
Metabolic Potential of Some Functional Groups of Bacteria in Aquatic Urban Systems

 , ,
, , 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Sampling Site
2.2. Sampling
2.3. Media and Cultivation
2.4. Data Analysis
3. Results
3.1. Environmental Parameters
3.2. Heterotrophic Bacteria
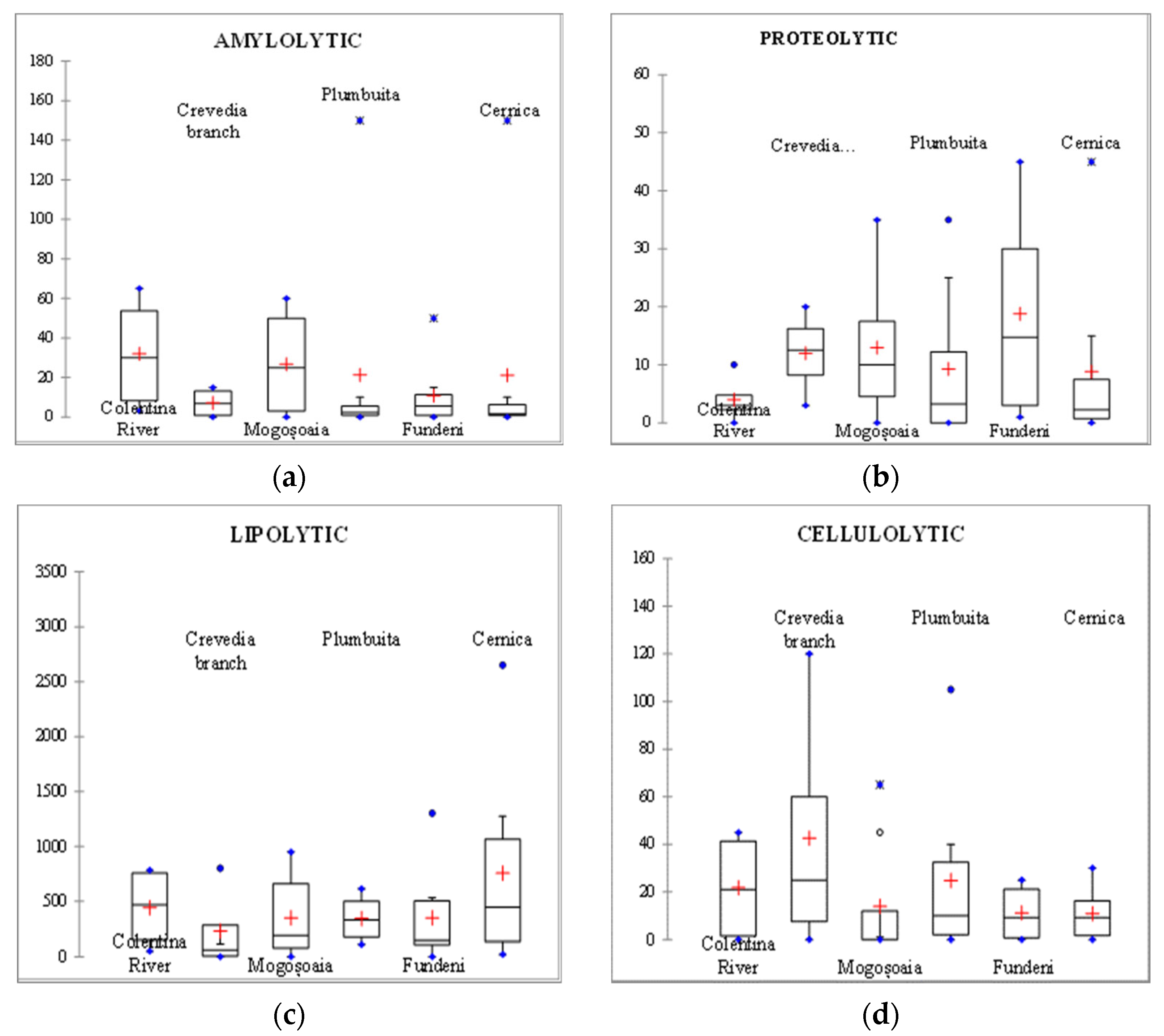
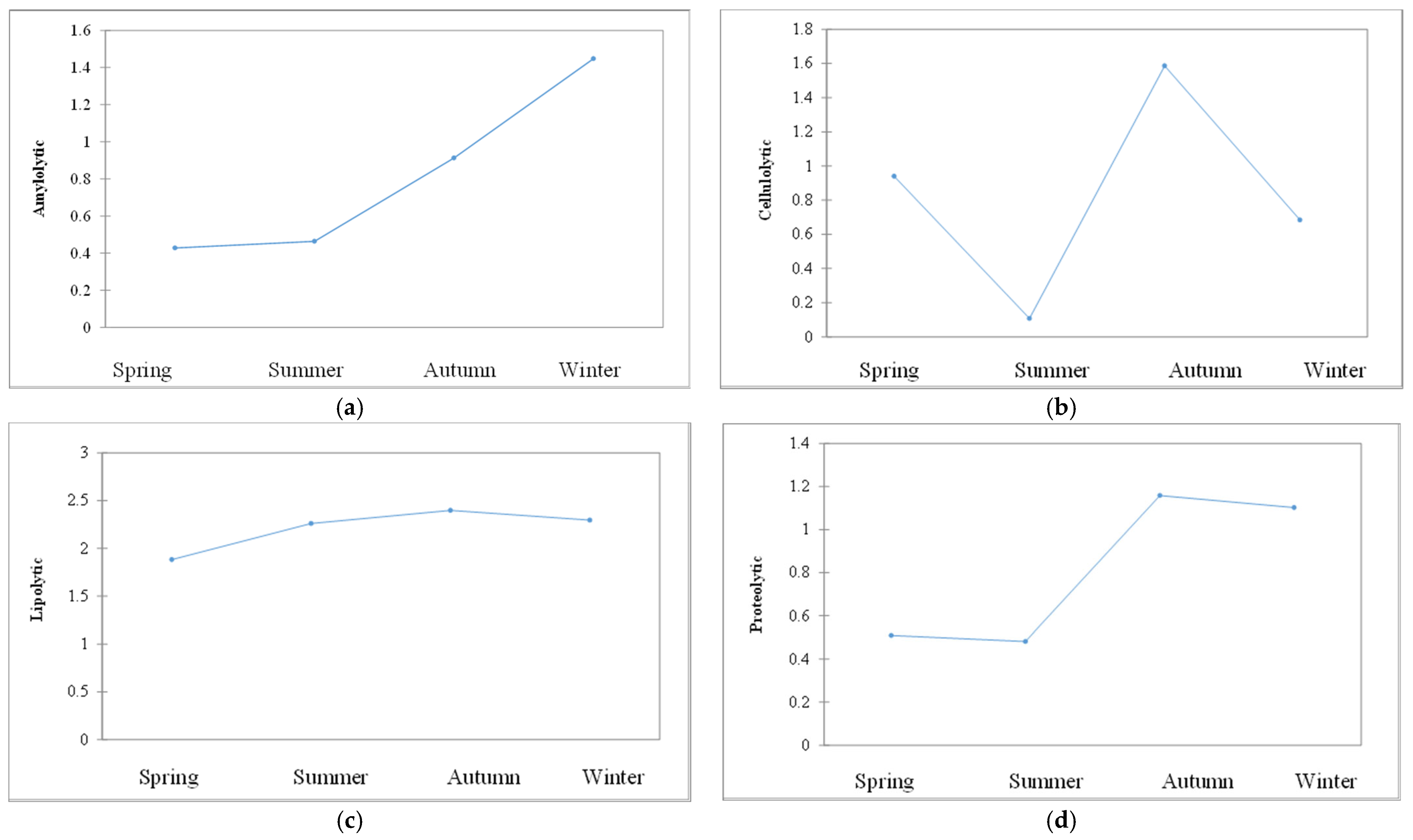
3.3. Hydrolytic Exoenzymes
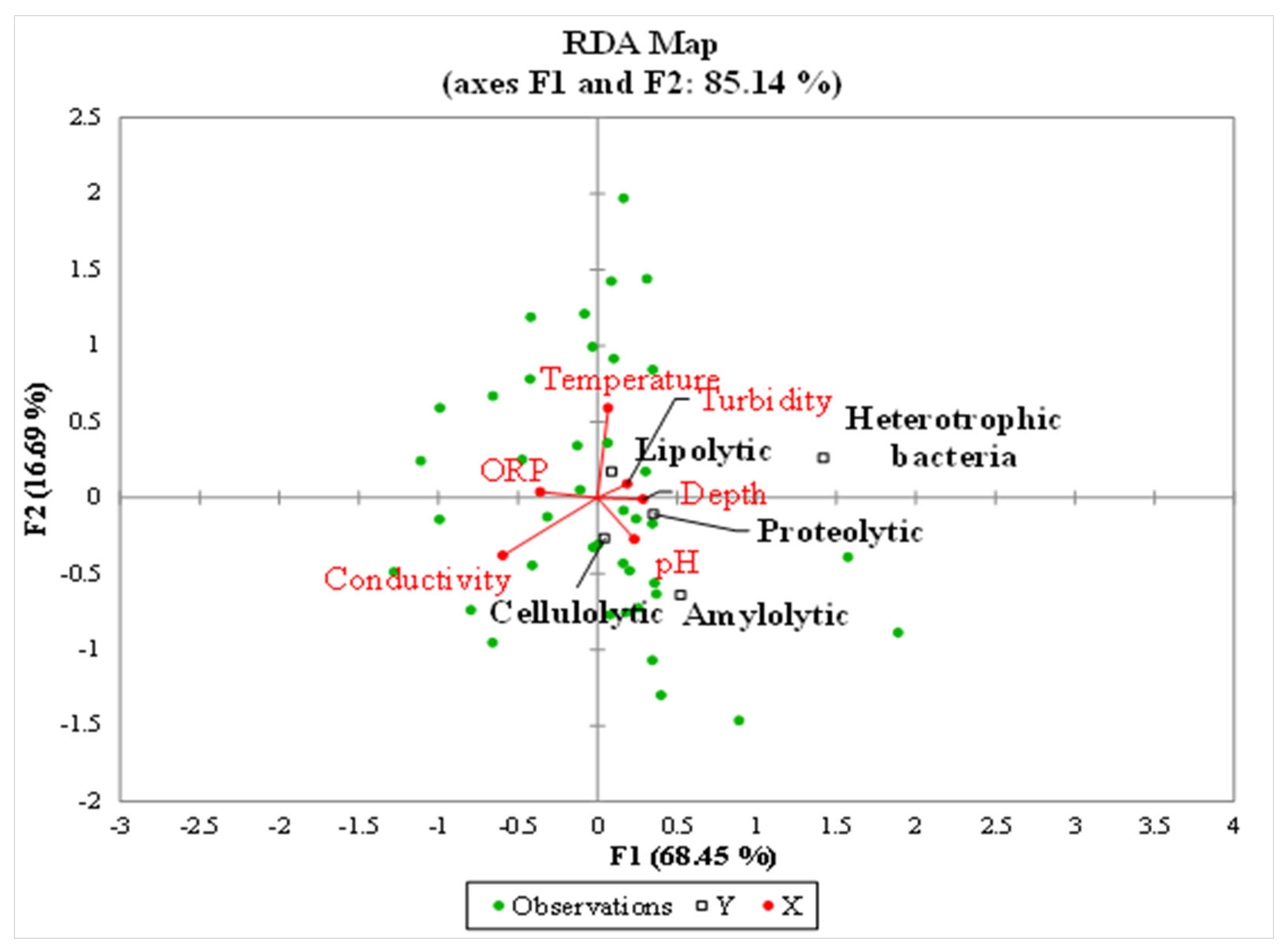
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azam, F.; Fenchel, T.; Field, J.; Gray, J.; Meyer-Reil, L.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Azam, F.; Smith, D.C.; Steward, G.F.; Hagström, Å. Bacteria-Organic Matter Coupling and Its Significance for Oceanic Carbon Cycling. Microb. Ecol. 1994, 28, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Chróst, R.J. Microbial Ectoenzymes in Aquatic Environments. In Aquatic Microbial Ecology; Overbeck, J., Chróst, R.J., Eds.; Brock/Springer Series in Contemporary Bioscience; Springer: New York, NY, USA, 1990; pp. 47–78. ISBN 978-1-4612-7991-4. [Google Scholar]
- Münster, U.; De Haan, H. The Role of Microbial Extracellular Enzymes in the Transformation of Dissolved Organic Matter in Humic Waters. In Aquatic Humic Substances; Hessen, D.O., Tranvik, L.J., Eds.; Ecological Studies; Springer Berlin Heidelberg: Berlin, Heidelberg, 1998; Volume 133, pp. 199–257. ISBN 978-3-642-08362-4. [Google Scholar]
- Shahid, M.J.; AL-surhanee, A.A.; Kouadri, F.; Ali, S.; Nawaz, N.; Afzal, M.; Rizwan, M.; Ali, B.; Soliman, M.H. Role of Microorganisms in the Remediation of Wastewater in Floating Treatment Wetlands: A Review. Sustainability 2020, 12, 5559. [Google Scholar] [CrossRef]
- Bullock, A.; Ziervogel, K.; Ghobrial, S.; Smith, S.; McKee, B.; Arnosti, C. A Multi-Season Investigation of Microbial Extracellular Enzyme Activities in Two Temperate Coastal North Carolina Rivers: Evidence of Spatial but Not Seasonal Patterns. Front. Microbiol. 2017, 8, 2589. [Google Scholar] [CrossRef]
- Arnosti, C.; Bell, C.; Moorhead, D.L.; Sinsabaugh, R.L.; Steen, A.D.; Stromberger, M.; Wallenstein, M.; Weintraub, M.N. Extracellular Enzymes in Terrestrial, Freshwater, and Marine Environments: Perspectives on System Variability and Common Research Needs. Biogeochemistry 2014, 117, 5–21. [Google Scholar] [CrossRef]
- Burns, R.G.; Dick, R.P. (Eds.) Enzymes in the Environment: Activity, Ecology, and Applications; Books in soils, plants, and the environment; Marcel Dekker: New York, NY, USA, 2002; ISBN 978-0-8247-0614-2. [Google Scholar]
- Tiquia, S.M. Extracellular Hydrolytic Enzyme Activities of the Heterotrophic Microbial Communities of the Rouge River: An Approach to Evaluate Ecosystem Response to Urbanization. Microb. Ecol. 2011, 62, 679–689. [Google Scholar] [CrossRef]
- Alves, P.D.D.; Siqueira, F.d.F.; Facchin, S.; Horta, C.C.R.; Victória, J.M.N.; Kalapothakis, E. Survey of Microbial Enzymes in Soil, Water, and Plant Microenvironments. TOMICROJ 2014, 8, 25–31. [Google Scholar] [CrossRef]
- Norashirene, M.J.; Rosliza, S.A.; Sharina, M.R.N.; Fithriah, M.A.N. Screening and Isolation of Lipolytic, Amylolytic and Cellulolytic Thermophiles from Hulu Langat Hot Spring, Selangor. In Proceedings of the 2012 IEEE Business, Engineering & Industrial Applications Colloquium (BEIAC), Kuala Lumpur, Malaysia, 7–8 April 2012; pp. 204–209. [Google Scholar]
- Cunha, A.; Almeida, A.; Coelho, F.J.R.C.; Gomes, N.C.M.; Oliveira, V.; Santos, A.L. Bacterial extracellular enzymatic activity in globally changing aquatic ecosystems. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; World Scientific Publishing Company: Singapore, 2010; Volume 1, pp. 124–135. [Google Scholar]
- Nybroe, O.; Jørgensen, P.E.; Henze, M. Enzyme Activities in Waste Water and Activated Sludge. Water Res. 1992, 26, 579–584. [Google Scholar] [CrossRef]
- Cârtsea, M. Geografical description of the 2nd district of Bucharest. PESD 2009, 3, 243–249. [Google Scholar]
- Stănescu, S.V.; Gavriloaie, C. Aspecte privind vegetația și fauna râului Colentina pe traseul din Municipiul București (România). Ecoterra 2011, 27, 49–52. [Google Scholar]
- Raducan, V.; Streza, I. Militant ephemeral in Mogosoaia ensemble. Horticulture 2014, LVIII, 359–368. [Google Scholar]
- Sanders, E.R. Aseptic Laboratory Techniques: Plating Methods. JoVE 2012, 63, 3064. [Google Scholar] [CrossRef] [PubMed]
- Ruginescu, R.M.; Cojoc, R.; Enache, M.; Lazăr, V. Preliminary Characterization of a Cellulase Producing Bacterial Strain Isolated from a Romanian Hypersaline Lake. JEP 2018, 09, 1066–1081. [Google Scholar] [CrossRef] [Green Version]
- Rohban, R.; Amoozegar, M.A.; Ventosa, A. Screening and Isolation of Halophilic Bacteria Producing Extracellular Hydrolyses from Howz Soltan Lake, Iran. J. Ind. Microbiol. Biotechnol. 2009, 36, 333–340. [Google Scholar] [CrossRef]
- Nick, T.G. Descriptive Statistics. In Topics in Biostatistics; Ambrosius, W.T., Ed.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2007; Volume 404, pp. 33–52. ISBN 978-1-58829-531-6. [Google Scholar]
- Nuzzo, R.L. The Box Plots Alternative for Visualizing Quantitative Data. PMR 2016, 8, 268–272. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK; Cambridge University Press: New York, NY, USA, 2002; ISBN 978-0-521-81128-6. [Google Scholar]
- Legendre, P.; Oksanen, J.; Ter Braak, C.J.F. Testing the Significance of Canonical Axes in Redundancy Analysis: Test of Canonical Axes in RDA. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Capblancq, T.; Luu, K.; Blum, M.G.B.; Bazin, E. Evaluation of Redundancy Analysis to Identify Signatures of Local Adaptation. Mol. Ecol. Resour. 2018, 18, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Hanashiro, F.T.T.; Mukherjee, S.; Souffreau, C.; Engelen, J.; Brans, K.I.; Busschaert, P.; De Meester, L. Freshwater Bacterioplankton Metacommunity Structure Along Urbanization Gradients in Belgium. Front. Microbiol. 2019, 10, 743. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, S.; Imai, A. Spatial and Temporal Variations in Sediment Enzyme Activities and Their Relationship with the Trophic Status of Erhai Lake. Ecol. Eng. 2015, 75, 365–369. [Google Scholar] [CrossRef]
- Song, H.; Xu, J.; Lavoie, M.; Fan, X.; Liu, G.; Sun, L.; Fu, Z.; Qian, H. Biological and Chemical Factors Driving the Temporal Distribution of Cyanobacteria and Heterotrophic Bacteria in a Eutrophic Lake (West Lake, China). Appl. Microbiol. Biotechnol. 2017, 101, 1685–1696. [Google Scholar] [CrossRef]
- Korzeniewska, E.; Brzozowska, R.; Czechowska, K.; Filipkowska, Z.; Niewolak, S. Seasonal Changes in the Number of Some Physiological Groups of Heterotrophic Bacteria in Water, Soil and Plants of the Wetlands near Olsztyn. Arch. Orchrony Sr. 2007, 33, 39–51. [Google Scholar]
- Mudryk, Z.; Donderski, W. The Occurrence of Heterotrophic Bacteria Decomposing Some Macromolecular Compounds in Shallow Estuarine Lakes. Hydrobiologia 1997, 342/343, 71–78. [Google Scholar] [CrossRef]
- Păceşilă, I. Extracellular Enzymatic Activities in the Aquatic Ecosystems of the Danube Delta. 1. β-Glucosidase Activity. Rom. Biotechnol. Lett. 2020, 25, 1908–1944. [Google Scholar] [CrossRef]
- Krstulovic, N.; Mladen, S. Distribution of Proteolityc, Amylolytic and Lipolityc Bacteria in the Kastela Bay. Acta Adriat. 1988, 29, 75–82. [Google Scholar]
- Souffreau, C.; Van der Gucht, K.; van Gremberghe, I.; Kosten, S.; Lacerot, G.; Lobão, L.M.; de Moraes Huszar, V.L.; Roland, F.; Jeppesen, E.; Vyverman, W.; et al. Environmental Rather than Spatial Factors Structure Bacterioplankton Communities in Shallow Lakes along a > 6000 Km Latitudinal Gradient in South America: Latitudinal Trends in Bacterioplankton Structure. Environ. Microbiol. 2015, 17, 2336–2351. [Google Scholar] [CrossRef]
- Câmara dos Reis, M.; Lacativa Bagatini, I.; de Oliveira Vidal, L.; Bonnet, M.-P.; da Motta Marques, D.; Sarmento, H. Correction: Spatial Heterogeneity and Hydrological Fluctuations Drive Bacterioplankton Community Composition in an Amazon Floodplain System. PLoS ONE 2020, 15, e0228721. [Google Scholar] [CrossRef] [Green Version]
- Chróst, R.J.; Siuda, W. Microbial Production, Utilization, and Enzymatic Degradation of Organic Matter in the Upper Trophogenic Layer in the Pelagial Zone of Lakes along a Eutrophication Gradient. Limnol. Oceanogr. 2006, 51, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Gutknecht, J.L.M.; Goodman, R.M.; Balser, T.C. Linking Soil Process and Microbial Ecology in Freshwater Wetland Ecosystems. Plant Soil 2006, 289, 17–34. [Google Scholar] [CrossRef]
- Menon, R.; Jackson, C.R.; Holland, M.M. The Influence of Vegetation on Microbial Enzyme Activity and Bacterial Community Structure in Freshwater Constructed Wetland Sediments. Wetlands 2013, 33, 365–378. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Minimum | Maximum | Mean | Std. Deviation |
|---|---|---|---|---|
| Depth (m) | 0.20 | 1.80 | 0.81 | 0.42 |
| Water velocity (m s−1) | 0.05 | 0.19 | 0.07 | 0.03 |
| Temperature (°C) | 7.42 | 29.60 | 17.37 | 6.41 |
| Transparency (m) | 0.20 | 1.00 | 0.48 | 0.18 |
| Turbidity (UM) | 8.00 | 94.60 | 19.65 | 13.59 |
| Conductivity (mS cm−1) | 387.00 | 710.00 | 510.23 | 86.78 |
| Salinity (UM) | 0.19 | 0.35 | 0.25 | 0.04 |
| Total dissolved solids | 194.00 | 355.00 | 255.00 | 43.44 |
| DO (mg O2L−1)-surface | 2.75 | 22.16 | 8.71 | 4.83 |
| DO (mg O2L−1)-bottom | 2.72 | 22.33 | 8.03 | 4.94 |
| DO (%) saturation-surface | 25.00 | 211.70 | 91.52 | 49.58 |
| DO (%) saturation-bottom | 24.00 | 212.50 | 82.85 | 47.94 |
| pH | 6.63 | 10.07 | 8.63 | 0.72 |
| ORP | −168.60 | 11.20 | −95.79 | 37.26 |
| Spring | Summer | Autumn | Winter | ||
|---|---|---|---|---|---|
| Colentina River | SP0 | 5.6 × 103 | 1.7 × 103 | 1.3 × 103 | 2.3 × 103 |
| Crevedia branch | SP1 | 3 × 102 | 3.6 × 103 | 4.6 × 103 | 1.6 × 103 |
| Mogoșoaia | SP2 | 3.5 × 102 | 1.1 × 103 | 1.2 × 106 | 1.5 × 103 |
| SP3 | 76.5 | 4.3 × 103 | 2.3 × 105 | 1.6 × 103 | |
| Plumbuita | SP4 | 5 × 102 | 3 × 103 | 1.6 × 103 | 1.7 × 104 |
| SP5 | 36 | 3.4 × 103 | 1.3 × 103 | 1.2 × 104 | |
| Fundeni | SP6 | 73.5 | 3.5 × 103 | 2.3 × 103 | 2.4 × 103 |
| SP7 | 75 | 5.9 × 102 | 6 × 103 | 7.7 × 103 | |
| Cernica | SP8 | 25 | 1.1 × 103 | 3.3 × 103 | 2.2 × 103 |
| SP9 | 10.3 × 102 | 10.3 × 103 | 7.5 × 103 | 1.5 × 102 |
| Contrast | Difference | Standardized Difference | Critical Value | Pr > Diff | Significant |
|---|---|---|---|---|---|
| winter vs. spring | 1.1027 | 3.4833 | 2.6933 | 0.0069 | Yes |
| winter vs. summer | 0.0238 | 0.0751 | 2.6933 | 0.9998 | No |
| autumn vs. spring | 1.6036 | 5.0658 | 2.6933 | <0.0001 | Yes |
| autumn vs. summer | 0.5247 | 1.6576 | 2.6933 | 0.3605 | No |
| autumn vs. winter | 0.5010 | 1.5825 | 2.6933 | 0.4011 | No |
| summer vs. spring | 1.0789 | 3.4082 | 2.6933 | 0.0085 | Yes |
| Statistic | Amylolytic | Proteolytic | Lipolytic | Cellulolytic |
|---|---|---|---|---|
| Minimum | 0 | 0 | 0 | 0 |
| Maximum | 150 | 45 | 2650 | 120 |
| Mean | 19.913 | 11.588 | 426.888 | 18.55 |
| Variance (n− 1) | 1286.345 | 184.922 | 259,498.814 | 759.715 |
| Standard deviation (n) | 35.414 | 13.428 | 503.002 | 27.216 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojovan, B.; Catana, R.; Neagu, S.; Cojoc, R.; Lucaci, A.I.; Marutescu, L.; Florescu, L.; Ruginescu, R.; Enache, M.; Moldoveanu, M. Metabolic Potential of Some Functional Groups of Bacteria in Aquatic Urban Systems. Fermentation 2021, 7, 242. https://doi.org/10.3390/fermentation7040242
Ojovan B, Catana R, Neagu S, Cojoc R, Lucaci AI, Marutescu L, Florescu L, Ruginescu R, Enache M, Moldoveanu M. Metabolic Potential of Some Functional Groups of Bacteria in Aquatic Urban Systems. Fermentation. 2021; 7(4):242. https://doi.org/10.3390/fermentation7040242
Chicago/Turabian StyleOjovan, Bianca, Rodica Catana, Simona Neagu, Roxana Cojoc, Anca Ioana Lucaci, Luminita Marutescu, Larisa Florescu, Robert Ruginescu, Madalin Enache, and Mirela Moldoveanu. 2021. "Metabolic Potential of Some Functional Groups of Bacteria in Aquatic Urban Systems" Fermentation 7, no. 4: 242. https://doi.org/10.3390/fermentation7040242
APA StyleOjovan, B., Catana, R., Neagu, S., Cojoc, R., Lucaci, A. I., Marutescu, L., Florescu, L., Ruginescu, R., Enache, M., & Moldoveanu, M. (2021). Metabolic Potential of Some Functional Groups of Bacteria in Aquatic Urban Systems. Fermentation, 7(4), 242. https://doi.org/10.3390/fermentation7040242