The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995

 ,
,
Abstract
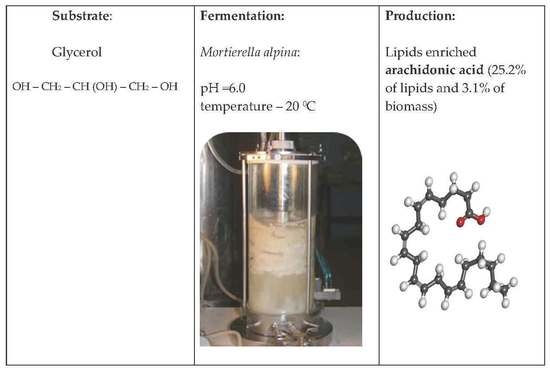
:
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Chemicals
2.2. Media and Cultivation Conditions
2.3. Assays
2.4. Calculations
3. Results and Discussion
3.1. The Effect of pH
3.2. The Effect of Temperature
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for cell single oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Ward, O.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Process Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Dyal, S.D.; Narine, S.S. Implications for the use of Mortierella fungi in the industrial production of essential fatty acids. Food Res. Int. 2005, 38, 445–467. [Google Scholar] [CrossRef]
- Dedyukhina, E.G.; Kamzolova, S.V.; Vainshtein, M.B. Arachidonic acid as an elicitor of the plant defense response to phytopathogens. Chem. Biol. Technol. Agric. 2014, 1, 18. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Dedyukhina, E.G.; Chistyakova, T.I.; Lunina, J.N.; Mironov, A.A.; Stepanova, N.N.; Shemshura, O.N.; Vainshtein, M.B. Application of organic acids for plant protection against phytopathogens. Appl. Microbiol. Biotechnol. 2017, 101, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, K.; Higashiyama, K.; Shimizu, A. Production of Arachidonic Acid-Containing Lipid and Dihomo-Gamma-Linolenic Acid-Containing Lipid. US Patent 2005/0266538A1, 1 December 2005. [Google Scholar]
- Barclay, W.R. Method for Production of Arachidonic Acid. Patent EP1726217 (A1), 29 November 2006. [Google Scholar]
- Barclay, W.R. Method for Production of Arachidonic Acid. US Patent 7,601,523,132, 13 October 2009. [Google Scholar]
- Higashiyama, K. Method for Polyunsaturated Fatty Acid Production Using Novel Cell Preservation Technique. Patent WO2006016702 (A1), 16 February 2006. [Google Scholar]
- Streekstra, H.; Brocken, P.J.M. Preparation of Microbial Oil. US Patent 7470527B2, 30 December 2008. [Google Scholar]
- Streekstra, H.; Brocken, P.J.M. Preparation of Microbial Oil. US Patent 20090053342 (A1), 26 February 2009. [Google Scholar]
- Li, X.; Lin, Y.; Chang, M.; Jin, Q.; Wang, X. Efficient production of arachidonic acid by Mortierella alpina through integrating fed-batch culture with a two-stage pH control strategy. Bioresour. Technol. 2015, 181, 275–382. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.J.; Wang, C.; Zheng, Z.Y.; Zhu, L.; Zhan, X.B.; Lin, C.C. Improving arachidonic acid fermentation by Mortierella alpina through multistage temperature and aeration rate control in bioreactor. Prep. Biochem. Biotechnol. 2016, 46, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Pradima, J.; Rajeswari Kulkarni, M.; Archna. Review on enzymatic synthesis of value added products of glycerol, a by-product derived from biodiesel production. Resour. Effic. Technol. 2017, 3, 394–405. [Google Scholar] [CrossRef]
- Nicol, R.W.; Marchand, K.; Lubitz, W.D. Bioconversion of crude glycerol by fungi. Appl. Microbiol. Biotechnol. 2012, 93, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Fakas, S.; Fick, M.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M.; Marc, I.; Aggelis, G. Biotechnological valorization of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process: Production of 1,3-propandiol, citric acid and single cell oil. Biomass Bioeng. 2008, 32, 60–71. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; Huijberts, G.N.M.; Eggink, G. High-cell-density cultivation of the lipid accumulating yeast Cryptococcus curvatus using glycerol as a carbon source. Appl. Microbiol. Biotechnol. 1996, 45, 575–579. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; van der Wal, H.; Weusthuis, R.; Eggink, G. Cultivation of the oleaginous yeast Cryptococcus curvatus in a new reactor with improved mixing and mass transfer characteristics (Surer®). Biotechnol. Techol. 1996, 10, 277–282. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipid production by Yarrowia lipolytica growing on industrial glycerol in a single-stage continuous culture. Bioresour. Technol. 2002, 82, 43–49. [Google Scholar] [CrossRef]
- Dedyukhina, E.G.; Chistyakova, T.I.; Kamzolova, S.V.; Vinter, M.V.; Vainshtein, M.B. Arachidonic acid synthesis by glycerol-grown Mortierella alpina. Eur. J. Lpid Sci. Technol. 2012, 114, 833–841. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Rontou, M.; Belka, A.; Athenaki, M.; Gardeli, C.; Mallouchos, A.; Kalantzi, O.; Koutinas, A.A.; Kookos, I.K.; Zeng, A.-P.; et al. Conversion of biodiesel-derived glycerol into biotechnological products of industrial significance by yeast and fungal strains. Eng. Life Sci. 2017, 17, 262–281. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Perevoznikova, O.A.; Shishkanova, N.V.; Finogenova, T.V. Pyruvic acid production by a thiamine auxotroph of Yarrowia lipolytica. Process Biochem. 2004, 39, 1469–1474. [Google Scholar] [CrossRef]
- Yin, X.; Madzak, C.; Du, G.; Zhou, J.; Chen, J. Enhanced alpha-ketoglutaric acid production in Yarrowia lipolytica WSH-Z06 by regulation of the pyruvate carboxylation pathway. Appl. Microbiol. Biotechnol. 2012, 96, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Yovkova, V.; Aurich, A.; Mauersberger, S.; Barth, G. Variation of the by-product spectrum during α-ketoglutaric acid production from raw glycerol by overexpression of fumarase and pyruvate carboxylase genes in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012, 95, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Rymowicz, W.; Fatykhova, A.R.; Kamzolova, S.V.; Rywinska, A.; Morgunov, I.G. Citric acid production from glycerol-containing waste of biodiesel industry by Yarrowia lipolytica in batch, repeated batch, and cell recycle regimes. Appl. Microbiol. Biotechnol. 2010, 87, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Abghari, A.; Madzak, C.; Chen, S. Combinatorial Engineering of Yarrowia lipolytica as a Promising Cell Biorefinery Platform for the de novo Production of Multi-Purpose Long Chain Dicarboxylic Acids. Fermentation 2017, 3, 40. [Google Scholar] [CrossRef]
- Sarchami, T.; Munch, G.; Johnson, E.; Kießlich, S.; Rehmann, L. A Review of Process-Design Challenges for Industrial Fermentation of Butanol from Crude Glycerol by Non-Biphasic Clostridium pasteurianum. Fermentation 2016, 2, 13. [Google Scholar] [CrossRef]
- Dedyukhina, E.G.; Chistyakova, T.I.; Vainshtein, M.B. Biosynthesis of arachidonic acid by Micromycetes. Appl. Biochem. Microbiol. 2011, 47, 109–117. [Google Scholar] [CrossRef]
- Dedyukhina, E.G.; Chistyakova, T.I.; Mironov, A.A.; Kamzolova, S.V.; Morgunov, I.G.; Vainshtein, M.B. Arachidonic acid synthesis from biodiesel-derived waste by Mortierella alpina. Eur. J. Lipid Sci. Technol. 2014, 116, 429–437. [Google Scholar] [CrossRef]
- Bellou, S.; Moustogianni, A.; Makri, A.; Aggelis, G. Lipids Containing Polyunsaturated Fatty Acids Synthesized by Zygomycetes Grown on Glycerol. Appl. Biochem. Biotechnol. 2012, 166, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Chatzifragkou, A.; Makri, A.; Belka, A.; Bellou, S.; Mavrou, M.; Mastoridou, M.; Mystrioti, P.; Onjaro, G.; Aggelis, G.; Papanikolaou, S. Biotechnological conversion of biodiesel derived waste glycerol by yeast and fungal species. Energy 2011, 36, 1097–1108. [Google Scholar] [CrossRef]
- Fakas, S.; Papanikolaou, S.; Batsos, A.; Galiotou-Panayotou, M.; Mallouchos, A.; Aggelis, G. Evaluating renewable carbon sources as substrates for single cell oil production by Cunninghamella echinulata and Mortierella isabellina. Biomass Bioeng. 2009, 33, 573–580. [Google Scholar] [CrossRef]
- Dedyukhina, E.G.; Chistyakova, T.I.; Mironov, A.A.; Kamzolova, S.V.; Minkevich, I.G.; Vainshtein, M.B. The effect of pH, aeration, and temperature on arachidonic acid synthesis by Mortierella alpina. Appl. Biochem. Microbiol. 2015, 5, 242–248. [Google Scholar] [CrossRef]
- Eroshin, V.K.; Dedyukhina, E.G.; Chistyakova, T.I.; Zhelifonova, V.P.; Kurtzman, C.P.; Bothast, J. Arachidonic-acid production by species of Mortierella. World J. Microbiol. Biotechnol. 1996, 12, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Eroshin, V.K.; Dedyukhina, E.G.; Chistyakova, T.I.; Zhelifonova, V.P.; Botast, R.J. Studies on arachidonic acid production by Mortierella fungi: A microbiological method for selecting arachidonic acid producers. Microbiology 1996, 65, 26–31. [Google Scholar]
- Lewis, T.; Nichols, P.D.; McMeekin, T.A. Evaluation of extraction methods for recovery of fatty acids from lipid-producing microheterotrophs. J. Microbiol. Methods 2000, 43, 107–116. [Google Scholar] [CrossRef]
- Minkevich, I.G.; Dedyukhina, E.G.; Chistyakova, T.I. The effect of lipid content on the elemental composition and energy capacity of yeast biomass. Appl. Microbiol. Biotechnol. 2010, 88, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.T. Production of arachidonic acid and dihomo-γ-linolenic acid from glycerol by oil-producing filamentous fungi, Morterella in the ARS culture collection. J. Ind. Microbiol. Biotechnol. 2008, 35, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Leman, J.; Brakoniecka-Sikorska, A. Effect of growth conditions on biosynthesis of oil by Mortierella ramanniana. Pol. J. Food Nutr. Sci. 1996, 46, 111–120. [Google Scholar]
- Peng, C.; Huang, He.; Ji, X.; Liu, X.; You, J.; Lu, J.; Cong, L.; Xu, X.; Ouyang, P. A temperature-shift strategy for efficient arachidonic acid fermentation by Mortierella alpina in batch culture. Biochem. Eng. J. 2010, 53, 92–96. [Google Scholar] [CrossRef]
- Kendrick, A.; Ratledge, C. Lipid formation in the oleaginous mold Entomophthora exitalis grown in continuous culture: Effects of growth rate, temperature and dissolved oxygen tension on polyunsaturated fatty acids. Appl. Microbiol. Biotechnol. 1992, 37, 18–22. [Google Scholar] [CrossRef]
- Chen, H.; Chang, C. Production of γ-linolenic acid by the fungus Cunninghamella echinulata CCRC 31840. Biotechnol. Prog. 1996, 12, 338–341. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Sarantou, S.; Komaitis, M.; Aggelis, G. Repression of reserve lipid turnover in Cunninghamella echinulata and Mortierella isabellina cultivated in multiple-limited media. J. Appl. Microbiol. 2004, 97, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.Y.; Chen, F. Lipid characterization of Mortierella alpina grown at different NaCl concentrations. J. Agric. Food Chem. 2008, 56, 7903–7909. [Google Scholar] [CrossRef] [PubMed]
- Shirashi, A.; Kawashima, H. Microbial Fermentation-Based Production of Phospholipids Containing Long-Chain Polyunsaturated Fatty Acids as Their Constituents. Patent WO2007091731 (A1), 16 August 2007. [Google Scholar]
- Evans, C.T.; Ratledge, C. A comparison of the oleaginous yeast, Candida curvata, grown on different carbon sources in continuous and batch culture. Lipids 1983, 18, 623–629. [Google Scholar] [CrossRef] [PubMed]

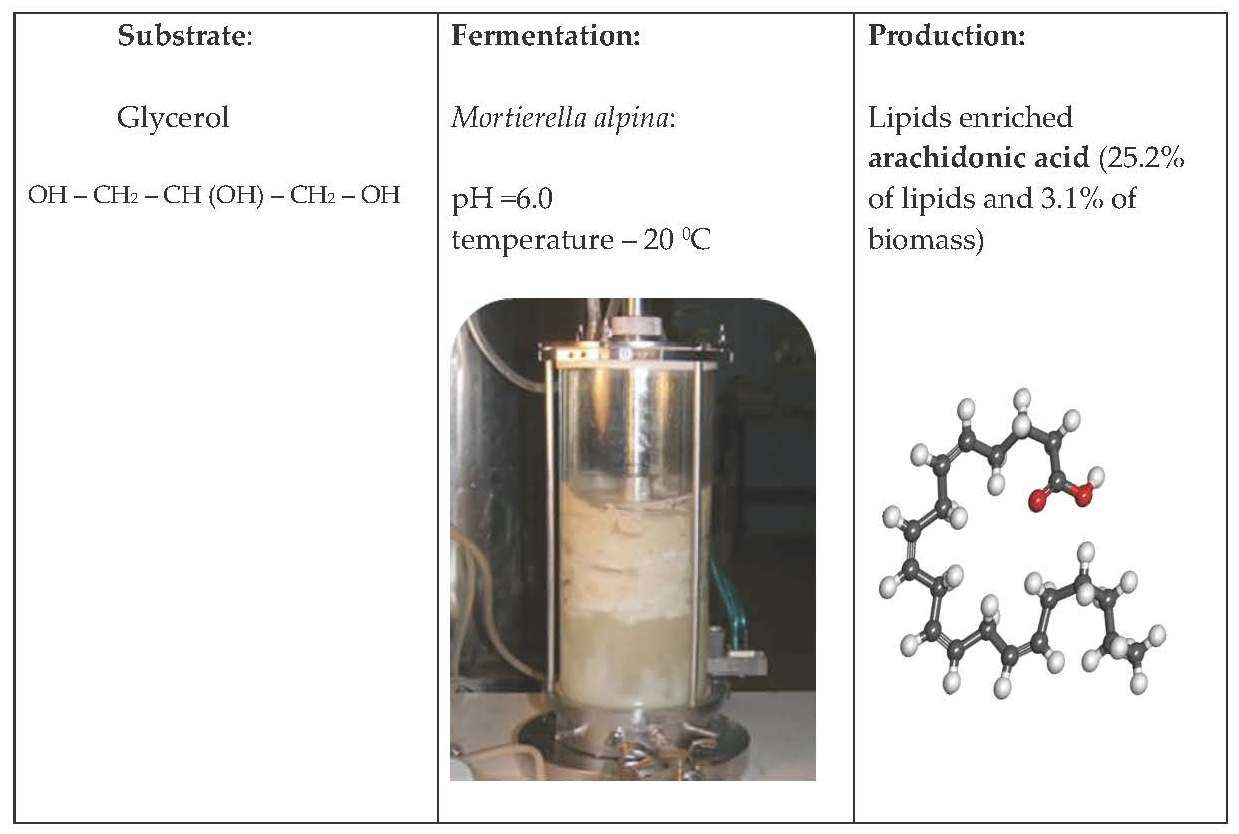{kind=link}
| Parameters | pH | |||||
|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | |
| Biomass (g L−1) | 6.8 | 7.4 | 14.4 | 10.8 | 10.4 | 8.2 |
| Lipids (% of biomass) | 11.3 | 10.3 | 12.8 | 16.1 | 13.5 | 12.0 |
| Lipids (g L−1) | 0.77 | 0.76 | 1.76 | 1.74 | 1.40 | 0.98 |
| Mycelium yield by mass (YX/S) (%) | 33.3 | 33.0 | 37.8 | 38.8 | 37.9 | 30.3 |
| Energy capacity of biomass (QB) (kJ g−1) | 18.3 | 18.0 | 18.7 | 19.6 | 18.9 | 18.5 |
| Mycelium yield by energy (ηX/S) (%) | 33.9 | 33.0 | 39.4 | 42.3 | 39.8 | 31.2 |
| Lipid yield by mass (YL/S) (%) | 3.7 | 3.4 | 4.6 | 6.3 | 5.1 | 3.6 |
| Lipid yield by energy (ηL/S) (%) | 8.8 | 8.1 | 10.9 | 14.9 | 12.1 | 8.5 |
| AA (% of lipids) | 11.3 | 14.7 | 17.8 | 20.1 | 21.2 | 22.1 |
| AA (g L−1) | 1.27 | 1.52 | 2.27 | 3.24 | 2.87 | 2.64 |
| AA yield by mass (YAA/S) (%) | 0.086 | 0.112 | 0.320 | 0.230 | 0.290 | 0.217 |
| AA yield by energy (ηAA/S) (%) | 0.4 | 0.5 | 0.8 | 0.8 | 1.0 | 0.8 |
| Parameters | pH | |||||
|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | |
| Biomass (g L−1) | 6.8 | 8.1 | 21.0 | 22.0 | 17.8 | 17.8 |
| Lipids (% of biomass) | 17.4 | 12.7 | 32.3 | 30.9 | 22.3 | 34.7 |
| Lipids (g L−1) | 18.1 | 18.0 | 18.3 | 19.0 | 19.4 | 18.7 |
| Mycelium yield by mass (YX/S) (%) | 17.5 | 12.7 | 32.9 | 32.6 | 24.6 | 36.1 |
| Energy capacity of biomass (QB) (kJ g−1) | 10.8 | 10.3 | 11.4 | 14.1 | 15.2 | 13.1 |
| Mycelium yield by energy (ηX/S) (%) | 0.73 | 0.83 | 2.39 | 3.10 | 2.70 | 2.33 |
| Lipid yield by mass (YL/S) (%) | 1.8 | 1.3 | 3.7 | 4.4 | 3.4 | 4.6 |
| Lipid yield by energy (ηL/S) (%) | 4.3 | 3.0 | 8.8 | 10.4 | 8.0 | 10.9 |
| AA (% of lipids) | 0 | 0.5 | 14.6 | 19.9 | 19.0 | 19.8 |
| AA (g L−1) | 0 | 0.13 | 1.72 | 2.83 | 2.67 | 2.65 |
| AA yield by mass (YAA/S) (%) | 0 | 0.008 | 0.367 | 0.622 | 0.516 | 0.463 |
| AA yield by energy (ηAA/S) (%) | 0 | 0.02 | 0.56 | 0.87 | 0.64 | 0.90 |
| Fatty Acids | pH | |||||
|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | |
| C14 | 1.9 | 1.6 | 1.2 | 1.2 | 1.2 | 1.4 |
| C15 | 0.1 | 0.1 | 0 | 0.1 | 0.1 | 0.1 |
| C16 | 12.2 | 10.1 | 8.4 | 6.8 | 6.8 | 6.6 |
| C18 | 18.4 | 17.6 | 12.9 | 12.5 | 14.0 | 12.3 |
| C18:1 | 41.1 | 42.0 | 45.5 | 44.7 | 40.7 | 41.5 |
| C18:2 | 9.1 | 6.9 | 10.7 | 7.8 | 8.9 | 8.9 |
| γ-C18:3 | 4.9 | 5.6 | 0.3 | 5.6 | 5.4 | 6.1 |
| C20 | 0.4 | 0.4 | 3.1 | 0.3 | 0.2 | 0.3 |
| C20:1 | 0 | 0.3 | 0 | 0.4 | 0.3 | 0 |
| C20:2 | 0 | 0 | 0 | 0 | 0 | 0 |
| C20:3 | 0.6 | 0.7 | 0.0 | 0.6 | 1.2 | 0.7 |
| C20:4 | 11.3 | 14.7 | 17.8 | 20.1 | 21.2 | 22.1 |
| C20:4/C18:2 | 1.24 | 2.13 | 1.66 | 2.57 | 2.38 | 2.48 |
| C18:2/C18:1 | 0.22 | 0.16 | 0.23 | 0.17 | 0.21 | 0.21 |
| C18:1/C18 | 2.23 | 2.38 | 3.52 | 3.57 | 2.90 | 3.37 |
| Fatty Acids | pH | |||||
|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | |
| C14 | 4.2 | 3.5 | 2.3 | 1.9 | 1.4 | 1.8 |
| C15 | 0.9 | 1.1 | 0.2 | 0.1 | 0.1 | 0.2 |
| C16 | 27.9 | 28.6 | 18.5 | 16.7 | 24.9 | 17.8 |
| C18 | 17.4 | 19.6 | 13.0 | 9.4 | 8.6 | 9.8 |
| C18:1 | 40.0 | 38.7 | 32.2 | 31.5 | 27.2 | 31.7 |
| C18:2 | 6.3 | 3.7 | 12.6 | 15.1 | 13.9 | 14.3 |
| γ-C18:3 | 1.6 | trace | 3.9 | 4.0 | 3.0 | 3.8 |
| C20 | 0.6 | 1.4 | 0.8 | trace | trace | trace |
| C20:1 | trace | trace | trace | 1.3 | 0.8 | 0.6 |
| C20:2 | trace | trace | 0.5 | trace | trace | trace |
| C20:3 | 1.03 | 2.4 | 1.3 | 0 | 1.2 | 0.3 |
| C20:4 | 0 | 0.5 | 14.6 | 19.9 | 19.0 | 19.8 |
| C20:4/C18:2 | 0 | 0.14 | 1.16 | 1.32 | 1.37 | 1.38 |
| C18:2/C18:1 | 0.16 | 0.09 | 0.39 | 0.48 | 0.51 | 0.45 |
| C18:1/C18 | 2.30 | 1.97 | 2.48 | 3.35 | 3.16 | 3.23 |
| Parameters | Temperature (°C) | ||||
|---|---|---|---|---|---|
| 20 | 22 | 24 | 26 | 28 | |
| Biomass (g L−1) | 16.4 | 17.2 | 16.2 | 17.2 | 17.7 |
| Consumed glycerol (g L−1) | 30.0 | 31.2 | 32.5 | 33.2 | 34.0 |
| Lipids (% of biomass) | 12.2 | 11.5 | 16.3 | 19.3 | 22.2 |
| Lipids (g L−1) | 2.0 | 1.9 | 2.6 | 3.4 | 3.9 |
| Mycelium yield by mass (YX/S) (%) | 54.7 | 55.1 | 49.8 | 51.8 | 52.0 |
| Energy capacity of biomass (QB) (kJ g−1) | 18.5 | 18.3 | 19.6 | 20.5 | 21.3 |
| Mycelium yield by energy (ηX/S) (%) | 56.3 | 56.1 | 54.3 | 59.1 | 61.7 |
| Lipid yield by mass (YL/S) (%) | 6.6 | 6.1 | 8.0 | 9.9 | 11.4 |
| Lipid yield by energy (ηL/S) (%) | 15.6 | 14.5 | 19.0 | 23.4 | 27.0 |
| AA (% of lipids) | 25.2 | 20.8 | 19.6 | 18.9 | 18.2 |
| AA (g L−1) | 0.5 | 0.4 | 0.5 | 0.6 | 0.7 |
| AA yield by mass (YAA/S) (%) | 1.6 | 1.3 | 1.5 | 1.8 | 2.1 |
| AA yield by energy (ηAA/S) (%) | 3.4 | 2.8 | 3.2 | 3.8 | 4.5 |
| Fatty Acids | Temperature (°C) | ||||
|---|---|---|---|---|---|
| 20 | 22 | 24 | 26 | 28 | |
| C14 | 1.7 | 1.8 | 1.8 | 1.7 | 1.4 |
| C15 | 0.2 | 0.2 | 0.3 | 0.2 | 0.5 |
| C16 | 18.9 | 21.3 | 22.6 | 21.5 | 23.5 |
| C18 | 11.0 | 12.9 | 12.4 | 15.5 | 8.8 |
| C18:1 | 23.9 | 22.1 | 27.3 | 23.9 | 29.9 |
| C18:2 | 15.0 | 16.7 | 12.6 | 12.9 | 8.8 |
| γ-C18:3 | 3.0 | 4.2 | 2.9 | 4.9 | 4.6 |
| C20 | 0.9 | 0 | 0.6 | 0.3 | 0.7 |
| C20:1 | 0 | 0 | 0 | 0.2 | 0.7 |
| C20:2 | 0.4 | 0 | 0 | 0 | 3.1 |
| C20:3 | 25.2 | 20.8 | 19.6 | 18.9 | 18.2 |
| C20:4 | 1.7 | 1.8 | 1.8 | 1.7 | 1.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mironov, A.A.; Nemashkalov, V.A.; Stepanova, N.N.; Kamzolova, S.V.; Rymowicz, W.; Morgunov, I.G. The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995. Fermentation 2018, 4, 17. https://doi.org/10.3390/fermentation4010017
Mironov AA, Nemashkalov VA, Stepanova NN, Kamzolova SV, Rymowicz W, Morgunov IG. The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995. Fermentation. 2018; 4(1):17. https://doi.org/10.3390/fermentation4010017
Chicago/Turabian StyleMironov, Aleksei A., Vitaly A. Nemashkalov, Nadezda N. Stepanova, Svetlana V. Kamzolova, Waldemar Rymowicz, and Igor G. Morgunov. 2018. "The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995" Fermentation 4, no. 1: 17. https://doi.org/10.3390/fermentation4010017
APA StyleMironov, A. A., Nemashkalov, V. A., Stepanova, N. N., Kamzolova, S. V., Rymowicz, W., & Morgunov, I. G. (2018). The Effect of pH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella alpina NRRL-A-10995. Fermentation, 4(1), 17. https://doi.org/10.3390/fermentation4010017