
Wine Aroma Characterization of the Two Main Fermentation Yeast Species of the Apiculate Genus Hanseniaspora

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeasts and Fermentation in Natural Grape Must
2.2. Aroma Characterization
2.3. Genomic Analysis
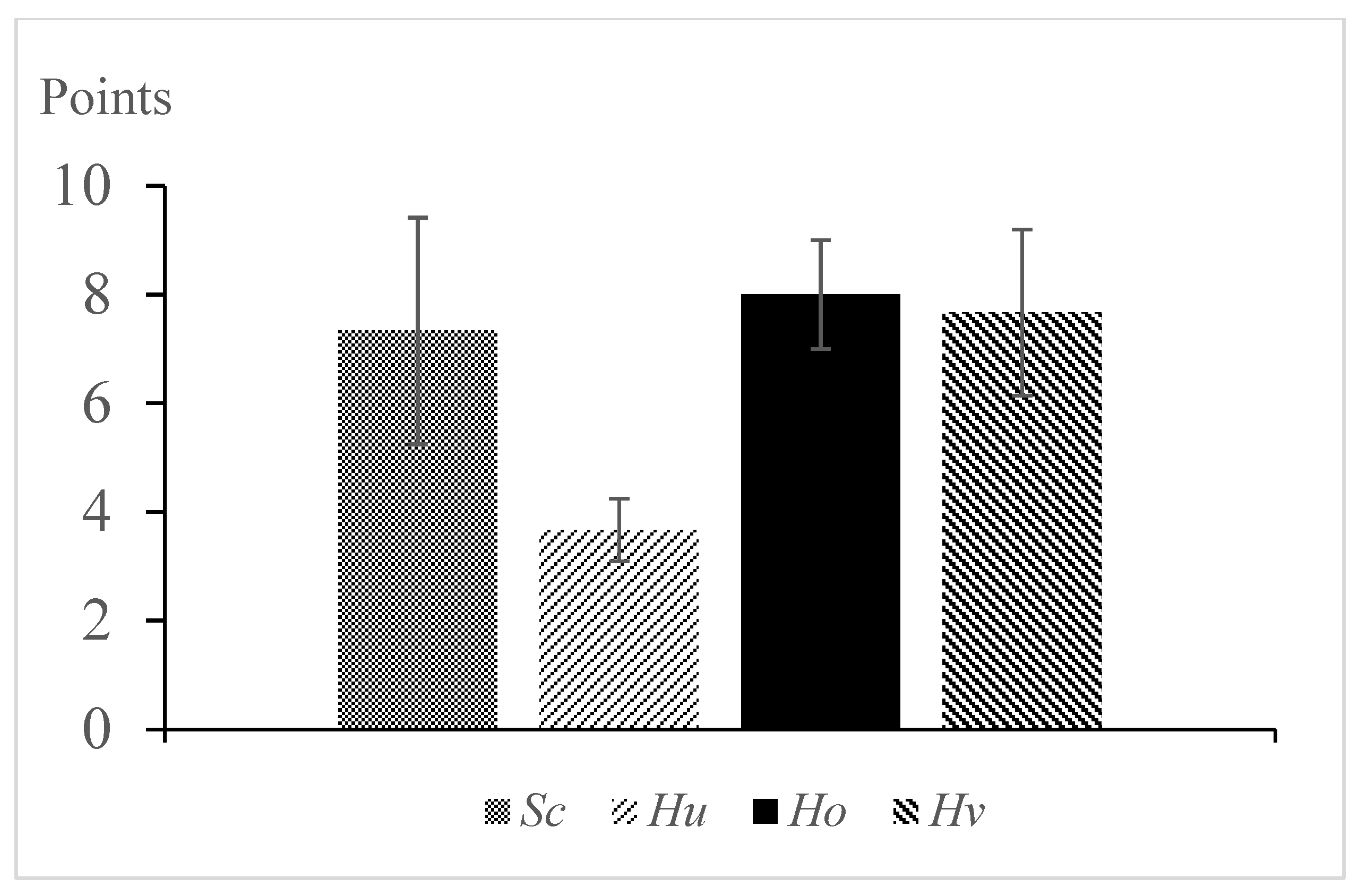
2.4. Sensory Analysis
2.5. Statistical Analysis
3. Results
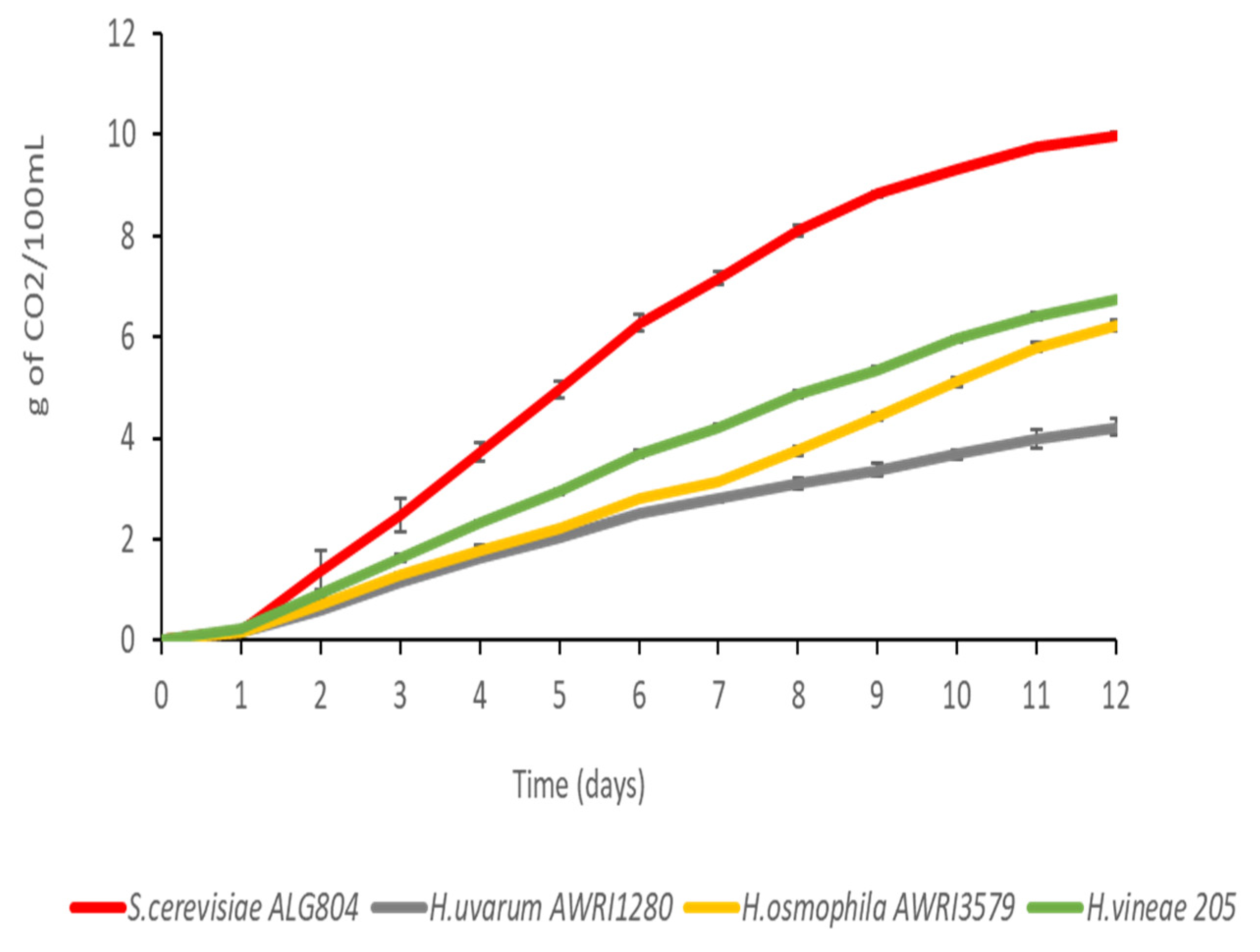
3.1. Fermentation and Aroma Profile of Wines Elaborated with Hanseniaspora Spp.
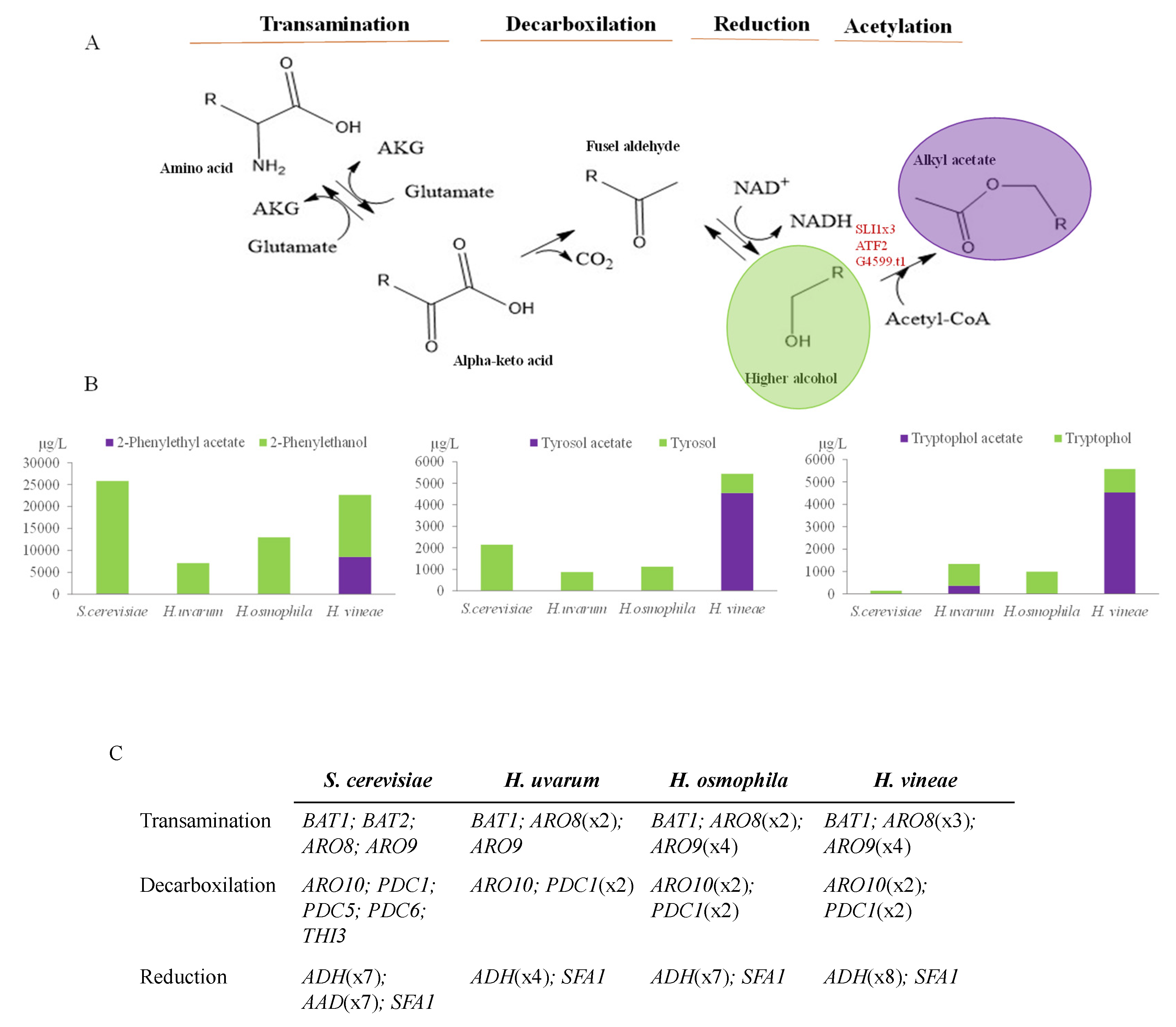
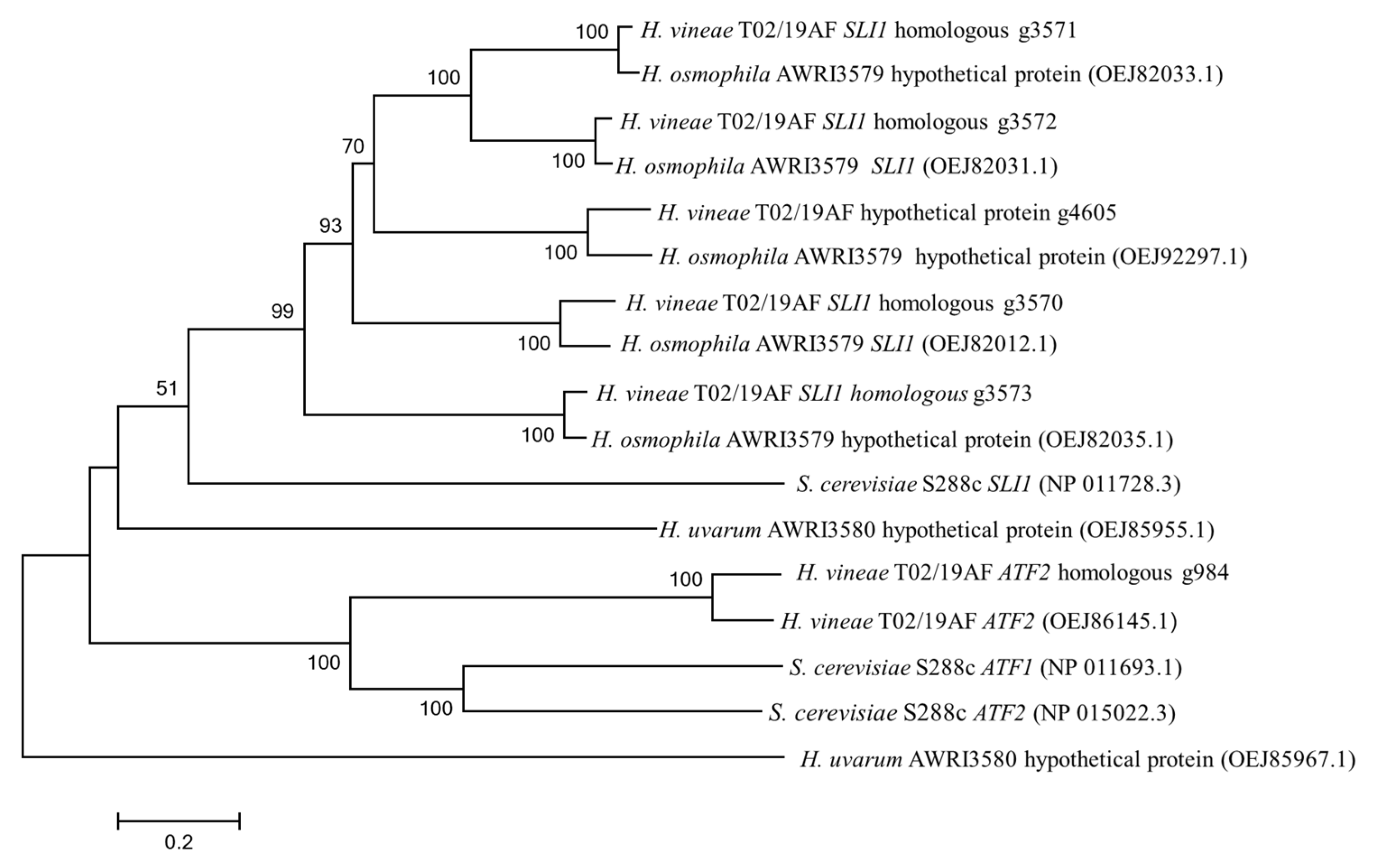
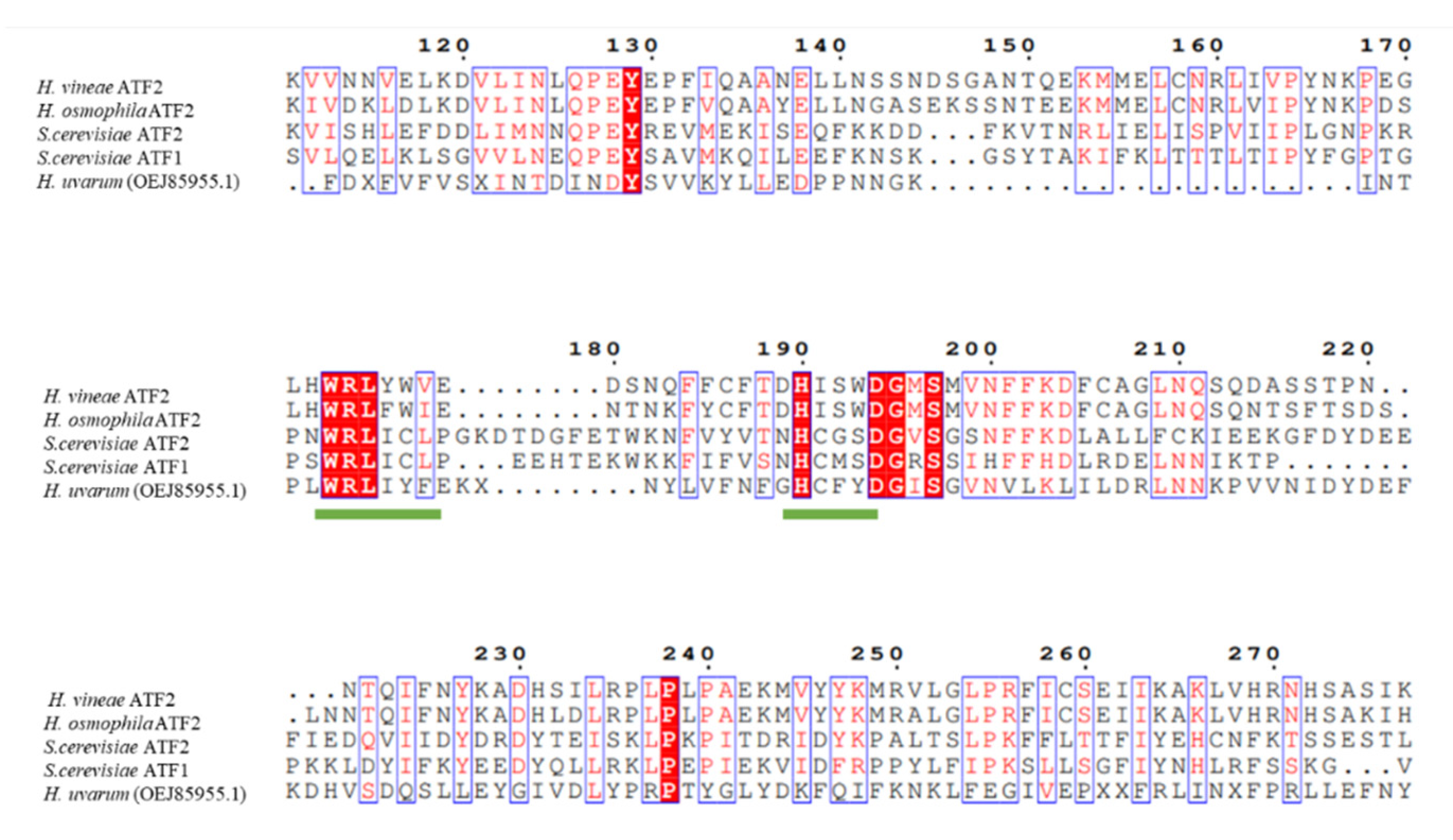
3.2. Genetic Analysis of Putative Acetyltransferases in Hanseniaspora Spp.
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Carrau, F.; Henschke, P.A. Hanseniaspora vineae and the concept of friendly yeasts to increase autochthonous wine flavor diversity. Front. Microbiol. 2021, 12, 702093. [Google Scholar] [CrossRef]
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and ecological diversity of food and beverage yeasts. In Yeasts in Foods and Beverages; Querol, A., Fleet, G., Eds.; Springer: Heidelberg, Germany, 2006; pp. 13–53. [Google Scholar]
- Albertin, W.; Setati, M.E.; Miot-Sertier, C.; Mostert, T.T.; Colonna-Ceccaldi, B.; Coulon, J.; Girard, P.; Moine, V.; Pillet, M.; Salin, F.; et al. Hanseniaspora uvarum from winemaking environments show spatial and temporal genetic clustering. Front. Microbiol. 2016, 6, 1569. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Growth of non-Saccharomyces yeasts affects nutrient availability for Saccharomyces cerevisiae during wine fermentation. Int. J. Food Microbiol. 2012, 157, 245–250. [Google Scholar] [CrossRef]
- Pina, C.; Santos, C.; Couto, J.A.; Hogg, T. Ethanol tolerance of five non-Saccharomyces wine yeasts in comparison with a strain of Saccharomyces cerevisiae—Influence of different culture conditions. Food Microbiol. 2004, 21, 439–447. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Lleixà, J.; Manzano, M.; Mas, A.; Portillo, M.d.C. Saccharomyces and non-Saccharomyces competition during microvinification under different sugar and nitrogen conditions. Front. Microbiol. 2016, 7, 1959. [Google Scholar] [CrossRef]
- González, B.; Vázquez, J.; Cullen, P.J.; Mas, A.; Beltran, G.; Torija, M.J. Aromatic amino acid-derived compounds induce morphological changes and modulate the cell growth of wine yeast species. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valera, M.J.; Morcillo-Parra, M.Á.; Zagórska, I.; Mas, A.; Beltran, G.; Torija, M.J. Effects of melatonin and tryptophol addition on fermentations carried out by Saccharomyces cerevisiae and non-Saccharomyces yeast species under different nitrogen conditions. Int. J. Food Microbiol. 2019, 289, 174–181. [Google Scholar] [CrossRef]
- Martin, V.; Valera, M.J.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Borren, E.; Tian, B. The important contribution of non-Saccharomyces yeasts to the aroma complexity of wine: A review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Čadež, N.; Bellora, N.; Ulloa, R.; Hittinger, C.T.; Libkind, D. Genomic content of a novel yeast species Hanseniaspora gamundiae sp. Nov. From fungal stromata (Cyttaria) associated with a unique fermented beverage in Andean Patagonia, Argentina. PLoS ONE 2019, 14, e0210792. [Google Scholar] [CrossRef] [PubMed]
- Valera, M.J.; Boido, E.; Dellacassa, E.; Carrau, F. Comparison of the glycolytic and alcoholic fermentation pathways of Hanseniaspora vineae with Saccharomyces cerevisiae wine yeasts. Fermentation 2020, 6, 78. [Google Scholar] [CrossRef]
- Steenwyk, J.L.; Opulente, D.A.; Kominek, J.; Shen, X.X.; Zhou, X.; Labella, A.L.; Bradley, N.P.; Eichman, B.F.; Čadež, N.; Libkind, D.; et al. Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts. PLoS Biol. 2019, 17, 1–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, K.; Jin, G.J.; Xu, Y.H.; Tao, Y.S. Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Medina, K. Biodiversidad de Levaduras No-Saccharomyces: Efecto del Metabolismo Secundario en el Color y el Aroma de Vinos de Calidad. Ph.D. Thesis, Universidad de la República, Montevideo, Uruguay, 2014. [Google Scholar]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Escott, C.; Loira, I.; Herbert-Pucheta, J.E.; Schneider, R.; Carrau, F.; Cuerda, R.; Morata, A. Impact of Hanseniaspora vineae in alcoholic fermentation and ageing on lees of high-quality white wine. Fermentation 2020, 6, 66. [Google Scholar] [CrossRef]
- Giorello, F.; Valera, M.J.; Martin, V.; Parada, A.; Salzman, V.; Camesasca, L.; Fariña, L.; Boido, E.; Medina, K.; Dellacassa, E.; et al. Genomic and transcriptomic basis of Hanseniaspora vineae’s impact on flavor diversity and wine quality. Appl. Environ. Microbiol. 2019, 85, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef]
- Viana, F.; Belloch, C.; Vallés, S.; Manzanares, P. Monitoring a mixed starter of Hanseniaspora vineae-Saccharomyces cerevisiae in natural must: Impact on 2-phenylethyl acetate production. Int. J. Food Microbiol. 2011, 151, 235–240. [Google Scholar] [CrossRef]
- Giorello, F.M.; Berna, L.; Greif, G.; Camesasca, L.; Salzman, V.; Medina, K.; Robello, C.; Gaggero, C.; Aguilar, P.S.; Carrau, F. Genome Sequence of the native apiculate wine yeast Hanseniaspora vineae T02/19AF. Genome Announc. 2014, 2, e00530-14. [Google Scholar] [CrossRef] [Green Version]
- Fariña, L.; Medina, K.; Urruty, M.; Boido, E.; Dellacassa, E.; Carrau, F. Redox effect on volatile compound formation in wine during fermentation by Saccharomyces cerevisiae. Food Chem. 2012, 134, 933–939. [Google Scholar] [CrossRef]
- Boido, E.; Lloret, A.; Medina, K.; Farñia, L.; Carrau, F.; Versini, G.; Dellacassa, E. Aroma composition of Vitis vinifera cv. Tannat: The typical red wine from Uruguay. J. Agric. Food Chem. 2003, 51, 5408–5413. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Boido, E.; Lorenzo, D.; Medina, K.; Carrau, F.; Dellacassa, E.; Versini, G. Aroma variation in Tannat wines: Effect of malolactic fermentation on ethyl lactate level and its enantiomeric distribution. Ital. J. Food Sci. 2002, 14, 175–180. [Google Scholar]
- Martin, V.; Giorello, F.; Fariña, L.; Minteguiaga, M.; Salzman, V.; Boido, E.; Aguilar, P.S.; Gaggero, C.; Dellacassa, E.; Mas, A.; et al. De novo synthesis of benzenoid compounds by the yeast Hanseniaspora vineae increases the flavor diversity of wines. J. Agric. Food Chem. 2016, 64, 4574–4583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrau, F.M.; Medina, K.; Farina, L.; Boido, E.; Henschke, P.A.; Dellacassa, E. Production of fermentation aroma compounds by Saccharomyces cerevisiae wine yeasts: Effects of yeast assimilable nitrogen on two model strains. FEMS Yeast Res. 2008, 8, 1196–1207. [Google Scholar] [CrossRef] [Green Version]
- Lopez De Lerma, N.; Bellincontro, A.; Mencarelli, F.; Moreno, J.; Peinado, R.A. Use of electronic nose, validated by GC-MS, to establish the optimum off-vine dehydration time of wine grapes. Food Chem. 2012, 130, 447–452; [Google Scholar] [CrossRef]
- Fariña, L.; Villar, V.; Ares, G.; Carrau, F.; Dellacassa, E.; Boido, E. Volatile composition and aroma profile of Uruguayan Tannat wines. Food Res. Int. 2015, 69, 244–255. [Google Scholar] [CrossRef]
- Kuzuyama, T.; Dairi, T.; Yamashita, H.; Shoji, Y.; Seto, H. Heterologous Mevalonate Production in Streptomyces lividans TK23. Biosci. Biotechnol. Biochem. 2004, 68, 931–934. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, H. Commercial production of mevalonolactone by fermentation and the application to skin cosmetics with anti-aging effect. Fragr. J. 2000, 28, 62–65. [Google Scholar]
- Romano, P.; Suzzi, G. Origin and production of acetoin during wine yeast fermentation. Appl. Environ. Microbiol. 1996, 62, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Momoi, M.; Tanoue, D.; Sun, Y.; Takematsu, H.; Suzuki, Y.; Suzuki, M.; Suzuki, A.; Fujita, T.; Kozutsumi, Y. SLI1 (YGR212W) is a major gene conferring resistance to the sphingolipid biosynthesis inhibitor ISP-1, and encodes an ISP-1 N-acetyltransferase in yeast. Biochem. J. 2004, 381, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.-P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef] [Green Version]
- Saerens, S.M.G.; Verbelen, P.J.; Vanbeneden, N.; Thevelein, J.M.; Delvaux, F.R. Monitoring the influence of high-gravity brewing and fermentation temperature on flavour formation by analysis of gene expression levels in brewing yeast. Appl. Microbiol. Biotechnol. 2008, 80, 1039–1051. [Google Scholar] [CrossRef]
- Van Laere, S.D.M.; Saerens, S.M.G.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Flavour formation in fungi: Characterisation of KlAtf, the Kluyveromyces lactis orthologue of the Saccharomyces cerevisiae alcohol acetyltransferases Atf1 and Atf2. Appl. Microbiol. Biotechnol. 2008, 78, 783–792. [Google Scholar] [CrossRef]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant. Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; Alvarez-Huerta, M.; Neef, E.; Verstappen, F.W.A.; Bouwmeester, H.J.; Aharoni, A. Functional characterization of enzymes forming volatile esters from strawberry and banana. Plant. Physiol. 2004, 135, 1865–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, A.B.; Dufour, J.P. Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 2000, 16, 1287–1298. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | S. cerevisiae ALG804 | H. uvarum AWRI1280 | H. osmophila AWRI3579 | H. vineae 025 |
|---|---|---|---|---|
| Residual sugars (g/L) | 1.6 ± 0.6 | 115.6 ± 11.8 | 86.8 ± 6.2 | 69.1 ± 12.1 |
| Volatile acidity (g/L) | 0.45 ± 0.05 | 0.91 ± 0.12 | 0.44 ± 0.14 | 0.34 ± 0.02 |
| S. cerevisiae | H. uvarum | H. osmophila | H. vineae | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LRI | Alcohols | ||||||||||||
| 1221 | 3-Methylbutanol | 113496 a* | ± | 35366 | 24130 | b ± | 8342 | 52005 | b ± | 10409 | 63075 | ab ± | 21210 |
| 1264 | Acetoin | 1764 a | ± | 2628 | 127315 b | ± | 4500 | 2351 a | ± | 741 | 20419 b | ± | 6535 |
| 1341 | 1-Hexanol | 487 | ± | 42 | 294 | ± | 227 | 266 | ± | 134 | 474 | ± | 168 |
| 1389 | 3-Ethoxy-1-propanol | 9868 b | ± | 1767 | 689 a | ± | 16 | 972 a | ± | 246 | 412 a | ± | 220 |
| 1453 | 2-Ethyl-1-hexanol | 28 a | ± | 12 | 75 a,b | ± | 8 | 48 a | ± | 10 | 137 b,c | ± | 37 |
| 1526 | 2,3-Butanediol | 1594 b | ± | 351 | 194 a | ± | 206 | 515 a | ± | 77 | 268 a | ± | 384 |
| 1822 | Benzyl alcohol | 7 a | ± | 1 | 9 a | ± | 2 | 9 a | ± | 1 | 25 b | ± | 5 |
| 1906 | 2-Phenylethanol | 25710 b | ± | 5352 | 7035 a | ± | 1081 | 12959 a | ± | 2781 | 14130ca | ± | 898 |
| 3052 | Tyrosol | 2149 b | ± | 802 | 879 a | ± | 114 | 1104 a,b | ± | 193 | 892 a | ± | 271 |
| 3514 | Tryptophol | 141 a | ± | 65 | 1107 c | ± | 13 | 982 b,c | ± | 304 | 1053 b,c | ± | 152 |
| Esters | |||||||||||||
| 1227 | Ethyl hexanoate | 347 b | ± | 110 | nd a | 59 a | ± | 15 | 9 a | ± | 9 | ||
| 1341 | Ethyl lactate | 55 a,b | ± | 7 | 62 a,b | ± | 34 | 201 c | ± | 57 | 45 a | ± | 19 |
| 1439 | Ethyl octanoate | 1027 b | ± | 292 | 42 a | ± | 31 | 164 a | ± | 27 | 38 a | ± | 7 |
| 1516 | Ethyl 3-hydroxybutanoate | 50 b | ± | 17 | nda | nda | 15 a | ± | 6 | ||||
| 1626 | Ethyl decanoate | 729 d | ± | 126 | 125 a,b | ± | 32 | 312 c | ± | 55 | 271 b,c | ± | 52 |
| 1650 | 1,3-Propanediol diacetate | 3401 b | ± | 875 | 567 a | ± | 318 | 1257 a | ± | 462 | 1197 a | ± | 313 |
| 1813 | 2-Phenylethyl acetate | 116 a | ± | 53 | 46 a | ± | 41 | 27 a | ± | 3 | 10524 b | ± | 3209 |
| 2130 | Ethyl pentadecanoate | nd a | 47 a,b | ± | 34 | 68 b | ± | 22 | 9 a | ± | 10 | ||
| 2995 | Tyrosol acetate | nd a | nda | 14 a | ± | 6 | 4547 b | ± | 3196 | ||||
| 3405 | Tryptophol acetate | nd a | 108 b | ± | 3 | nd a | 6787 c | ± | 2007 | ||||
| Acids | |||||||||||||
| 1510 | Propanoic acid | 46 | ± | 25 | 158 | ± | 117 | 192 | ± | 50 | 59 | ± | 6 |
| 1588 | Isobutanoic acid | 138 a | ± | 11 | 394 b | ± | 136 | 410 b | ± | 93 | 834 c | ± | 56 |
| 1625 | Butanoic acid | 170 b | ± | 29 | 47 a | ± | 10 | 67 a | ± | 9 | 81 a | ± | 34 |
| 1650 | 3-Methylbutanoic acid | 306 | ± | 160 | 343 | ± | 293 | 541 | ± | 298 | 313 | ± | 293 |
| 1843 | Hexanoic acid | 1365 b | ± | 133 | 234 a | ± | 136 | 290 a | ± | 25 | 211 a | ± | 109 |
| 2070 | Octanoic acid | 2143 b | ± | 370 | 182 a | ± | 120 | 382 a | ± | 60 | 128 a | ± | 29 |
| 2243 | Decanoic acid | 472 c | ± | 51 | 96 a | ± | 29 | 167 a,b | ± | 81 | 473 b | ± | 276 |
| Phenols | |||||||||||||
| 2173 | 4-Vinylguaiacol | 55 | ± | 19 | 26 | ± | 10 | 61 | ± | 56 | 33 | ± | 8 |
| Others | |||||||||||||
| 1620 | γ-Butyrolactone | 125 a | ± | 28 | 270 b | ± | 76 | 206 a,b | ± | 13 | 147 a | ± | 12 |
| 1750 | Valerolactone | nd a | nd a | 25 b | ± | 8 | 37 c | ± | 4 | ||||
| 2097 | Pantolactone | nd a | nda | 17 b | ± | 1 | nd a | ||||||
| 2007 | 1,4-Dimethyl piperazine | nd a | 112 b | ± | 29 | nd a | nd a | ||||||
| 2594 | Mevalonolactone | 67 a,b | ± | 16 | 72 a,b | ± | 19 | 38 a | ± | 7 | 235 c | ± | 64 |
| S. cerevisiae | H. uvarum | H. osmophila | H. vineae | |
|---|---|---|---|---|
| ATF-like | ATF1; ATF2 | ATF2 | ATF2 | |
| SLI1-like | SLI1 | SLI1x2 | SLI1x4 | |
| Non-annotated | OEJ85955.1; OEJ85967.1 | OEJ82033.1; OEJ92297.1; OEJ82035.1 | g4605.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valera, M.J.; Olivera, V.; Boido, E.; Dellacassa, E.; Carrau, F. Wine Aroma Characterization of the Two Main Fermentation Yeast Species of the Apiculate Genus Hanseniaspora. Fermentation 2021, 7, 162. https://doi.org/10.3390/fermentation7030162
Valera MJ, Olivera V, Boido E, Dellacassa E, Carrau F. Wine Aroma Characterization of the Two Main Fermentation Yeast Species of the Apiculate Genus Hanseniaspora. Fermentation. 2021; 7(3):162. https://doi.org/10.3390/fermentation7030162
Chicago/Turabian StyleValera, María José, Valentina Olivera, Eduardo Boido, Eduardo Dellacassa, and Francisco Carrau. 2021. "Wine Aroma Characterization of the Two Main Fermentation Yeast Species of the Apiculate Genus Hanseniaspora" Fermentation 7, no. 3: 162. https://doi.org/10.3390/fermentation7030162
APA StyleValera, M. J., Olivera, V., Boido, E., Dellacassa, E., & Carrau, F. (2021). Wine Aroma Characterization of the Two Main Fermentation Yeast Species of the Apiculate Genus Hanseniaspora. Fermentation, 7(3), 162. https://doi.org/10.3390/fermentation7030162