2.1. The Idea of Interaction of Biosphere and Geologic Processes. The Model of the Global Cycle of Biosphere Carbon and Its Development
As early as 1926, the famous Russian naturalist Vladimir Vernadsky put forward an idea on the interaction and interconditionality of the biosphere and geological processes. Developing and concretizing his idea, we have suggested a model of the global cycle of biosphere carbon [
1,
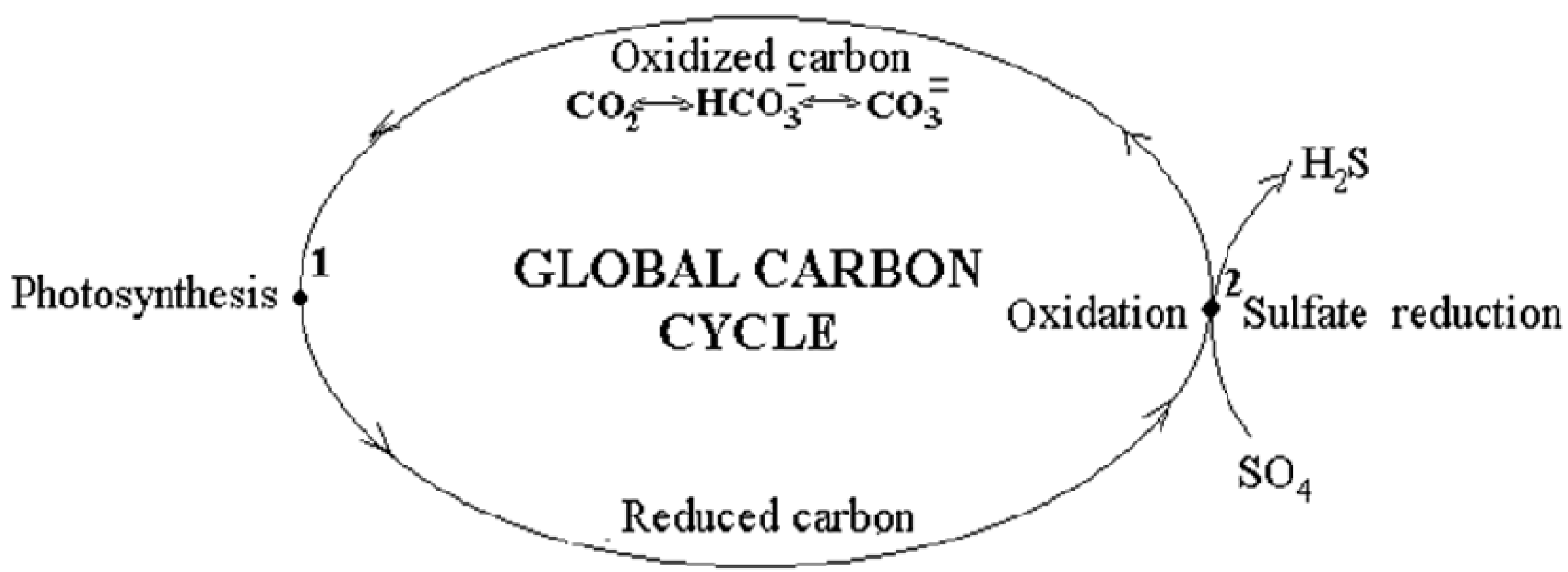
2]. According to the proposed model, carbon transformations occur along a closed loop. In the course of it, the completely oxidized state in the natural “carbon dioxide—bicarbonate—carbonate “ system transforms into a reduced state that arises during photosynthesis and the subsequent transformations, including those occurring in the earth’s crust after the burial of the “living” matter and its transition into sedimentary organic forms. The reverse transition into the initial completely oxidized state carbon makes in the subduction zone, where sedimentary organic matter is oxidized to CO
2, thus ending the redox transformations in the loop (
Figure 1).
The energy for the carbon transition into the reduced state in photosynthesis is provided by the sun. The energy for the carbon oxidation in reverse transition is taken from the energy evolved in collisions of the oceanic lithosphere plates with the continental plate, on which organic matter is deposited. The collisions of lithosphere plates occur in the subduction zone. It is important to stress that the cycle is called biosphere one since it is assumed that the carbon of carbonate thicknesses does not participate in the global carbon cycle because of its extraordinarily low rate in the reactions of chemical exchange as compared with the rates of other components of the natural “carbon dioxide-bicarbonate-carbonate” system [
1].
The carbon cycle can be formally considered as a virtual natural self-regulated machine, which finally provides the renewable synthesis of biomass in the course of the evolution. The machine consists of geological and biosphere parts. The geological part comprises the permanently moving lithosphere plates performing irregular motion, which, according to the hypothesis, occur due to the gravitational interaction of celestial bodies with the Earth. Two phases are distinguished in motion [
1,
2]. The short-term phase of relatively quick motion accompanied by frequent collisions is called an orogenic period. It is characterized by intense volcanism, magmatism, and mountain building. The long-term phase of slow motion, accompanied by rare collisions, occurs in the quiet state of the Earth’s crust when flattening and weathering become dominant. This phase is called the geosynclinal period.
The interaction between celestial bodies is one of the theories, and not the only one; in fact, there is also a theory of internal terrestrial processes. However, in any case, this origin is not essential from the point of view of the discussion and conclusions set out in the text.
The geological part is a leading one since it determines the duration of both periods of the geological part. The impact of the processes in the geological part of the model on its biosphere part is transmitted via the injections of CO2, which arise during the oxidation of sedimentary organic matter in the reaction of thermochemical sulfate reduction occurring in the subduction zone. The subsequent CO2 distribution over the planet initiates photosynthesis and coupled biosphere processes. In the orogenic period, while oxidation of organic matter goes on, the CO2 concentration in “atmosphere—hydrosphere” system is maximal, providing a maximal photosynthesis rate.
In the geosynclinal period, when the number and the intensity of collisions of lithosphere plates decrease, it causes a sharp decline in the intensity of sulfate reduction and slowing of CO2 flux into the “atmosphere—hydrosphere” system. The concentration of CO2 in the system drops, causing the corresponding reduction of the photosynthesis rate. It leads to the conclusion that in the geosynclinal period, photosynthesis functions under CO2 pool depletion.
Due to repetitive orogenic cycles, the above means that evolution of photosynthesis is not continuous, but occurs in the form of pulsations, caused by the filling of the Earth’s “atmosphere-hydrosphere” system with CO2 during the orogenic period of the cycle and its depletion during the geosynclinal period.
Before considering the operation of the virtual machine under study, it is necessary to understand how global photosynthesis works in large systems and how it differs from conventional photosynthesis in an “environment—plant leaf” system.
2.2. Global Photosynthesis in Large Systems: Its Description and Properties
As follows from the above, each orogenic cycle in a geologic part of the virtual machine generates a photosynthesizing cycle in its biosphere part. Photosynthesis in large systems (global photosynthesis), unlike photosynthesis of an individual organism of C3 type (conventional photosynthesis), represents generalized characteristics of the photosynthetic activity of all organisms living at the moment and has all the properties of photosynthesis of an individual organism, excepting the ability to ontogenesis [
3]. They include the presence of CO
2 assimilation and photorespiration. The first process is reciprocal to the second. The CO
2 assimilation is responsible for the accumulation of organic matter (biomass) in the sediment, whereas the photorespiration is responsible for its decrease.
Like conventional photosynthesis, global one has the ability to fractionate carbon isotopes. An increase in the CO
2/O
2 concentration ratio in the environment stimulates CO
2 assimilation and, hence, the organic matter accumulation in sediment, while the reduction in the CO
2/O
2 concentration ratio stimulates photorespiration and leads to a decrease in the accumulation. Accordingly, the increase in CO
2 concentration in the orogenic period of the cycle results in the
12C enrichment of organic matter, while the increase of oxygen concentration in the course of the geosynclynal period results in the gradual enrichment of organic matter in
13C. The organic matter most enriched in
12C is observed in the orogenic period of the cycle, whereas the maximal
13C enrichment is achieved by the end of the geosynclinal period. Formally, the photosynthesis of an individual organism is usually described by Equation (1).
Here, the left side of the equation shows the carbon dioxide and the water necessary for photosynthesis; the source of them is the environment; hv denotes the energy of the sun; (CH2O) is the product of photosynthesis, formally denoting the biomass produced during photosynthesis; O2 denotes oxygen, the second product of photosynthesis.
It is shown [
3] that photosynthesis in large systems, under certain assumptions, can be described by a similar equation. Let us see first what is understood as biomass for the biosphere. To do this, we will use the concept of “living” substance, introduced by Vladimir Vernadsky [
4]. It is obvious that biomass consists of autotrophic (photosynthetic) and heterotrophic parts.
Both parts of the “living” matter can be considered photosynthesis products if the heterotrophic part is taken as a secondary product obtained by using photosynthetic biomass in the food chain. At the same time, oxygen released during the formation of the primary product, before its part was used for the synthesis of heterotrophic biomass, accumulates in the atmosphere.
Thus, the equation of global photosynthesis for the biosphere can be rewritten [
3] as follows:
Similarly, Equation (3) of global photosynthesis can be written for the system of the global cycle of biosphere carbon as Equation (4):
The difference between Equations (3) and (4) is in the fact that in the right part of (4), as an analog of biomass, the “living” matter appears to be the sum of “living” matter and sedimentary organic matter. Obviously, the first term is much less as compared with the second and can be neglected. Atmospheric oxygen will also include the oxygen that replenishes the atmosphere at all stages of photosynthesis of both primary and secondary (heterotrophic) products over the same period.
Given the above approximations, Equation (4) presenting photosynthesis for the global carbon cycle system can be shown as follows
The validity of using Equation (5) for a description of photosynthesis in the global carbon cycle is proved by the fact that the parameters of global photosynthesis, operating in the global carbon cycle, analogs of the parameters of conventional photosynthesis in the Equation (1), i.e., the relationship between substrate and product (CO2 and O2) of photosynthesis is anti-phase, while the relationship between photosynthesis products (CH2O and O2) is in-phase.
Indeed,
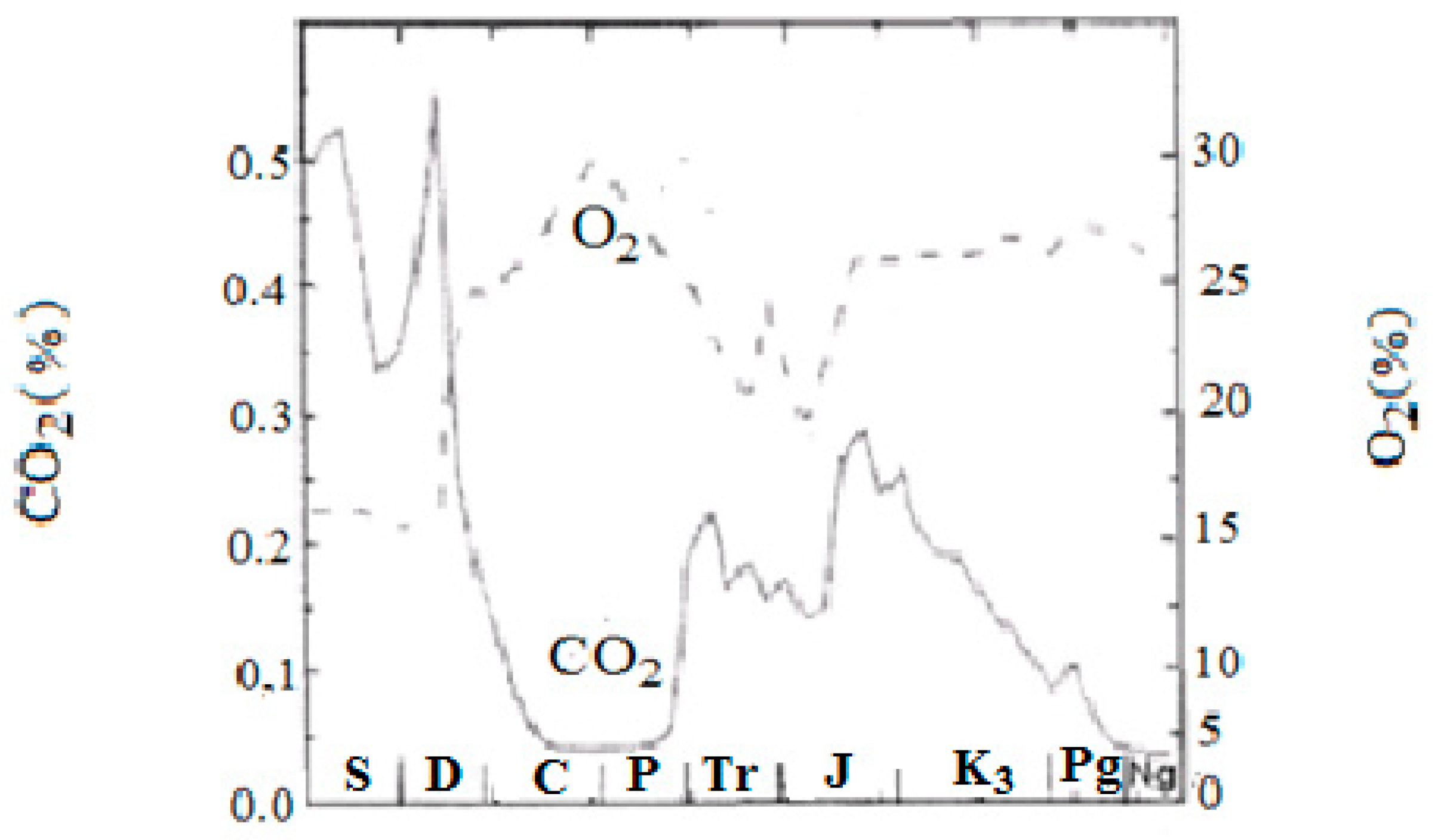
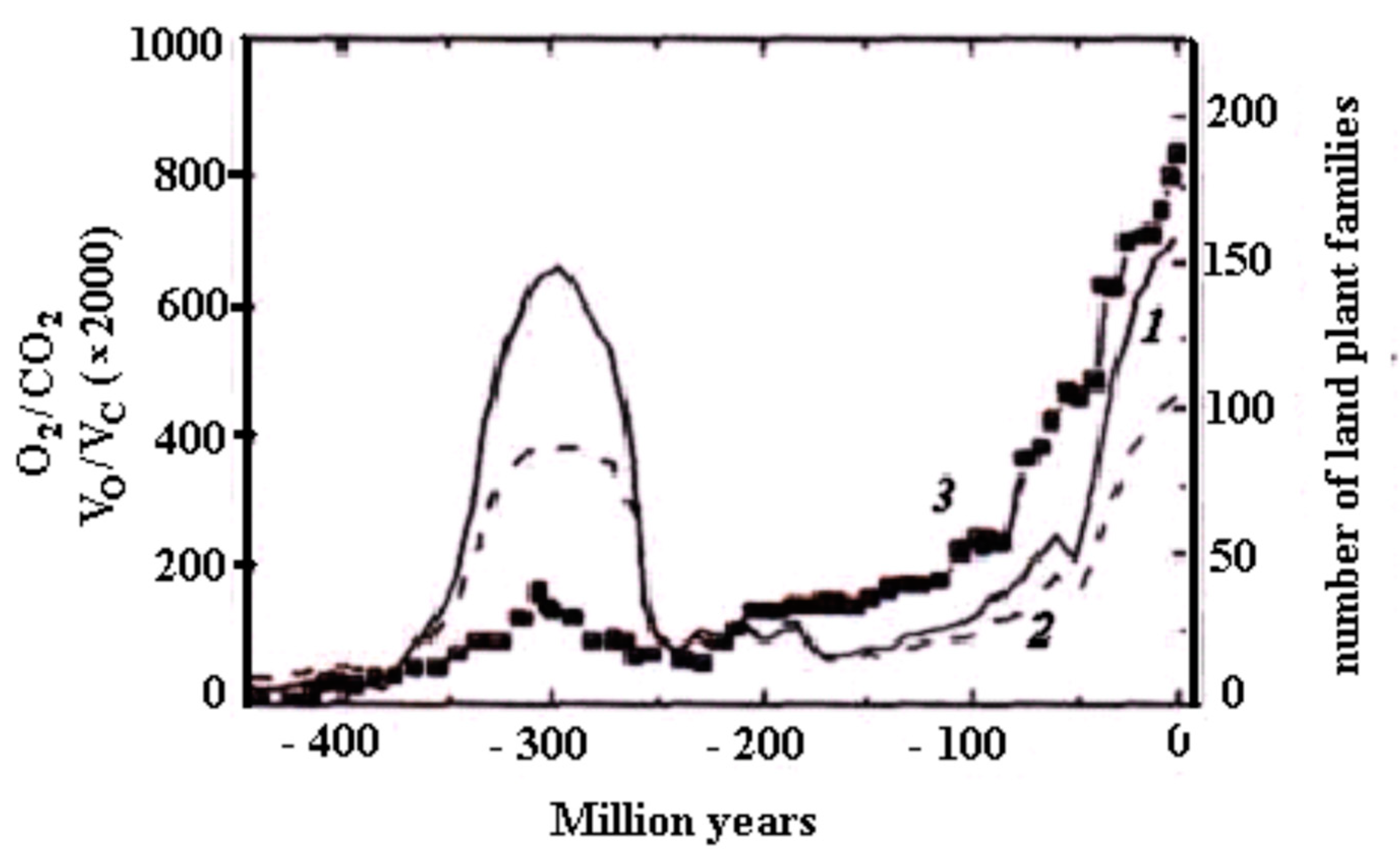
Figure 2 demonstrates the anti-phase relationship between CO
2 and O
2 in the Phanerozoic, calculated by means of the Geocarb III model.
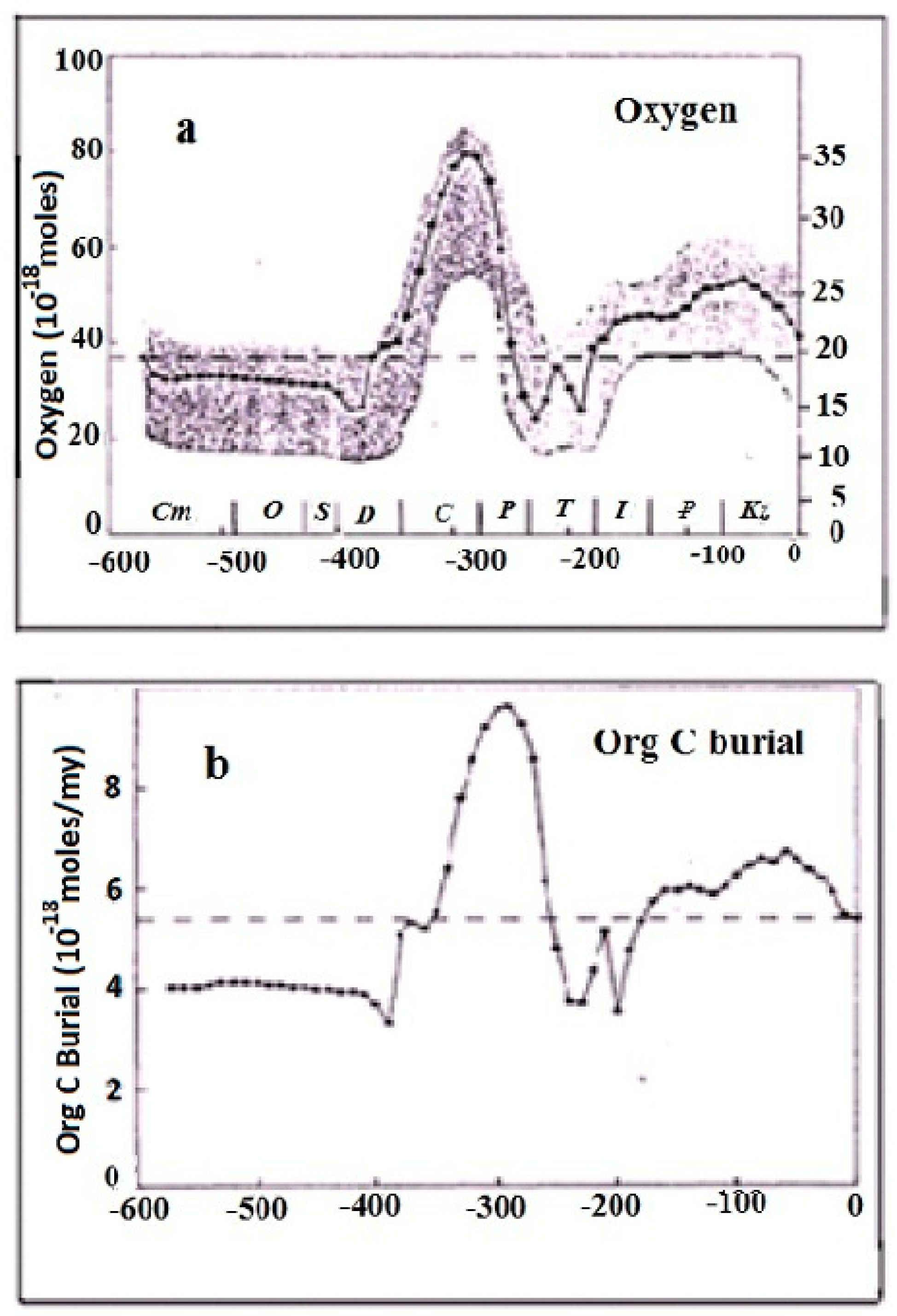
Figure 3 demonstrates the in-phase relationship between oxygen content in the atmosphere and sedimentation rate in the Phanerozoic.
2.3. The Influence of Geologic Processes on Global Photosynthesis Is Carried out via the Oxidation of Sedimentary Organic Matter in the Subduction Zone: The Ecological Compensation Point
Having found out the meaning of the term “global photosynthesis”, it is time to proceed considering the work of the biosphere part of the natural virtual machine, where global photosynthesis plays a key role. The analysis of the biosphere part functioning begins with the reaction of sedimentary organic matter oxidation in the subduction zone. It can be presented as follows:
The term designated as (CH2O) means organic matter accumulated on the continental plate during all preceding orogenic cycles. Organic matter acts as a reducing agent, and the bound oxygen of gypsum (SO4 =), formed from the sulfate of marine waters, acts as an oxidizer. The carbon dioxide evolved in the reaction is injected into the atmosphere and carried over the planet, initiating photosynthesis. Another reaction product, hydrogen sulfide (S=) also rises to the surface, but being more reactive than CO2, binds with oxygen, accumulated in the preceding geosynclinal period decreasing its concentration in the atmosphere. Oxygen is also bound by the reduced forms of metals from igneous rocks that rise to the Earth’s surface with volcanic exhalations. It should be emphasized that the reaction under study is the main point of coupling geological processes with biosphere ones.
In the present work, the subduction is considered only between the oceanic and continental lithospheric plates since the sedimentary organic matter is accumulated mainly on the continental plates, which is further oxidized.
As said before, the biosphere part of the natural virtual machine is under the guidance of the geological part since the lithospheric plates’ movement determines the duration of the orogenic and geosynclynal periods of the cycle. At the same time, variations in global photosynthesis within the orogenic cycle cause climatic and associated changes in the biosphere.
As noted earlier, the properties of global photosynthesis, as well as the properties of conventional photosynthesis, depend on the CO2/O2 ratio in the environment. The ratio changes in the course of the orogenic cycle. In the orogenic period of the cycle, when the sulfate reduction proceeds, “the atmosphere—hydrosphere” system is filled with CO2. Its concentration achieves maximum. Taking into account the shortness of the orogenic period, as compared with the geosynclinal one, we have accepted that the concentration of CO2 in the orogenic period is maximal and constant.
In the geosynclinal period, the intensity of the sulfate reduction reaction slows down sharply, causing a considerable reduction of CO2 flux from the subduction zone. At the same time, thanks to photosynthesis, the CO2 consumption goes on. It allows concluding that global photosynthesis within the geosynclinal period functions under the condition of CO2 pool depletion achieving minimal CO2 concentration by the end of the period. It means that photosynthesis works in a periodic mode.
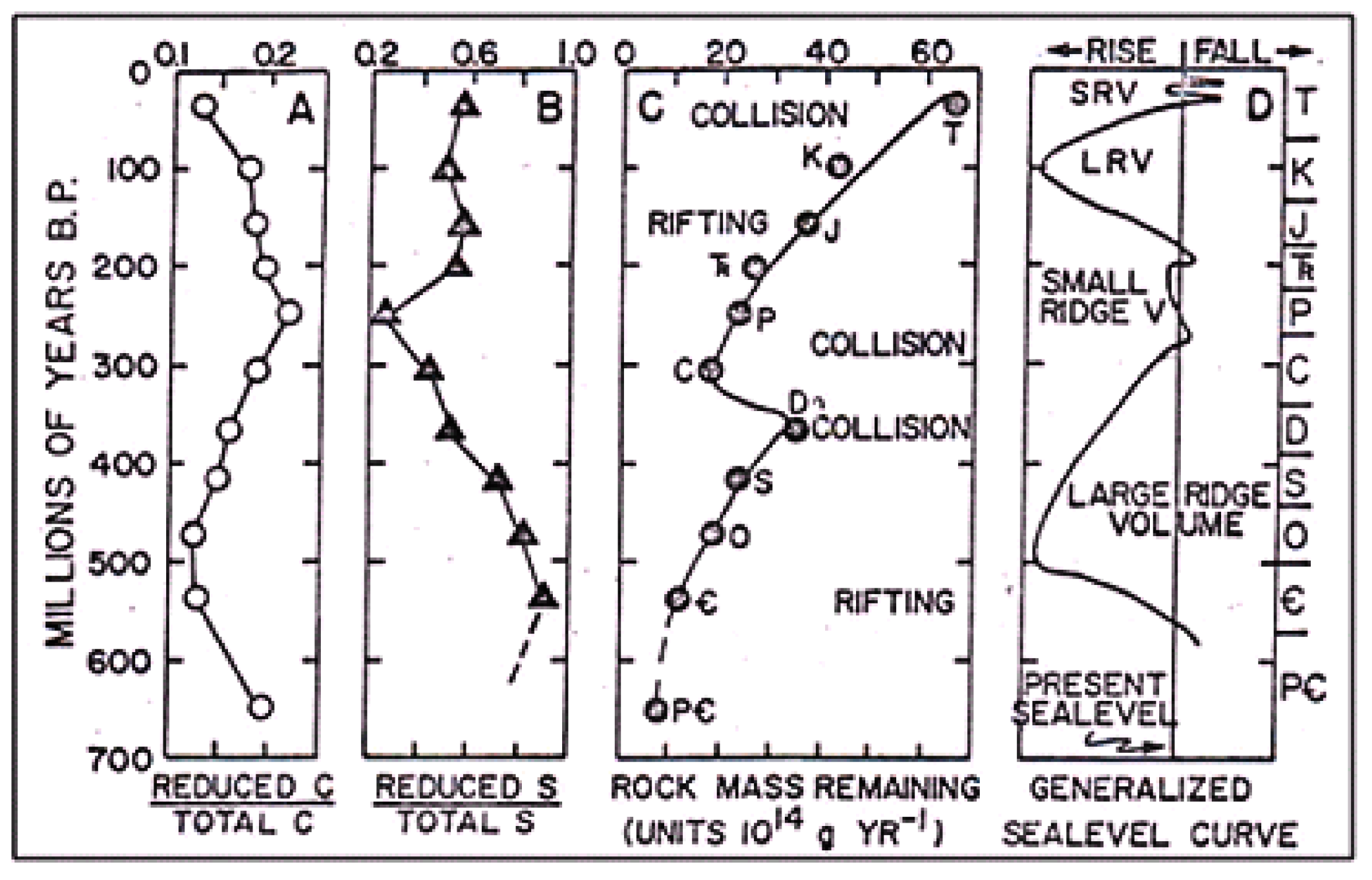
To answer the question of how the concentration of O
2 changes during the repetitive orogenic cycles (in the course of geological time), let us turn to
Table 1. It is easy to see that with the emergence of photosynthesis, the oxygen concentration first increases rapidly, reaching the maximum values in the middle of the Phanerozoic (Carboniferous/Permian period). Then, it decreases slightly. Taking into account the present oxygen concentration in the atmosphere (~21%), it can be assumed that the oxygen concentration strives to stationary values over time.
Accordingly, the accumulation of organic matter in the Earth’s crust began to decrease, which occurred synchronously with an increase in the oxygen content in the atmosphere. As already mentioned, the predominance of the assimilation component in global photosynthesis entails the enrichment of carbon of organic matter with the isotope
12C, while the predominance of the photorespiratory component leads to the enrichment of organic matter with the isotope
13C. The described mechanism of accumulation of sedimentary organic matter in the Earth’s crust and oxygen in the atmosphere is reflected in the observed isotopic enrichment of carbon of sedimentary organic matter in isotope
13C with time [
15].
In the course of repetitive orogenic cycles, with each new cycle, the photorespiration contribution has increased, while the contribution of CO2 assimilation decreased. This meant that the increment of sedimentary organic matter (the newly formed organic matter) in the Earth’s crust was decreasing, but the total amount of organic matter has increased, though at a lesser rate. In the same way, the increment of O2 has decreased, but the total content of oxygen in the atmosphere has increased at a lesser rate.
This situation continued until the contribution of CO2 assimilation became equal to the contribution of photorespiration. Simultaneously, the system of the global carbon cycle came to a stationary state. It meant the system had reached an ecological compensation point (like a state of a compensation point in conventional photosynthesis).
Resuming the obtained results of the consideration of the biosphere part of the natural virtual machine (the global cycle of biosphere carbon), it can be argued that global photosynthesis developed from cycle to cycle in the form of pulsations. More exactly, pulsations have appeared with the advent of oxygen in the atmosphere and coupled photorespiration. In other words, global photosynthesis in the long-term geosynclinal period of the cycle evolved in conditions of CO2 pool depletion. This allowed concluding on the emergence of the isotope Rayleigh effect, which conditions carbon isotope fractionation within the geosynclinal period of the cycle and determines the isotope composition of organic and carbonate carbon in the cycle. In particular, the produced sedimentary organic matter turned out to be the most enriched in 12C in the orogenic period, whereas in the geosynclinal period, it was getting enriched with 13C, achieving maximal values by the end of the period.
In the course of repetitive orogenic cycles, the described evolution of global photosynthesis eventually leads to a stationary state, when the amount of organic carbon produced during photosynthesis becomes equal to the amount of carbon returned back to the oxidized state. From this moment, the accumulation of organic matter in the Earth’s crust and oxygen in the atmosphere no longer occurs.
Thus, the evolution of global photosynthesis during repetitive cycles ensures the achievement of stationary levels of oxygen concentrations in the atmosphere close to the maximum. Sedimentary organic matter in each new cycle differs in its composition from the composition synthesized in the previous cycle. This is caused by a higher level of oxidation in the environment and a higher level of evolution. The latter means that changes in the composition of “living matter” are associated with adaptive changes in the biomass of living organisms to a higher level of oxygen concentration, increasing with time.
2.4. The Coupled Changes of Global Photosynthesis and Environmental Parameters within the Orogenic Cycle
As shown in the previous section, the evolution of global photosynthesis in the biosphere part of the virtual machine begins with the arrival of CO2 as a result of thermochemical sulfate reduction, where sedimentary organic matter is oxidized in the subduction zone. The rate of global photosynthesis depends on the concentration of the reaction substrate (CO2); the concentration of another substrate, H2O (before the appearance of terrestrial life), is considered to be excessive, i.e., not rate-limiting. Since oxygen (O2) is one of the products of global photosynthesis, it is convenient to determine the evolution of global photosynthesis in the orogenic cycle via the variations of the CO2/O2 ratio. It varies in the cycle from the maximum value at the beginning of the cycle (in the orogenic period) to the minimum value at its end (at the end of the geosynclinal period).
Let us see now the coupled variations of the global photosynthesis evolution and environmental and life parameters within the orogenic cycle.
In the orogenic period of the cycle, when the oxidation of sedimentary organic matter in the subduction zone intensively occurs, the CO2 concentration in the “atmosphere–hydrosphere” system on the Earth reaches its maximum. Taking into account the “greenhouse effect”, it is obvious that surface temperatures on the Earth in this period achieve their maximum values.
In the geosynclinal period, the sulfate reduction in the subduction zone slows down, whereas global photosynthesis goes on. Hence, the CO2 concentration begins to decrease. By the end of the geosynclinal period, the lowest temperature is reached, and glaciations occur. In parallel with the temperature decline due to photosynthesis, the oxygen concentration in the atmosphere increases. It means the CO2/O2 concentration ratio in the “atmosphere–hydrosphere” system declines in parallel with CO2 concentration. Accordingly, the CO2/O2 ratio achieves its minimum at the end of the geosynclinal period. Thus, one can see that all environmental characteristics vary within the orogenic cycle simultaneously with photosynthesis variations.
Thus, in the biosphere part of the virtual machine, the evolution of global photosynthesis within a separate orogenic cycle is accompanied by simultaneous and coupled variations of environmental parameters: temperature, oxygen and carbon dioxide content, and the CO2/O2 ratio. In addition to the aforementioned environmental parameters, so-called life parameters change, which are dependent on environmental parameters.
Gorikan and co-authors [
16] have investigated radiolarians, which are single-celled organisms with a wide range of adaptive capabilities. Among them, there were species that dominated in an anaerobic environment and at elevated temperatures, i.e., under conditions typical to the orogenic period. The other species dominated in an oxygen environment and at low temperatures, i.e., dominated in conditions that corresponded to the geosynclinal period of the cycle. The studied group of radiolarians included 60 genera, including 157 species.
As a model of the orogenic cycle, the authors considered a geological time interval in the Jurassic Pliensbachian—Toarcian, in which climatic oscillations have occurred, one phase of which (Pliensbachian) was analogous to the geosynclinal period of the orogenic cycle, the other (Toarcian) was analogous to the orogenic period of the cycle.
The dominance of thermophilic anaerobes in the “orogenic” period was replaced by the dominance of cold-resistant anaerobes by the end of the “geosynclinal” period. This corresponded well to the changes in temperature and CO2 concentration expected from the model—from a maximum in the orogenic period to a minimum by the end of the geosynclinal period. Oxygen concentration changes occurred in the opposite direction from the minimum O2 concentration in the orogenic period to the maximum by the end of the geosynclinal period.
Thus, the data obtained by Gorican and co-authors (16) confirm the connection between the evolution of “living” organisms and, hence, the chemical composition of sedimentary organic matter with changes in environmental conditions.
One more life parameter bound to the changes in CO2/O2 concentration ratio within the orogenic cycle is the carbon isotope ratio of sedimentary organic matter. As said before, the carbon isotope ratio of organic matter depends on the contribution of CO2 assimilation and photorespiration to global photosynthesis.
The greater the contribution of CO2 assimilation to global photosynthesis, the more sedimentary organic matter is enriched in 12C. On the contrary, the greater the contribution of photorespiration, the more the enrichment of organic matter in 13C. An indicator of the contribution of the photosynthesis evolution is the CO2/O2 ratio, the change of which, in the cycle, was discussed above as it follows from the previous consideration, the most “light “ carbon isotope composition organic matter has in the orogenic period. In the geosynclinal period, when the CO2/O2 ratio begins to decrease because of the cessation of the influx of CO2 from the subduction zone, the organic matter is enriched in 13C.
Hayes [
17] has suggested using another life parameter based on carbon isotope composition. It is an analog of modern
13C discrimination used in conventional photosynthesis. The parameter defines the difference between the carbon isotope composition of synthesized biomass and that of CO
2. Based on the actualism principle, he proposed to express
13C discrimination in the past, as follows:
where Δ
13C discrimination in the past is the difference between the carbon isotope composition of sedimentary organic matter and that of carbonates coeval to this organic matter; δ
13C
org is the carbon isotope ratio of sedimentary organic matter, which is the analog of biomass; δ
13C
carb is carbon isotope ratio of carbonates, coeval to organic matter. It is an analog of CO
2.
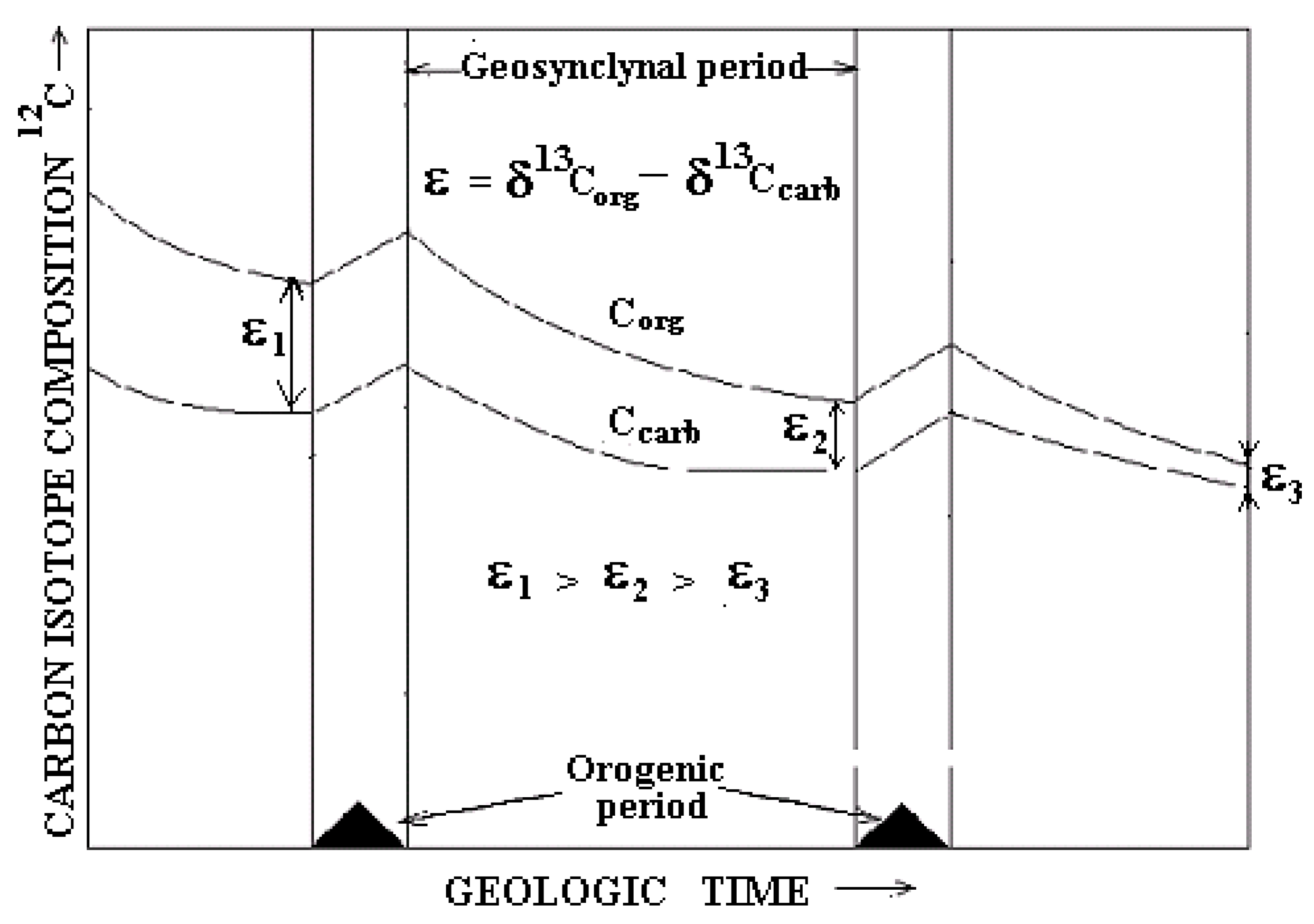
How does Δ 13C discrimination vary within the orogenic cycle? Since the CO2 concentration in the geosynclinal period of the cycle decreases, then under the action of Rayleigh isotope depletion effect, the δ13Corg and δ 13Ccarb values become more positive. The increase of O2 content within the orogenic cycle due to photosynthesis makes additional enrichment of organic matter in 13C. The value δ13Corg becomes more positive. As a result, the value Δ13C becomes lesser by the end of the cycle.
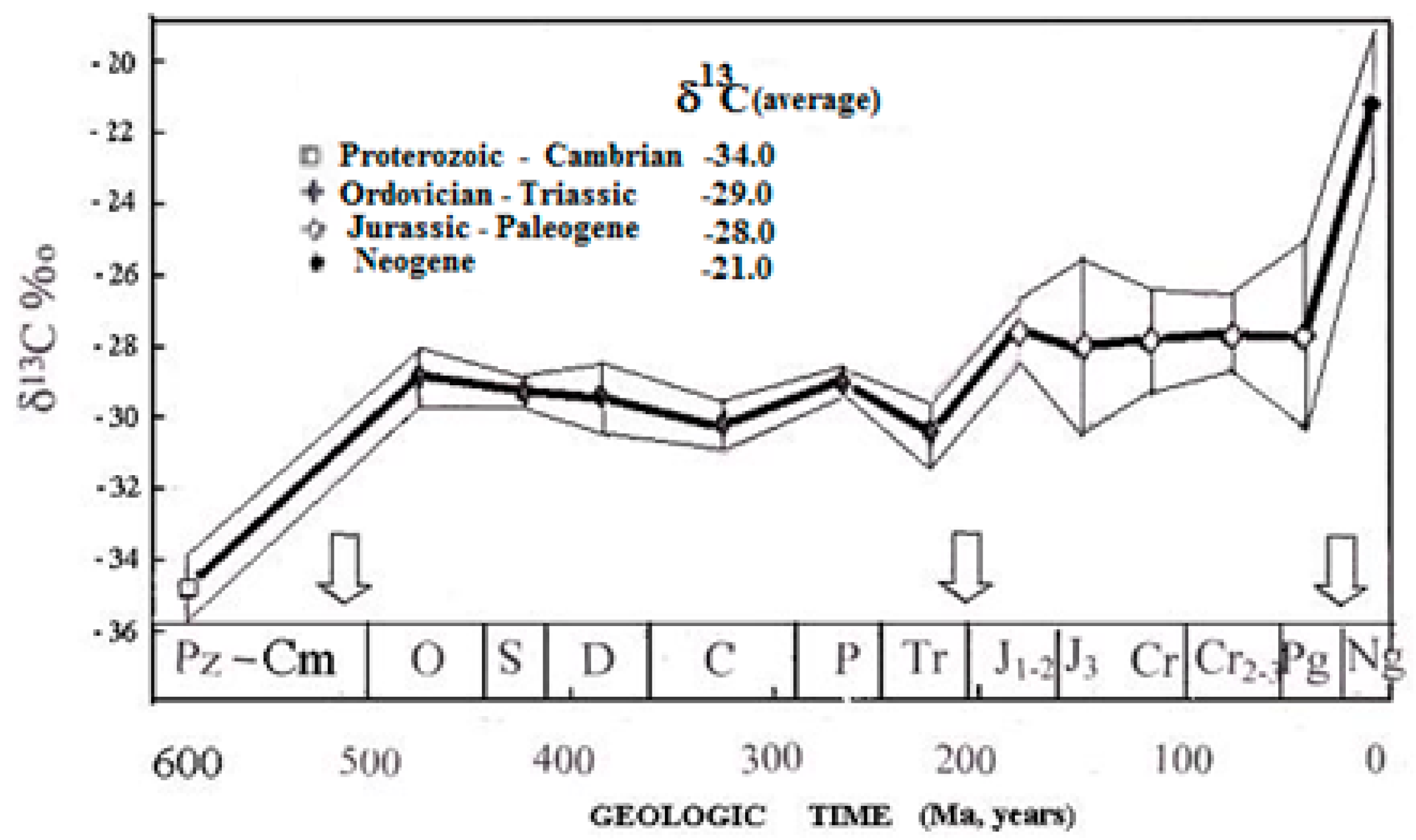
Considering the trend of Δ
13C change in repetitive orogenic cycles and taking into account the progressive increase of oxygen content in the atmosphere, one can conclude that
13C isotope discrimination in the past should decrease step by step, as shown in
Figure 4; this trend is supported by the observed enrichment in
13C of sedimentary organic matter with time [
15].
In conclusion, I can say that the considered material allows asserting that in spite of variations of global photosynthesis within the orogenic cycle, the general trend with time shows that the evolution of global photosynthesis went on in the pulsation regime step by step.
This happened until the evolution of photosynthesis brought the global carbon cycle system to the point of ecological compensation. As shown in the work [
1], this took place in the Miocene, when a new C4 type of CO
2 assimilation appeared. The biological sense of it is in the fact that such evolution helped to select and to consolidate useful properties for the adaptation of organisms to varying conditions.
Of course, the proposed model is just an approximation of the description of the real situation. However, in the following part of the paper, we will show that it is a good approximation.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}