Francia, S.; Meers, C.; Neeb, Z.T.; Rossiello, F.; Gioia, U.; Aguado, J.; Jones-Weinert, C.; Luke, B.; et al. From “Cellular” RNA to “Smart” RNA: Multiple Roles of RNA in Genome Stability and Beyond. Chem. Rev. 2018, 118, 4365–4403. [Google Scholar] [CrossRef]
- Batista, P.J.; Chang, H.Y. Long Noncoding RNAs: Cellular Address Codes in Development and Disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, S.U.; Grote, P.; Herrmann, B.G. Mechanisms of long noncoding RNA function in development and disease. Cell. Mol. Life Sci. 2016, 73, 2491–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constanty, F.; Shkumatava, A. lncRNAs in development and differentiation: From sequence motifs to functional characterization. Development 2021, 148, 182741. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M. The selective elimination of messenger RNA underlies the mitosis-meiosis switch in fission yeast. Proc. Jpn. Acad. Ser. B 2010, 86, 788–797. [Google Scholar] [CrossRef] [Green Version]
- Mata, J.; Lyne, R.; Burns, G.; Bähler, J. The transcriptional program of meiosis and sporulation in fission yeast. Nat. Genet. 2002, 32, 143–147. [Google Scholar] [CrossRef]
- Mata, J.; Wilbrey, A.; Bähler, J. Transcriptional regulatory network for sexual differentiation in fission yeast. Genome Biol. 2007, 8, R217. [Google Scholar] [CrossRef] [Green Version]
- Hiriart, E.; Verdel, A. Long noncoding RNA-based chromatin control of germ cell differentiation: A yeast perspective. Chromosom. Res. 2013, 21, 653–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsubo, Y.; Yamamoto, M. Signaling pathways for fission yeast sexual differentiation at a glance. J. Cell Sci. 2012, 125, 2789–2793. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, A.; Iino, Y.; Maeda, T.; Watanabe, Y.; Yamamoto, M. Schizosaccharomyces pombe ste11+ encodes a transcription factor with an HMG motif that is a critical regulator of sexual development. Genes Dev. 1991, 5, 1990–1999. [Google Scholar] [CrossRef] [Green Version]
- Anandhakumar, J.; Fauquenoy, S.; Materne, P.; Migeot, V.; Hermand, D. Regulation of entry into gametogenesis by Ste11: The endless game. Biochem. Soc. Trans. 2013, 41, 1673–1678. [Google Scholar] [CrossRef]
- Mata, J.; Bahler, J. Global roles of Ste11p, cell type, and pheromone in the control of gene expression during early sexual differentiation in fission yeast. Proc. Natl. Acad. Sci. USA 2006, 103, 15517–15522. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.; Hoe, K.-L.; Yu, Y.M.; Yeon, J.-H.; Maeng, P.J. The Fission Yeast GATA Factor, Gaf1, Modulates Sexual Development via Direct Down-Regulation of ste11+ Expression in Response to Nitrogen Starvation. PLoS ONE 2012, 7, e42409. [Google Scholar] [CrossRef] [PubMed]
- Fauquenoy, S.; Migeot, V.; Finet, O.; Yague-Sanz, C.; Khorosjutina, O.; Ekwall, K.; Hermand, D. Repression of Cell Differentiation by a cis-Acting lincRNA in Fission Yeast. Curr. Biol. 2018, 28, 383–391.e3. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Huarte, M.; Zaratiegui, M.; Vaughn, M.; Shi, Y.; Martienssen, R.; Cande, W.Z. Lid2 Is Required for Coordinating H3K4 and H3K9 Methylation of Heterochromatin and Euchromatin. Cell 2008, 135, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roguev, A.; Schaft, D.; Shevchenko, A.; Aasland, R.; Shevchenko, A.; Stewart, A.F. High Conservation of the Set1/Rad6 Axis of Histone 3 Lysine 4 Methylation in Budding and Fission Yeasts. J. Biol. Chem. 2003, 278, 8487–8493. [Google Scholar] [CrossRef] [Green Version]
- Materne, P.; Anandhakumar, J.; Migeot, V.; Soriano, I.; Yague-Sanz, C.; Hidalgo, E.; Mignion, C.; Quintales, L.; Antequera, F.; Hermand, D. Promoter nucleosome dynamics regulated by signalling through the CTD code. eLife 2015, 4, e09008. [Google Scholar] [CrossRef] [Green Version]
- Kunitomo, H.; Higuchi, T.; Iino, Y.; Yamamoto, M. A Zinc-Finger Protein, Rst2p, Regulates Transcription of the Fission Yeast ste11 + Gene, Which Encodes a Pivotal Transcription Factor for Sexual Development. Mol. Biol. Cell 2000, 11, 3205–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Werven, F.; Neuert, G.; Hendrick, N.; Lardenois, A.; Buratowski, S.; van Oudenaarden, A.; Primig, M.; Amon, A. Transcription of Two Long Noncoding RNAs Mediates Mating-Type Control of Gametogenesis in Budding Yeast. Cell 2012, 150, 1170–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretto, F.; Wood, N.E.; Kelly, G.; Doncic, A.; Van Werven, F.J. A regulatory circuit of two lncRNAs and a master regulator directs cell fate in yeast. Nat. Commun. 2018, 9, 780. [Google Scholar] [CrossRef] [PubMed]
- Moretto, F.; Wood, N.E.; Chia, M.; Li, C.; Luscombe, N.M.; van Werven, F.J. Transcription levels of a noncoding RNA orchestrate opposing regulatory and cell fate outcomes in yeast. Cell Rep. 2021, 34, 108643. [Google Scholar] [CrossRef]
- Harigaya, Y.; Tanaka, H.; Yamanaka, S.; Tanaka, K.; Watanabe, Y.; Tsutsumi, C.; Chikashige, Y.; Hiraoka, Y.; Yamashita, A.; Yamamoto, M. Selective elimination of messenger RNA prevents an incidence of untimely meiosis. Nat. Cell Biol. 2006, 442, 45–50. [Google Scholar] [CrossRef]
- Yamashita, A.; Shichino, Y.; Tanaka, H.; Hiriart, E.; Touat-Todeschini, L.; Vavasseur, A.; Ding, D.-Q.; Hiraoka, Y.; Verdel, A.; Yamamoto, M. Hexanucleotide motifs mediate recruitment of the RNA elimination machinery to silent meiotic genes. Open Biol. 2012, 2, 120014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilchert, C.; Wittmann, S.; Passoni, M.; Shah, S.; Granneman, S.; Vasiljeva, L. Regulation of mRNA Levels by Decay-Promoting Introns that Recruit the Exosome Specificity Factor Mmi1. Cell Rep. 2015, 13, 2504–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touat-Todeschini, L.; Shichino, Y.; Dangin, M.; Thierry-Mieg, N.; Gilquin, B.; Hiriart, E.; Sachidanandam, R.; Lambert, E.; Brettschneider, J.; Reuter, M.; et al. Selective termination of lnc RNA transcription promotes heterochromatin silencing and cell differentiation. EMBO J. 2017, 36, 2626–2641. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, Y.; Bao, H.; Jiang, Y.; Xu, C.; Wu, J.; Shi, Y. A novel RNA-binding mode of the YTH domain reveals the mechanism for recognition of determinant of selective removal by Mmi1. Nucleic Acids Res. 2015, 44, 969–982. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Xu, J.; Su, S.; Liu, H.; Gan, J.; Ma, J. Structural insights into the specific recognition of DSR by the YTH domain containing protein Mmi1. Biochem. Biophys. Res. Commun. 2017, 491, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Thillainadesan, G.; Chalamcharla, V.R.; Meng, Z.; Balachandran, V.; Dhakshnamoorthy, J.; Zhou, M.; Grewal, S.I. Enhancer of Rudimentary Cooperates with Conserved RNA-Processing Factors to Promote Meiotic mRNA Decay and Facultative Heterochromatin Assembly. Mol. Cell 2016, 61, 747–759.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.; Vo, T.V.; Thillainadesan, G.; Holla, S.; Zhang, B.; Jiang, Y.; Lv, M.; Xu, Z.; Wang, C.; Balachandran, V.; et al. A conserved dimer interface connects ERH and YTH family proteins to promote gene silencing. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hazra, D.; Andrić, V.; Palancade, B.; Rougemaille, M.; Graille, M. Formation of S. pombe Erh1 homodimer mediates gametogenic gene silencing and meiosis progression. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- McPheeters, D.S.; Cremona, N.; Sunder, S.; Chen, H.-M.; Averbeck, N.; Leatherwood, J.; Wise, J.A. A complex gene regulatory mechanism that operates at the nexus of multiple RNA processing decisions. Nat. Struct. Mol. Biol. 2009, 16, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-M.; Futcher, B.; Leatherwood, J. The Fission Yeast RNA Binding Protein Mmi1 Regulates Meiotic Genes by Controlling Intron Specific Splicing and Polyadenylation Coupled RNA Turnover. PLoS ONE 2011, 6, e26804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St-André, O.; Lemieux, C.; Perreault, A.; Lackner, D.H.; Bähler, J.; Bachand, F. Negative Regulation of Meiotic Gene Expression by the Nuclear Poly(a)-binding Protein in Fission Yeast. J. Biol. Chem. 2010, 285, 27859–27868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, S.; Yamashita, A.; Harigaya, Y.; Iwata, R.; Yamamoto, M. Importance of polyadenylation in the selective elimination of meiotic mRNAs in growing S. pombe cells. EMBO J. 2010, 29, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hung, S.; Esnault, C.; Pathak, R.; Johnson, K.R.; Bankole, O.; Yamashita, A.; Zhang, H.; Levin, H.L. Dense Transposon Integration Reveals Essential Cleavage and Polyadenylation Factors Promote Heterochromatin Formation. Cell Rep. 2020, 30, 2686–2698.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.N.; Chalamcharla, V.R.; Reyes-Turcu, F.; Mehta, S.; Zofall, M.; Balachandran, V.; Dhakshnamoorthy, J.; Taneja, N.; Yamanaka, S.; Zhou, M.; et al. Mtr4-like Protein Coordinates Nuclear RNA Processing for Heterochromatin Assembly and for Telomere Maintenance. Cell 2013, 155, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Takayama, T.; Iwata, R.; Yamamoto, M. A novel factor Iss10 regulates Mmi1-mediated selective elimination of meiotic transcripts. Nucleic Acids Res. 2013, 41, 9680–9687. [Google Scholar] [CrossRef] [Green Version]
- Egan, E.; Braun, C.R.; Gygi, S.P.; Moazed, D. Post-transcriptional regulation of meiotic genes by a nuclear RNA silencing complex. RNA 2014, 20, 867–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhu, J.; Schermann, G.; Ohle, C.; Bendrin, K.; Sugioka-Sugiyama, R.; Sugiyama, T.; Fischer, T. The fission yeast MTREC complex targets CUTs and unspliced pre-mRNAs to the nuclear exosome. Nat. Commun. 2015, 6, 7050. [Google Scholar] [CrossRef] [Green Version]
- Shichino, Y.; Otsubo, Y.; Yamamoto, M.; Yamashita, A. Meiotic gene silencing complex MTREC/NURS recruits the nuclear exosome to YTH-RNA-binding protein Mmi1. PLoS Genet. 2020, 16, e1008598. [Google Scholar] [CrossRef]
- Shichino, Y.; Otsubo, Y.; Kimori, Y.; Yamamoto, M.; Yamashita, A. YTH-RNA-binding protein prevents deleterious expression of meiotic proteins by tethering their mRNAs to nuclear foci. eLife 2018, 7, 32155. [Google Scholar] [CrossRef] [Green Version]
- Shichino, Y.; Yamashita, A.; Yamamoto, M. Meiotic long non-coding meiRNA accumulates as a dot at its genetic locus facilitated by Mmi1 and plays as a decoy to lure Mmi1. Open Biol. 2014, 4, 140022. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Yamamoto, M.S. Pombe mei2+ encodes an RNA-binding protein essential for premeiotic DNA synthesis and meiosis I, which cooperates with a novel RNA species meiRNA. Cell 1994, 78, 487–498. [Google Scholar] [CrossRef]
- Yamashita, A. meiRNA, A Polyvalent Player in Fission Yeast Meiosis. Non-Coding RNA 2019, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, T.; Sugioka-Sugiyama, R. Red1 promotes the elimination of meiosis-specific mRNAs in vegetatively growing fission yeast. EMBO J. 2011, 30, 1027–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Lee, N.N.; Pan, L.; Dhakshnamoorthy, J.; Sun, L.-L.; Zofall, M.; Wheeler, D.; Grewal, S.I.S. TOR targets an RNA processing network to regulate facultative heterochromatin, developmental gene expression and cell proliferation. Nat. Cell Biol. 2021, 23, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Cotobal, C.; Rodríguez-López, M.; Duncan, C.; Hasan, A.; Yamashita, A.; Yamamoto, M.; Bähler, J.; Mata, J. Role of Ccr4-Not complex in heterochromatin formation at meiotic genes and subtelomeres in fission yeast. Epigenetics Chromatin 2015, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukleja, M.; Cuéllar, J.; Siwaszek, A.; Kasprzak, J.M.; Czarnocki-Cieciura, M.; Bujnicki, J.M.; Dziembowski, A.; Valpuesta, J. The architecture of the Schizosaccharomyces pombe CCR4-NOT complex. Nat. Commun. 2016, 7, 10433. [Google Scholar] [CrossRef] [Green Version]
- Stowell, J.A.; Webster, M.; Kögel, A.; Wolf, J.; Shelley, K.L.; Passmore, L.A. Reconstitution of Targeted Deadenylation by the Ccr4-Not Complex and the YTH Domain Protein Mmi1. Cell Rep. 2016, 17, 1978–1989. [Google Scholar] [CrossRef] [Green Version]
- Simonetti, F.; Candelli, T.; Leon, S.; Libri, D.; Rougemaille, M. Ubiquitination-dependent control of sexual differentiation in fission yeast. eLife 2017, 6, e28046. [Google Scholar] [CrossRef]
- Miller, J.E.; Reese, J.C. Ccr4-Not complex: The control freak of eukaryotic cells. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 315–333. [Google Scholar] [CrossRef] [Green Version]
- Hagkarim, N.C.; Grand, R. The Regulatory Properties of the Ccr4-Not Complex. Cells 2020, 9, 2379. [Google Scholar] [CrossRef] [PubMed]
- Andric, V.; Nevers, A.; Hazra, D.; Auxilien, S.; Menant, A.; Graille, M.; Palancade, B.; Rougemaille, M. A scaffold lncRNA shapes the mitosis to meiosis switch. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Wilusz, J.E. Long noncoding RNAs: Re-writing dogmas of RNA processing and stability. Biochim. Biophys. Acta BBA Bioenerg. 2016, 1859, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Ernst, E.H.; Nielsen, J.; Ipsen, M.B.; Villesen, P.; Lykke-Hartmann, K. Transcriptome Analysis of Long Non-coding RNAs and Genes Encoding Paraspeckle Proteins During Human Ovarian Follicle Development. Front. Cell Dev. Biol. 2018, 6, 78. [Google Scholar] [CrossRef]
- Hosseini, E.S.; Meryet-Figuiere, M.; Sabzalipoor, H.; Kashani, H.H.; Nikzad, H.; Asemi, Z. Dysregulated expression of long noncoding RNAs in gynecologic cancers. Mol. Cancer 2017, 16, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.-S.; Li, T.-P.; Ton, H.; Mao, X.-D.; Chen, Y.-J. Advances of Long Noncoding RNAs-mediated Regulation in Reproduction. Chin. Med. J. 2018, 131, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Abdelmohsen, K.; Kim, J.; Yang, X.; Martindale, J.L.; Tominaga-Yamanaka, K.; White, E.J.; Orjalo, A.V.; Rinn, J.L.; Kreft, S.G.; et al. Scaffold function of long non-coding RNA HOTAIR in protein ubiquitination. Nat. Commun. 2013, 4, 2939. [Google Scholar] [CrossRef]
- Xue, M.; Chen, L.-Y.; Wang, W.-J.; Su, T.-T.; Shi, L.-H.; Wang, L.; Zhang, W.; Si, J.-M.; Wang, L.-J.; Chen, S.-J. HOTAIR induces the ubiquitination of Runx3 by interacting with Mex3b and enhances the invasion of gastric cancer cells. Gastric Cancer 2018, 21, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalamcharla, V.R.; Folco, H.D.; Dhakshnamoorthy, J.; Grewal, S.I.S. Conserved factor Dhp1/Rat1/Xrn2 triggers premature transcription termination and nucleates heterochromatin to promote gene silencing. Proc. Natl. Acad. Sci. USA 2015, 112, 15548–15555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo, T.V.; Dhakshnamoorthy, J.; Larkin, M.; Zofall, M.; Thillainadesan, G.; Balachandran, V.; Holla, S.; Wheeler, D.; Grewal, S.I. CPF Recruitment to Non-canonical Transcription Termination Sites Triggers Heterochromatin Assembly and Gene Silencing. Cell Rep. 2019, 28, 267–281.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ard, R.; Tong, P.; Allshire, R.C. Long non-coding RNA-mediated transcriptional interference of a permease gene confers drug tolerance in fission yeast. Nat. Commun. 2014, 5, 5576. [Google Scholar] [CrossRef]
- Shah, S.; Wittmann, S.; Kilchert, C.; Vasiljeva, L. lncRNA recruits RNAi and the exosome to dynamically regulate pho1 expression in response to phosphate levels in fission yeast. Genes Dev. 2014, 28, 231–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Sanchez, A.M.; Goldgur, Y.; Shuman, S.; Schwer, B. Transcription of lncRNA prt, clustered prt RNA sites for Mmi1 binding, and RNA polymerase II CTD phospho-sites govern the repression of pho1 gene expression under phosphate-replete conditions in fission yeast. RNA 2016, 22, 1011–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, A.M.; Shuman, S.; Schwer, B. Poly(A) site choice and Pol2 CTD Serine-5 status govern lncRNA control of phosphate-responsive tgp1 gene expression in fission yeast. RNA 2018, 24, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Shuman, S. Transcriptional interference at tandem lncRNA and protein-coding genes: An emerging theme in regulation of cellular nutrient homeostasis. Nucleic Acids Res. 2020, 48, 8243–8254. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, H.P.; Riggs, M.; Rodgers, L.; Wigler, M. byr2, a Schizosaccharomyces pombe gene encoding a protein kinase capable of partial suppression of the ras1 mutant phenotype. Mol. Cell. Biol. 1991, 11, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Styrkarsdottir, U.; Egel, R.; Nielsen, O. Functional conservation between Schizosaccharomyces pombe ste8 and Saccharomyces cerevisiae STE11 protein kinases in yeast signal transduction. Mol. Genet. Genom. 1992, 235, 122–130. [Google Scholar] [CrossRef]
- Thillainadesan, G.; Xiao, H.; Holla, S.; Dhakshnamoorthy, J.; Jenkins, L.M.M.; Wheeler, D.; Grewal, S.I.S. Conserved protein Pir2ARS2 mediates gene repression through cryptic introns in lncRNAs. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Chikashige, Y.; Ding, D.Q.; Funabiki, H.; Haraguchi, T.; Mashiko, S.; Yanagida, M.; Hiraoka, Y. Telomere-led premeiotic chromosome movement in fission yeast. Science 1994, 264, 270–273. [Google Scholar] [CrossRef]
- Hiraoka, Y. Meiotic telomeres: A matchmaker for homologous chromosomes. Genes Cells 1998, 3, 405–413. [Google Scholar] [CrossRef]
- Ding, D.-Q.; Yamamoto, A.; Haraguchi, T.; Hiraoka, Y. Dynamics of Homologous Chromosome Pairing during Meiotic Prophase in Fission Yeast. Dev. Cell 2004, 6, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.L.; Pryce, D.W.; McFarlane, R.J. Homologous chromosome pairing in Schizosaccharomyces pombe. Yeast 2006, 23, 977–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraoka, Y.; Ding, D.-Q.; Yamamoto, A.; Tsutsumi, C.; Chikashige, Y. Characterization of fission yeast meiotic mutants based on live observation of meiotic prophase nuclear movement. Chromosoma 2000, 109, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.-Q.; Okamasa, K.; Yamane, M.; Tsutsumi, C.; Haraguchi, T.; Yamamoto, M.; Hiraoka, Y. Meiosis-Specific Noncoding RNA Mediates Robust Pairing of Homologous Chromosomes in Meiosis. Science 2012, 336, 732–736. [Google Scholar] [CrossRef]
- Ding, D.-Q.; Haraguchi, T.; Hiraoka, Y. The role of chromosomal retention of noncoding RNA in meiosis. Chromosom. Res. 2013, 21, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.-Q.; Okamasa, K.; Katou, Y.; Oya, E.; Nakayama, J.-I.; Chikashige, Y.; Shirahige, K.; Haraguchi, T.; Hiraoka, Y. Chromosome-associated RNA–protein complexes promote pairing of homologous chromosomes during meiosis in Schizosaccharomyces pombe. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Hyman, A.A.; Weber, C.A.; Julicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, T.; Souquere, S.; Chujo, T.; Kobelke, S.; Chong, Y.S.; Fox, A.H.; Bond, C.S.; Nakagawa, S.; Pierron, G.; Hirose, T. Functional Domains of NEAT1 Architectural lncRNA Induce Paraspeckle Assembly through Phase Separation. Mol. Cell 2018, 70, 1038–1053.e7. [Google Scholar] [CrossRef] [Green Version]
- Kakihara, Y.; Nabeshima, K.; Hirata, A.; Nojima, H. Overlapping omt1 + and omt2 + genes are required for spore wall maturation in Schizosaccharomyces pombe. Genes Cells 2003, 8, 547–558. [Google Scholar] [CrossRef]
- Ni, T.; Tu, K.; Wang, Z.; Song, S.; Wu, H.; Xie, B.; Scott, K.C.; Grewal, S.I.; Gao, Y.; Zhu, J. The Prevalence and Regulation of Antisense Transcripts in Schizosaccharomyces pombe. PLoS ONE 2010, 5, e15271. [Google Scholar] [CrossRef] [Green Version]
- Rhind, N.; Chen, Z.; Yassour, M.; Thompson, D.A.; Haas, B.J.; Habib, N.; Wapinski, I.; Roy, S.; Lin, M.F.; Heiman, D.; et al. Comparative Functional Genomics of the Fission Yeasts. Science 2011, 332, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Bitton, D.A.; Grallert, A.; Scutt, P.J.; Yates, T.; Li, Y.; Bradford, J.; Hey, Y.; Pepper, S.D.; Hagan, I.; Miller, C. Programmed fluctuations in sense/antisense transcript ratios drive sexual differentiation in S. pombe. Mol. Syst. Biol. 2011, 7, 559. [Google Scholar] [CrossRef]
- Chen, H.-M.; Rosebrock, A.P.; Khan, S.R.; Futcher, B.; Leatherwood, J.K. Repression of Meiotic Genes by Antisense Transcription and by Fkh2 Transcription Factor in Schizosaccharomyces pombe. PLoS ONE 2012, 7, e29917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hongay, C.F.; Grisafi, P.L.; Galitski, T.; Fink, G.R. Antisense Transcription Controls Cell Fate in Saccharomyces cerevisiae. Cell 2006, 127, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Gelfand, B.; Mead, J.; Bruning, A.; Apostolopoulos, N.; Tadigotla, V.; Nagaraj, V.; Sengupta, A.M.; Vershon, A.K. Regulated Antisense Transcription Controls Expression of Cell-Type-Specific Genes in Yeast. Mol. Cell. Biol. 2011, 31, 1701–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, S.R.; Marguerat, S.; Bitton, D.A.; Rodríguez-López, M.; Rallis, C.; Lemay, J.-F.; Cotobal, C.; Malecki, M.; Smialowski, P.; Mata, J.; et al. Long noncoding RNA repertoire and targeting by nuclear exosome, cytoplasmic exonuclease, and RNAi in fission yeast. RNA 2018, 24, 1195–1213. [Google Scholar] [CrossRef] [Green Version]
- Wery, M.; Gautier, C.; Descrimes, M.; Yoda, M.; Vennin-Rendos, H.; Migeot, V.; Gautheret, D.; Hermand, D.; Morillon, A. Native elongating transcript sequencing reveals global anti-correlation between sense and antisense nascent transcription in fission yeast. RNA 2017, 24, 196–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 880–893. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andric, V.; Rougemaille, M. Long Non-Coding RNAs in the Control of Gametogenesis: Lessons from Fission Yeast. Non-Coding RNA 2021, 7, 34. https://doi.org/10.3390/ncrna7020034
Andric V, Rougemaille M. Long Non-Coding RNAs in the Control of Gametogenesis: Lessons from Fission Yeast. Non-Coding RNA. 2021; 7(2):34. https://doi.org/10.3390/ncrna7020034
Chicago/Turabian StyleAndric, Vedrana, and Mathieu Rougemaille. 2021. "Long Non-Coding RNAs in the Control of Gametogenesis: Lessons from Fission Yeast" Non-Coding RNA 7, no. 2: 34. https://doi.org/10.3390/ncrna7020034
APA StyleAndric, V., & Rougemaille, M. (2021). Long Non-Coding RNAs in the Control of Gametogenesis: Lessons from Fission Yeast. Non-Coding RNA, 7(2), 34. https://doi.org/10.3390/ncrna7020034