
Interdependent Transcription of a Natural Sense/Antisense Transcripts Pair (SLC34A1/PFN3)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
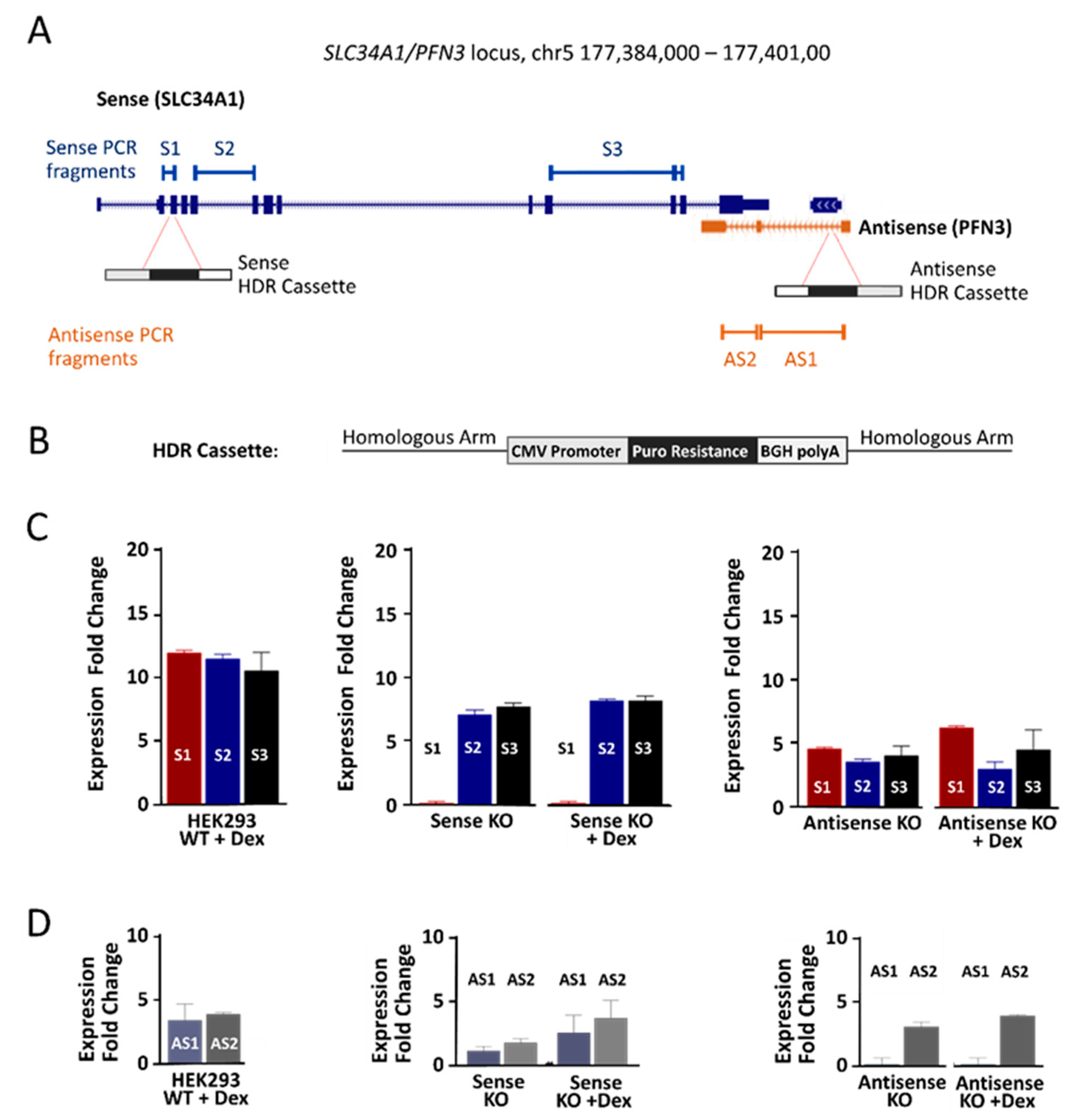
2.1. The SLC34A1/PFN3 Locus
2.2. Drug-Induced Expression of SLC34A1/PFN3 Sense-Antisense Transcripts
2.3. Epigenetic Changes in Response to Zebularine and Dexamethasone
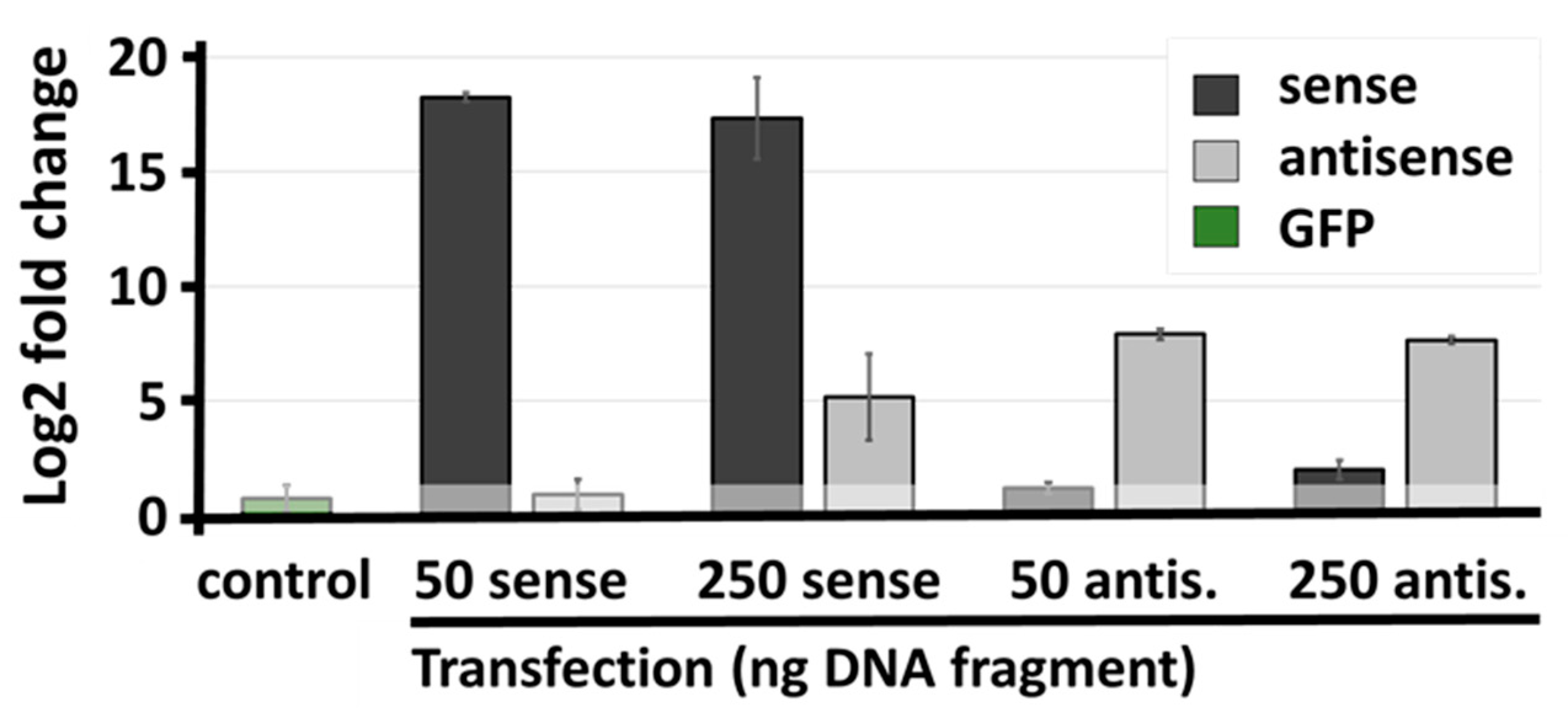
2.4. Genetic Interference with SLC34A1/PFN3 Sense–Antisense Expression
2.5. Protein Expression
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beiter, T.; Reich, E.; Williams, R.W.; Simon, P. Antisense transcription: A critical look in both directions. Cell. Mol. Life Sci. 2009, 66, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Faghihi, M.A.; Wahlestedt, C. Regulatory roles of natural antisense transcripts. Nat. Rev. Mol. Cell Biol. 2009, 10, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Zinad, H.S.; Natasya, I.; Werner, A. Natural antisense transcripts at the interface between host genome and mobile genetic elements. Front. Microbiol. 2017, 8, 2292. [Google Scholar] [CrossRef] [PubMed]
- Shendure, J.; Church, G.M. Computational discovery of sense-antisense transcription in the human and mouse genomes. Genome Biol. 2002, 3, RESEARCH0044. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sun, M.; Kent, W.J.; Huang, X.; Xie, H.; Wang, W.; Zhou, G.; Shi, R.Z.; Rowley, J.D. Over 20% of human transcripts might form sense-antisense pairs. Nucleic Acids Res. 2004, 32, 4812–4820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyosawa, H.; Yamanaka, I.; Osato, N.; Kondo, S.; Hayashizaki, Y. Antisense transcripts with FANTOM2 clone set and their implications for gene regulation. Genom. Res. 2003, 13, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- Piatek, M.J.; Henderson, V.; Zynad, H.S.; Werner, A. Natural antisense transcription from a comparative perspective. Genomics 2016, 108, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Katayama, S.; Tomaru, Y.; Kasukawa, T.; Waki, K.; Nakanishi, M.; Nakamura, M.; Nishida, H.; Yap, C.C.; Suzuki, M.; Kawai, J.; et al. Antisense transcription in the mammalian transcriptome. Science 2005, 309, 1564–1566. [Google Scholar] [CrossRef]
- Ling, M.H.; Ban, Y.; Wen, H.; Wang, S.M.; Ge, S.X. Conserved expression of natural antisense transcripts in mammals. BMC Genom. 2013, 14, 243. [Google Scholar] [CrossRef] [Green Version]
- Wood, E.J.; Chin-Inmanu, K.; Jia, H.; Lipovich, L. Sense-antisense gene pairs: Sequence, transcription, and structure are not conserved between human and mouse. Front. Genet. 2013, 4, 183. [Google Scholar] [CrossRef] [Green Version]
- Lapidot, M.; Pilpel, Y. Genome-wide natural antisense transcription: Coupling its regulation to its different regulatory mechanisms. EMBO Rep. 2006, 7, 1216–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, A.; Carlile, M.; Swan, D. What do natural antisense transcripts regulate? RNA Biol. 2009, 6, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wight, M.; Werner, A. The functions of natural antisense transcripts. Essays Biochem. 2013, 54, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, Y.; Song, Y.; Wang, S.S.; Cheng, K.W.; Zhang, Z.Q.; Yao, J.; Zhou, L.N.; Ling, Z.Y.; Cao, C. LncRNA PINK1-AS promotes Gαi1-driven gastric cancer tumorigenesis by sponging microRNA-200a. Oncogene 2021, 40, 3826–3844. [Google Scholar] [CrossRef]
- Werner, A.; Clark, J.E.; Samaranayake, C.; Casement, J.; Zinad, H.S.; Sadeq, S.; Al-Hashimi, S.; Smith, M.; Kotaja, N.; Mattick, J.S. Widespread formation of double-stranded RNAs in testis. Genom. Res. 2021, 31, 1174–1186. [Google Scholar] [CrossRef]
- Hur, S. Double-Stranded RNA Sensors and modulators in innate immunity. Annu. Rev. Immunol. 2019, 37, 349–375. [Google Scholar] [CrossRef]
- Sadeq, S.; Al-Hashimi, S.; Cusack, C.M.; Werner, A. Endogenous double-stranded RNA. Non-Cod. RNA 2021, 7, 15. [Google Scholar] [CrossRef]
- Dhir, A.; Dhir, S.; Borowski, L.S.; Jimenez, L.; Teitell, M.; Rotig, A.; Crow, Y.J.; Rice, G.I.; Duffy, D.; Tamby, C.; et al. Mitochondrial double-stranded RNA triggers antiviral signalling in humans. Nature 2018, 560, 238–242. [Google Scholar] [CrossRef]
- Kim, Y.; Park, J.; Kim, S.; Kim, M.; Kang, M.G.; Kwak, C.; Kang, M.; Kim, B.; Rhee, H.W.; Kim, V.N. PKR Senses nuclear and mitochondrial signals by interacting with endogenous double-stranded RNAs. Mol. Cell 2018, 71, 1051–1063. [Google Scholar] [CrossRef] [Green Version]
- Werner, A.; Cockell, S.; Falconer, J.; Carlile, M.; Alnumeir, S.; Robinson, J. Contribution of natural antisense transcription to an endogenous siRNA signature in human cells. BMC Genom. 2014, 15, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlile, M.; Nalbant, P.; Preston-Fayers, K.; McHaffie, G.S.; Werner, A. Processing of naturally occurring sense/antisense transcripts of the vertebrate Slc34a gene into short RNAs. Physiol. Genom. 2008, 34, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlile, M.; Swan, D.; Jackson, K.; Preston-Fayers, K.; Ballester, B.; Flicek, P.; Werner, A. Strand selective generation of endo-siRNAs from the Na/phosphate transporter gene Slc34a1 in murine tissues. Nucleic Acids Res. 2009, 37, 2274–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Hennig, G.W.; Wu, Q.; Jose, C.; Zheng, H.; Yan, W. Male germ cells express abundant endogenous siRNAs. Proc. Natl. Acad. Sci. USA 2011, 108, 13159–13164. [Google Scholar] [CrossRef] [Green Version]
- Murer, H.; Forster, I.; Biber, J. The sodium phosphate cotransporter family SLC34. Pflugers Arch. 2004, 447, 763–767. [Google Scholar] [CrossRef]
- Chen, J.; Sun, M.; Hurst, L.D.; Carmichael, G.G.; Rowley, J.D. Human antisense genes have unusually short introns: Evidence for selection for rapid transcription. Trends Genet. 2005, 21, 203–207. [Google Scholar] [CrossRef]
- Murk, K.; Ornaghi, M.; Schiweck, J. Profilin isoforms in health and disease—All the same but different. Front. Cell Dev. Biol. 2021, 9, 681122. [Google Scholar] [CrossRef]
- Malmstrom, K.; Murer, H. Parathyroid hormone inhibits phosphate transport in OK cells but not in LLC-PK1 and JTC-12.P3 cells. Am. J. Physiol. 1986, 251, C23–C31. [Google Scholar] [CrossRef]
- Brown, C.D.; Sayer, R.; Windass, A.S.; Haslam, I.S.; De Broe, M.E.; D’Haese, P.C.; Verhulst, A. Characterisation of human tubular cell monolayers as a model of proximal tubular xenobiotic handling. Toxicol. Appl. Pharmacol. 2008, 233, 428–438. [Google Scholar] [CrossRef]
- Okazaki, Y.; Furuno, M.; Kasukawa, T.; Adachi, J.; Bono, H.; Kondo, S.; Nikaido, I.; Osato, N.; Saito, R.; Suzuki, H.; et al. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature 2002, 420, 563–573. [Google Scholar]
- Liu, Y.; Han, X.; Yuan, J.; Geng, T.; Chen, S.; Hu, X.; Cui, I.H.; Cui, H. Biallelic insertion of a transcriptional terminator via the CRISPR/Cas9 system efficiently silences expression of protein-coding and non-coding RNA genes. J. Biol. Chem. 2017, 292, 5624–5633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, A. Natural antisense transcripts. RNA Biol. 2005, 2, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Lai, E.C. Endogenous small interfering RNAs in animals. Nat. Rev. Mol. Cell. Biol. 2008, 9, 673–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Totoki, Y.; Toyoda, A.; Kaneda, M.; Kuramochi-Miyagawa, S.; Obata, Y.; Chiba, H.; Kohara, Y.; Kono, T.; Nakano, T.; et al. Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes. Nature 2008, 453, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Fiskin, E.; Gutschner, T.; Polycarpou-Schwarz, M.; Gross, M.; Neugebauer, J.; Gandhi, M.; Caudron-Herger, M.; Benes, V.; Diederichs, S. A cautionary tale of sense-antisense gene pairs: Independent regulation despite inverse correlation of expression. Nucleic Acids Res. 2017, 45, 12496–12508. [Google Scholar] [CrossRef]
- Rezaei-Lotfi, S.; Vujovic, F.; Simonian, M.; Hunter, N.; Farahani, R.M. Programmed genomic instability regulates neural transdifferentiation of human brain microvascular pericytes. Genom. Biol. 2021, 22, 334. [Google Scholar] [CrossRef]
- Ebisuya, M.; Yamamoto, T.; Nakajima, M.; Nishida, E. Ripples from neighbouring transcription. Nat. Cell Biol. 2008, 10, 1106–1113. [Google Scholar] [CrossRef]
- Ghanbarian, A.T.; Hurst, L.D. Neighboring genes show correlated evolution in gene expression. Mol. Biol. Evol. 2015, 32, 1748–1766. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, P.; Kustatscher, G.; Rappsilber, J. Epigenetic variability confounds transcriptome but not proteome profiling for coexpression-based gene function prediction. Mol. Cell. Proteom. MCP 2018, 17, 2082–2090. [Google Scholar] [CrossRef] [Green Version]
- Braun, A.; Aszodi, A.; Hellebrand, H.; Berna, A.; Fassler, R.; Brandau, O. Genomic organization of profilin-III and evidence for a transcript expressed exclusively in testis. Gene 2002, 283, 219–225. [Google Scholar] [CrossRef]
- Umer, N.; Arévalo, L.; Phadke, S.; Lohanadan, K.; Kirfel, G.; Sons, D.; Sofia, D.; Witke, W.; Schorle, H. Loss of profilin3 impairs spermiogenesis by affecting acrosome biogenesis, autophagy, manchette development and mitochondrial organization. Front. Cell Dev. Biol. 2021, 9, 749559. [Google Scholar] [CrossRef] [PubMed]
- Huelseweh, B.; Kohl, B.; Hentschel, H.; Kinne, R.K.; Werner, A. Translated anti-sense product of the Na/phosphate co-transporter (NaPi-II). Biochem. J. 1998, 332, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, A.; Schmutzler, G.; Carlile, M.; Miles, C.G.; Peters, H. Expression profiling of antisense transcripts on DNA arrays. Physiol. Genom. 2007, 28, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalbant, P.; Boehmer, C.; Dehmelt, L.; Wehner, F.; Werner, A. Functional characterization of a Na+-phosphate cotransporter (NaPi-II) from zebrafish and identification of related transcripts. J. Physiol. 1999, 520, 79–89. [Google Scholar] [CrossRef]
- Soumillon, M.; Necsulea, A.; Weier, M.; Brawand, D.; Zhang, X.; Gu, H.; Barthes, P.; Kokkinaki, M.; Nef, S.; Gnirke, A.; et al. Cellular source and mechanisms of high transcriptome complexity in the Mammalian testis. Cell Rep. 2013, 3, 2179–2190. [Google Scholar] [CrossRef]
- Piatek, M.J.; Henderson, V.; Fearn, A.; Chaudhry, B.; Werner, A. Ectopically expressed Slc34a2a sense-antisense transcripts cause a cerebellar phenotype in zebrafish embryos depending on RNA complementarity and Dicer. PLoS ONE 2017, 12, e0178219. [Google Scholar] [CrossRef]
- Chen, J.; Gao, C.; Zhu, W. Long non-coding RNA SLC25A25-AS1 exhibits oncogenic roles in non-small cell lung cancer by regulating the microRNA-195-5p/ITGA2 axis. Oncol. Lett. 2021, 22, 529. [Google Scholar] [CrossRef]
- Li, J.; Fan, S.; Liu, S.; Yang, G.; Jin, Q.; Xiao, Z. LncRNA NOP14-AS1 Promotes tongue squamous cell carcinoma progression by targeting microRNA-665/HMGB3 axis. Cancer Manag. Res. 2021, 13, 2821–2834. [Google Scholar] [CrossRef]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Lavarone, E.; Barbieri, C.M.; Pasini, D. Dissecting the role of H3K27 acetylation and methylation in PRC2 mediated control of cellular identity. Nat. Commun. 2019, 10, 1679. [Google Scholar] [CrossRef]
- Barman, P.; Reddy, D.; Bhaumik, S.R. Mechanisms of antisense transcription initiation with implications in gene expression, genomic integrity and disease pathogenesis. Non Cod. RNA 2019, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowthaman, U.; García-Pichardo, D.; Jin, Y.; Schwarz, I.; Marquardt, S. DNA processing in the context of noncoding transcription. Trends Biochem. Sci. 2020, 45, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Dutta, B.; Hee, Y.T.; Chng, W.J. Towards understanding of PRC2 binding to RNA. RNA Biol. 2019, 16, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ohsumi, T.K.; Kung, J.T.; Ogawa, Y.; Grau, D.J.; Sarma, K.; Song, J.J.; Kingston, R.E.; Borowsky, M.; Lee, J.T. Genome-wide identification of polycomb-associated RNAs by RIP-seq. Mol. Cell 2010, 40, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillay, S.; Takahashi, H.; Carninci, P.; Kanhere, A. Antisense RNAs during early vertebrate development are divided in groups with distinct features. Genom. Res. 2021, 31, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Piatek, M.J.; Mattick, J.S. Transpositional shuffling and quality control in male germ cells to enhance evolution of complex organisms. Ann. N. Y. Acad. Sci. 2015, 1341, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, P.; Chung, G.; Pye, K.; Yukawa, T.; Imanishi, A.; Takai, Y.; Brown, C.; Wagoner, M.P. Freshly isolated primary human proximal tubule cells as an in vitro model for the detection of renal tubular toxicity. Toxicology 2020, 442, 152535. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Markovich, D. Expression cloning and radiotracer uptakes in Xenopus laevis oocytes. Nat. Protoc. 2008, 3, 1975–1980. [Google Scholar] [CrossRef]
- Fearn, A.; Allison, B.; Rice, S.J.; Edwards, N.; Halbritter, J.; Bourgeois, S.; Pastor-Arroyo, E.M.; Hildebrandt, F.; Tasic, V.; Wagner, C.A.; et al. Clinical, biochemical, and pathophysiological analysis of SLC34A1 mutations. Physiol. Rep. 2018, 6, e13715. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zinad, H.S.; Sae-Lee, C.; Ariza-Mateos, M.A.; Adamson, G.; Khazeem, M.M.; Knox, A.; Chung, G.; Mann, J.; Werner, A. Interdependent Transcription of a Natural Sense/Antisense Transcripts Pair (SLC34A1/PFN3). Non-Coding RNA 2022, 8, 19. https://doi.org/10.3390/ncrna8010019
Zinad HS, Sae-Lee C, Ariza-Mateos MA, Adamson G, Khazeem MM, Knox A, Chung G, Mann J, Werner A. Interdependent Transcription of a Natural Sense/Antisense Transcripts Pair (SLC34A1/PFN3). Non-Coding RNA. 2022; 8(1):19. https://doi.org/10.3390/ncrna8010019
Chicago/Turabian StyleZinad, Hany S., Chanachai Sae-Lee, Maria Ascensión Ariza-Mateos, Grace Adamson, Mushtaq Mufleh Khazeem, Amber Knox, Git Chung, Jelena Mann, and Andreas Werner. 2022. "Interdependent Transcription of a Natural Sense/Antisense Transcripts Pair (SLC34A1/PFN3)" Non-Coding RNA 8, no. 1: 19. https://doi.org/10.3390/ncrna8010019
APA StyleZinad, H. S., Sae-Lee, C., Ariza-Mateos, M. A., Adamson, G., Khazeem, M. M., Knox, A., Chung, G., Mann, J., & Werner, A. (2022). Interdependent Transcription of a Natural Sense/Antisense Transcripts Pair (SLC34A1/PFN3). Non-Coding RNA, 8(1), 19. https://doi.org/10.3390/ncrna8010019