
Highly Stable, Cold-Active Aldehyde Dehydrogenase from the Marine Antarctic Flavobacterium sp. PL002


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning and Expression of F-ALDH Coding Gene
2.2. Purification of Recombinant F-ALDH and Size Exclusion Chromatography
2.3. Enzyme Assay
2.4. Biochemical Characterization of F-ALDH
2.5. Kinetic Parameters
2.6. Sequence Analyses and Phylogeny
3. Results
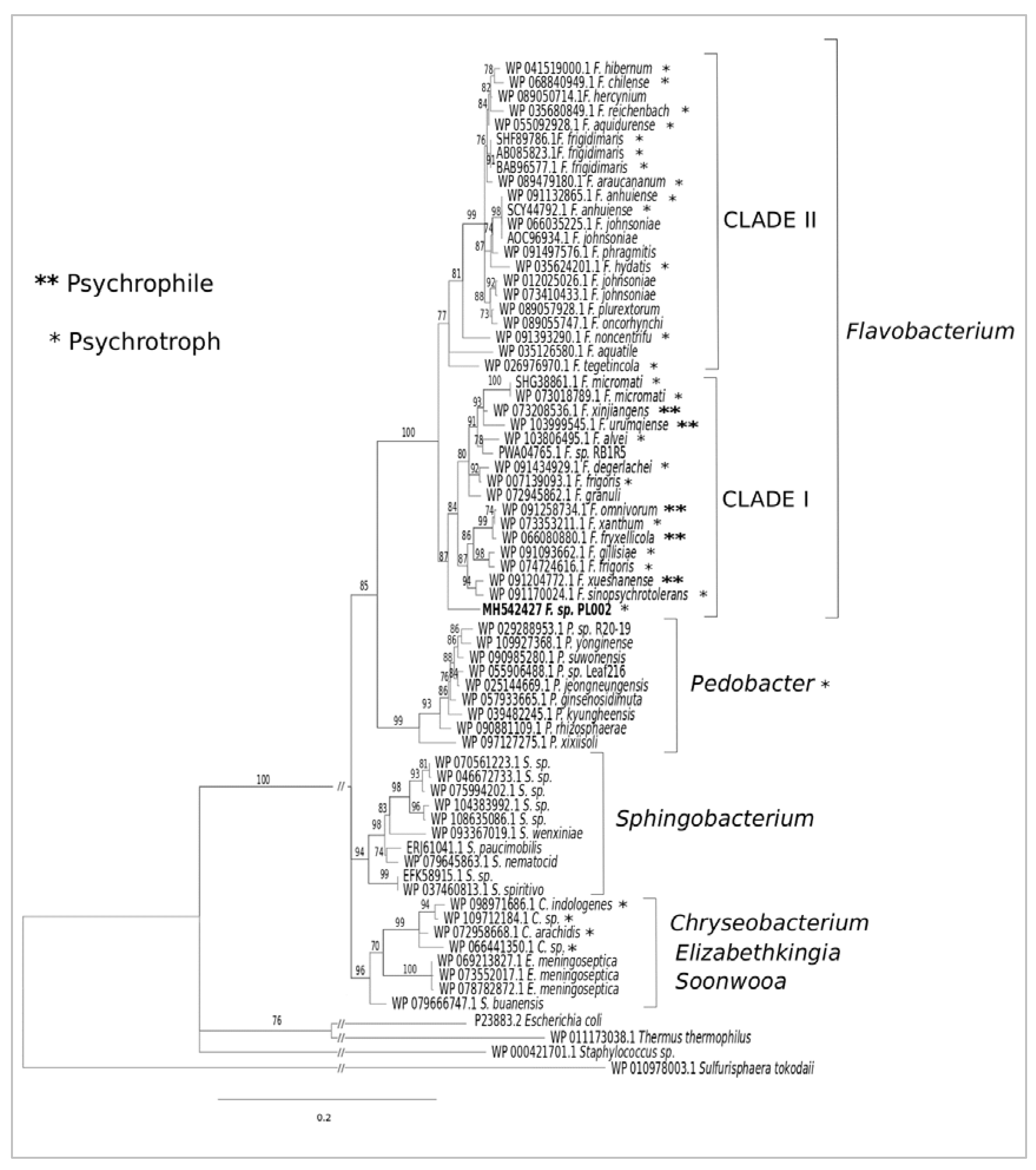
3.1. Phylogeny and Sequence Analyses of F-ALDH
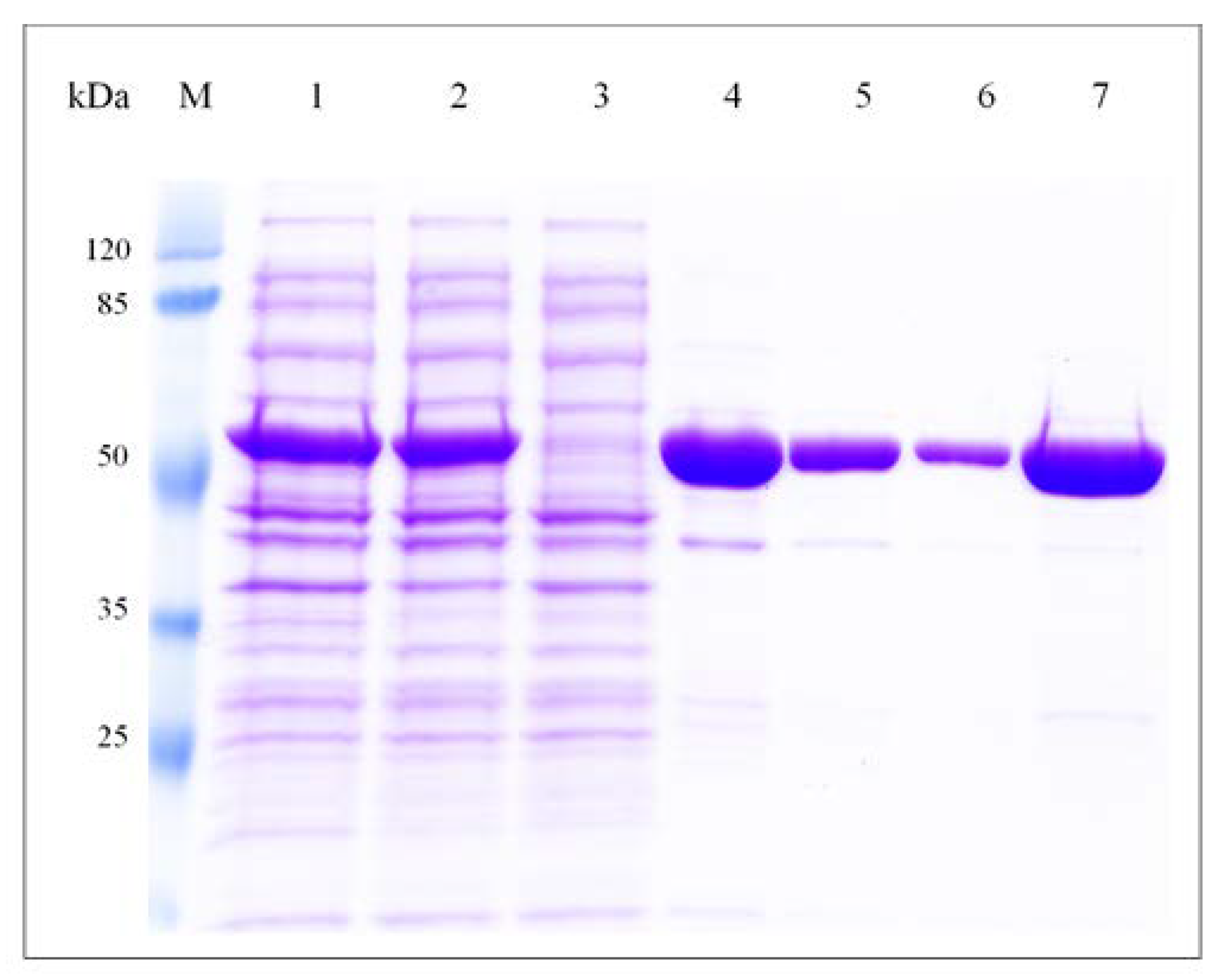
3.2. Cloning, Expression and Purification of F-ALDH
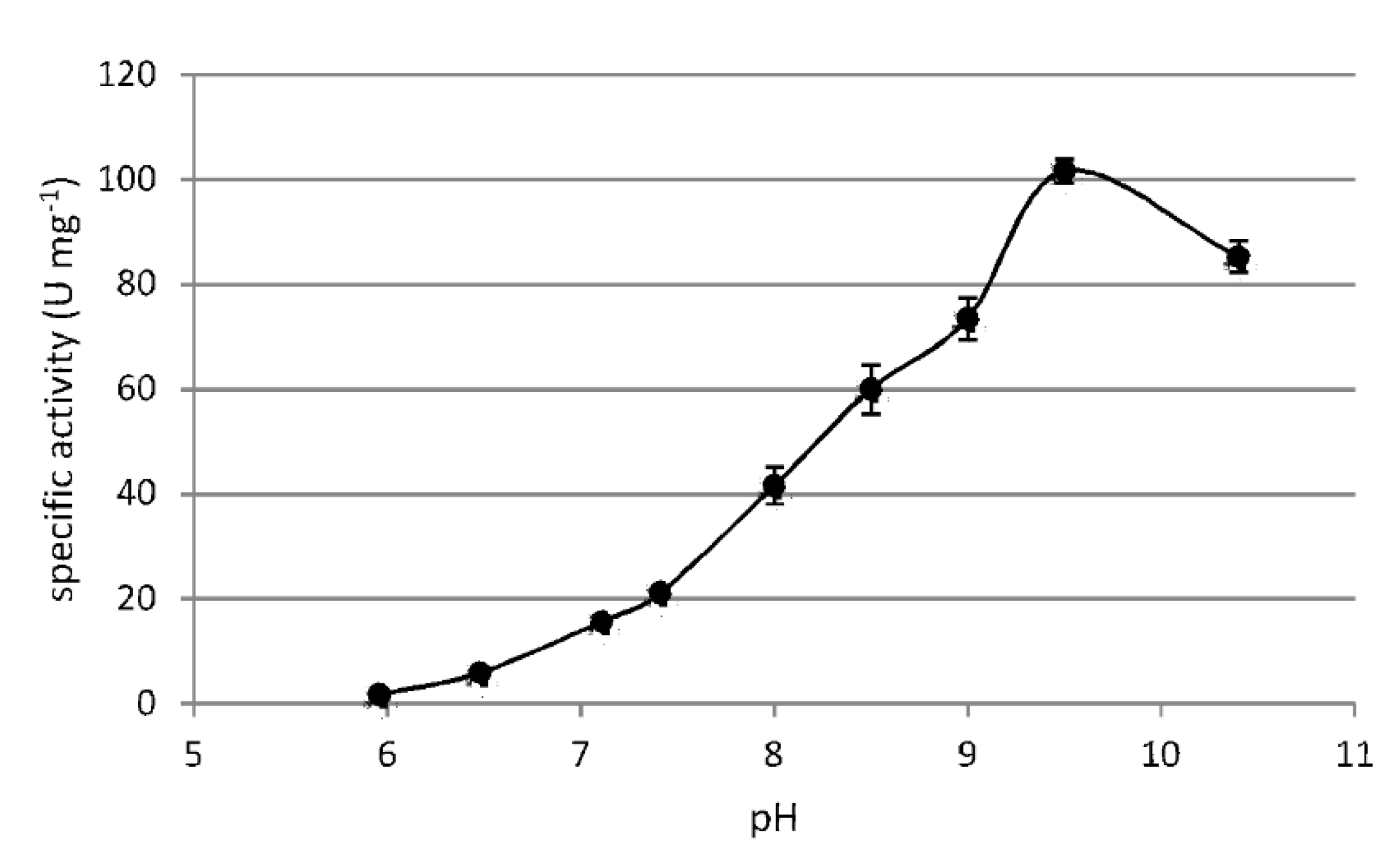
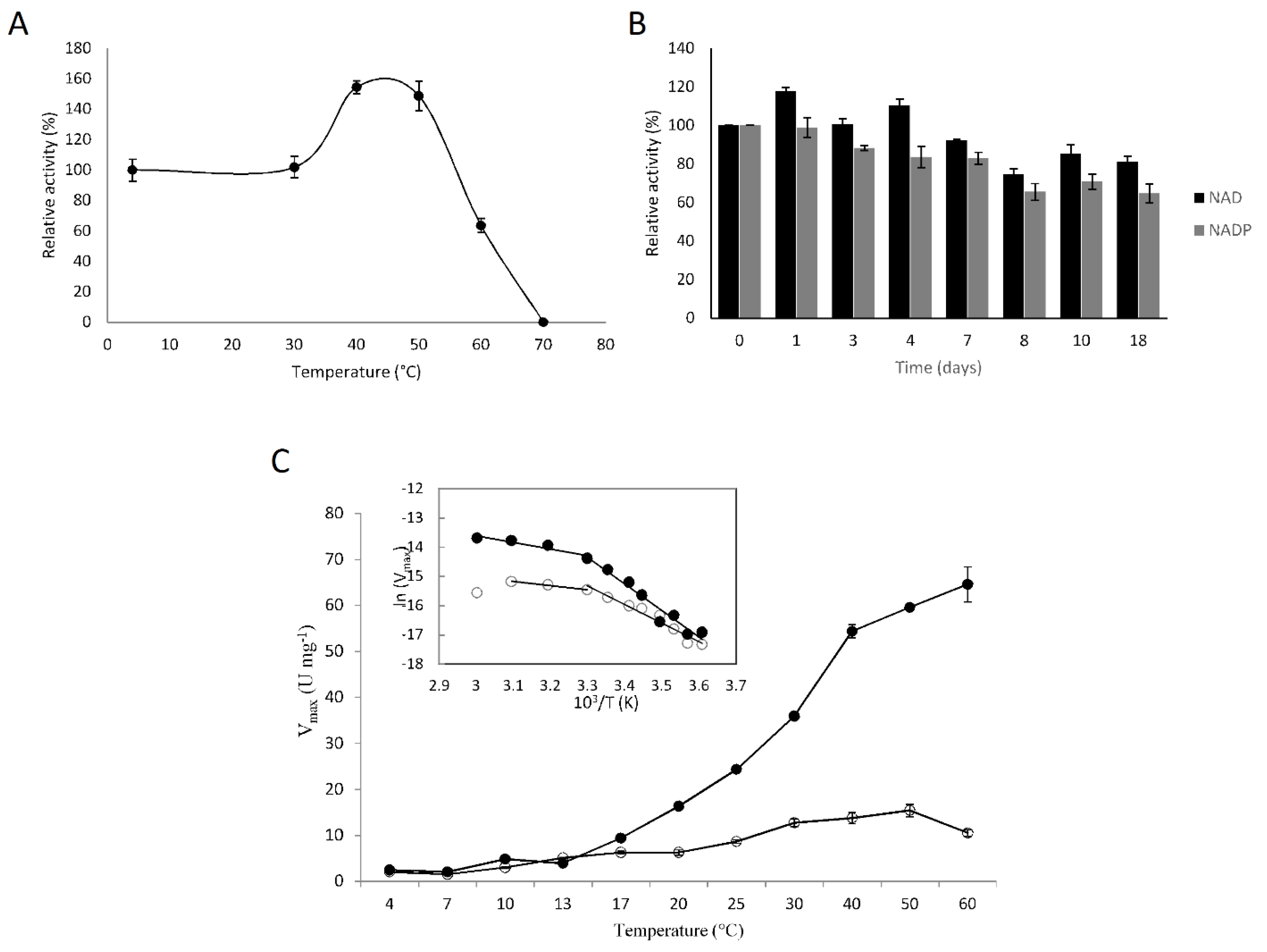
3.3. Biochemical Characterization of F-ALDH
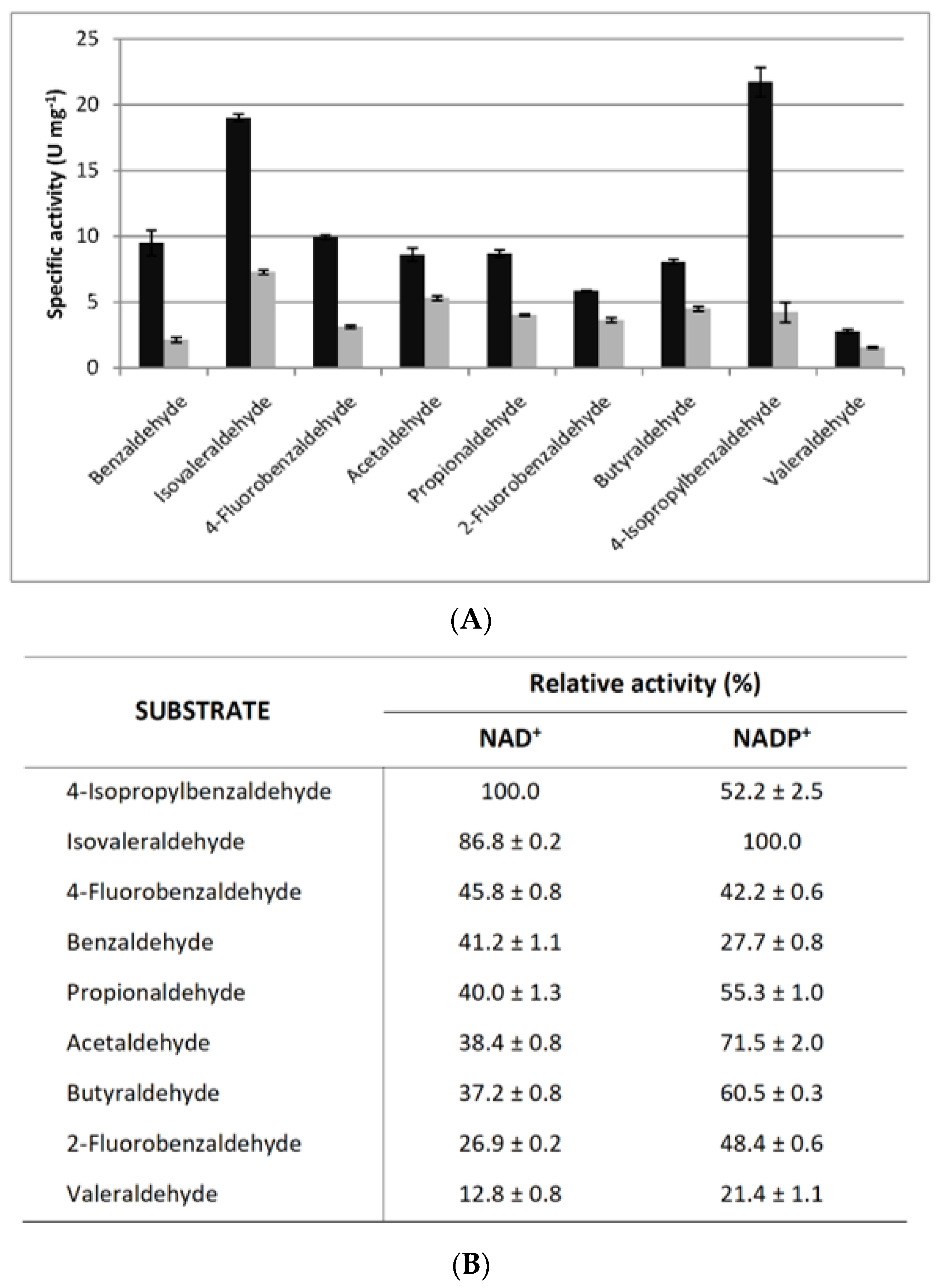
3.4. Kinetics of the Recombinant F-ALDH
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seckbach, J.O.A.S.-L.H. Polyextremophiles: Life under Multiple Forms of Stress; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, E.; Hornero-Méndez, D.; Azziz, G.; Lareo, C.; Saravia, V. Carotenoids from heterotrophic bacteria isolated from Fildes Peninsula, King George Island, Antarctica. Biotechnol. Rep. 2019, 21, e00306. [Google Scholar] [CrossRef] [PubMed]
- Paris, L.; Atala, C.; Gallardo-Cerda, J.; Gonzalez-Aravena, M.; Rodrigo De La, I.; Rómulo, O.; Torres-Díaz, C.; Trefault, N.; Molina-Montenegro, M.A.; IV, H.D.L. Isolation and characterization of an Antarctic Flavobacterium strain with agarase and alginate lyase activities. Pol. Polar Res. 2016, 37, 403–419. [Google Scholar]
- Williams, T.J.; Wilkins, D.; Long, E.; Evans, F.; DeMaere, M.Z.; Raftery, M.J.; Cavicchioli, R. The role of planktonic Flavobacteria in processing algal organic matter in coastal East Antarctica revealed using metagenomics and metaproteomics. Environ. Microbiol. 2013, 15, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Sharma, S. Enzymes in green chemistry: The need for environment and sustainability. Int. J. Appl. Res. 2016, 2, 337–341. [Google Scholar]
- Xu, F. Applications of oxidoreductases: Recent progress. Ind. Biotechnol. 2005, 1, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, K.S.; Cavicchioli, R. Cold-adapted enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Struvay, C.; Feller, G. Optimization to low temperature activity in psychrophilic enzymes. Int. J. Mol. Sci. 2012, 13, 11643–11665. [Google Scholar] [CrossRef] [Green Version]
- Georlette, D.; Blaise, V.; Collins, T.; D’Amico, S.; Gratia, E.; Hoyoux, A.; Marx, J.C.; Sonan, G.; Feller, G.; Gerday, C. Some like it cold: Biocatalysis at low temperatures. FEMS Microbiol. Rev. 2004, 28, 25–42. [Google Scholar] [CrossRef] [Green Version]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar] [CrossRef] [PubMed]
- Laybourn-Parry, J.; Pearce, D.A. The biodiversity and ecology of Antarctic lakes: Models for evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 2273–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Donato, P.; Buono, A.; Poli, A.; Finore, I.; Abbamondi, G.R.; Nicolaus, B.; Lama, L. Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses. Sustainability 2019, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Danilovich, M.; Sánchez, L.; Acosta, F.; Delgado, O. Antarctic bioprospecting: In pursuit of microorganisms producing new antimicrobials and enzymes. Polar Biol. 2018, 41, 1417–1433. [Google Scholar] [CrossRef]
- Santiago, M.; Ramírez-Sarmiento, C.A.; Zamora, R.A.; Parra, L.P. Discovery, Molecular Mechanisms, and Industrial Applications of Cold-Active Enzymes. Front. Microbiol. 2016, 7, 1408. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; Clarke, A. Antarctic marine biology. Curr. Biol. 2011, 21, R451–R457. [Google Scholar] [CrossRef] [Green Version]
- Lavin, P.; Henríquez-Castillo, C.; Yong, S.T.; Valenzuela-Heredia, D.; Oses, R.; Frez, K.; Borba, M.P.; Purcarea, C.; Wong, C. Draft Genome Sequence of Antarctic Psychrotroph Streptomyces fildesensis Strain INACH3013, Isolated from King George Island Soil. Microbiol. Resour. Announc. 2021, 10, e01453-20. [Google Scholar] [CrossRef]
- Muthusamy, C.; Sukumaran, R. Marine Microbial Enzymes. Biotechnology 2010, 9, 47–79. [Google Scholar]
- Bakunina, I.; Nedashkovskaya, O.; Kim, S.; Zvyagintseva, T.; Mikhailov, V. Diversity of glycosidase activities in the bacteria of the phylum Bacteroidetes isolated from marine algae. Microbiology 2012, 81. [Google Scholar] [CrossRef]
- Tropeano, M.; Coria, S.; Turjanski, A.; Cicero, D.; Bercovich, A.; Mac Cormack, W.; Vázquez, S. Culturable heterotrophic bacteria from Potter Cove, Antarctica, and their hydrolytic enzymes production. Polar Res. 2012, 31, 18507. [Google Scholar] [CrossRef]
- Nb, C.; Ruiz-Toquica, J.; Lopez-Kleine, L.; Montoya-Castano, D. Epiphytic Bacteria of Macroalgae of the Genus Ulva and Their Potential in Producing Enzymes Having Biotechnological Interest. J. Mar. Biol. Oceanogr. 2016, 5, 2–9. [Google Scholar] [CrossRef]
- Xie, M.; Li, J.; He, P.; Lin, X. Expression and characterization of a bifunctional alginate lyase named Al163 from the Antarctic bacterium Pseudoalteromonas sp. NJ-21. J. Oceanol. Limnol. 2018, 36, 1304–1314. [Google Scholar] [CrossRef]
- Marchitti, S.A.; Brocker, C.; Stagos, D.; Vasiliou, V. Non-P450 aldehyde oxidizing enzymes: The aldehyde dehydrogenase superfamily. Expert Opin. Drug Metab. Toxicol. 2008, 4, 697–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bains, J.; Boulanger, M.J. Structural and biochemical characterization of a novel aldehyde dehydrogenase encoded by the benzoate oxidation pathway in Burkholderia xenovorans LB400. J. Mol. Biol. 2008, 379, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Borisova, S.A.; Christman, H.D.; Metcalf, M.E.; Zulkepli, N.A.; Zhang, J.K.; van der Donk, W.A.; Metcalf, W.W. Genetic and biochemical characterization of a pathway for the degradation of 2-aminoethylphosphonate in Sinorhizobium meliloti 1021. J. Biol. Chem. 2011, 286, 22283–22290. [Google Scholar] [CrossRef] [Green Version]
- Jo, J.E.; Mohan Raj, S.; Rathnasingh, C.; Selvakumar, E.; Jung, W.C.; Park, S. Cloning, expression, and characterization of an aldehyde dehydrogenase from Escherichia coli K-12 that utilizes 3-Hydroxypropionaldehyde as a substrate. Appl. Microbiol. Biotechnol. 2008, 81, 51–60. [Google Scholar] [CrossRef]
- Zhou, S.; Catherine, C.; Rathnasingh, C.; Somasundar, A.; Park, S. Production of 3-hydroxypropionic acid from glycerol by recombinant Pseudomonas denitrificans. Biotechnol. Bioeng. 2013, 110, 3177–3187. [Google Scholar] [CrossRef]
- Ohta, T.; Tani, A.; Kimbara, K.; Kawai, F. A novel nicotinoprotein aldehyde dehydrogenase involved in polyethylene glycol degradation. Appl. Microbiol. Biotechnol. 2005, 68, 639–646. [Google Scholar] [CrossRef]
- Okibe, N.; Amada, K.; Hirano, S.; Haruki, M.; Imanaka, T.; Morikawa, M.; Kanaya, S. Gene cloning and characterization of aldehyde dehydrogenase from a petroleum-degrading bacterium, strain HD-1. J. Biosci. Bioeng. 1999, 88, 7–11. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Wei, D.; Li, P.; Wang, L.; Feng, L. Characterization of a broad-range aldehyde dehydrogenase involved in alkane degradation in Geobacillus thermodenitrificans NG80-2. Microbiol. Res. 2010, 165, 706–712. [Google Scholar] [CrossRef]
- Kato, T.; Miyanaga, A.; Kanaya, S.; Morikawa, M. Gene cloning and characterization of an aldehyde dehydrogenase from long-chain alkane-degrading Geobacillus thermoleovorans B23. Extremophiles 2010, 14, 33–39. [Google Scholar] [CrossRef]
- Avramescu, A.; Andreescu, S.; Noguer, T.; Bala, C.; Andreescu, D.; Marty, J.L. Biosensors designed for environmental and food quality control based on screen-printed graphite electrodes with different configurations. Anal. Bioanal. Chem. 2002, 374, 25–32. [Google Scholar] [CrossRef]
- Badalyan, A.; Dierich, M.; Stiba, K.; Schwuchow, V.; Leimkühler, S.; Wollenberger, U. Electrical Wiring of the Aldehyde Oxidoreductase PaoABC with a Polymer Containing Osmium Redox Centers: Biosensors for Benzaldehyde and GABA. Biosensors 2014, 4, 403–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badalyan, A.; Neumann-Schaal, M.; Leimkühler, S.; Wollenberger, U. A Biosensor for Aromatic Aldehydes Comprising the Mediator Dependent PaoABC-Aldehyde Oxidoreductase. Electroanalysis 2013, 25, 101–108. [Google Scholar] [CrossRef]
- Ghica, M.E.; Pauliukaite, R.; Marchand, N.; Devic, E.; Brett, C.M. An improved biosensor for acetaldehyde determination using a bienzymatic strategy at poly(neutral red) modified carbon film electrodes. Anal. Chim. Acta. 2007, 591, 80–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iitani, K.; Chien, P.-J.; Suzuki, T.; Toma, K.; Arakawa, T.; Iwasaki, Y.; Mitsubayashi, K. Improved Sensitivity of Acetaldehyde Biosensor by Detecting ADH Reverse Reaction-Mediated NADH Fluoro-Quenching for Wine Evaluation. ACS Sens. 2017, 2, 940–946. [Google Scholar] [CrossRef]
- Pariente, F.; Lorenzo, E.; Tobalina, F.; Abruna, H.D. Aldehyde Biosensor Based on the Determination of NADH Enzymically Generated by Aldehyde Dehydrogenase. Anal. Chem. 1995, 67, 3936–3944. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Kazuoka, T.; Yoshida, M.; Yamanaka, K.; Oikawa, T.; Soda, K. Thermostable aldehyde dehydrogenase from psychrophile, Cytophaga sp. KUC-1: Enzymological characteristics and functional properties. Biochem. Bioph. Res. Commun. 2002, 298, 632–637. [Google Scholar] [CrossRef]
- Teoh, C.P.; Lavin, P.; Najimudin, N.; Lee, P.C.; Iancu, L.; Purcarea, C.; Wong, C. Draft Genome Sequence of Flavobacterium sp. Strain PL002, Isolated from Antarctic Porphyra Algae. Microbiol. Resour. Announc. 2021, 10, e00063-21. [Google Scholar] [CrossRef]
- Reasoner, D.J.; Geldreich, E.E. A new medium for the enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Purcarea, C.; Ahuja, A.; Lu, T.; Kovari, L.; Guy, H.I.; Evans, D.R. Aquifex aeolicus aspartate transcarbamoylase, an enzyme specialized for the efficient utilization of unstable carbamoyl phosphate at elevated temperature. J. Biol. Chem. 2003, 278, 52924–52934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Joo, J.C.; Brown, G.; Stolnikova, E.; Halavaty, A.S.; Savchenko, A.; Anderson, W.F.; Yakunin, A.F. Structure-based mutational studies of substrate inhibition of betaine aldehyde dehydrogenase BetB from Staphylococcus aureus. Appl. Environ. Microbiol. 2014, 80, 3992–4002. [Google Scholar] [CrossRef] [Green Version]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Van Trappen, S.; Vandecandelaere, I.; Mergaert, J.; Swings, J. Flavobacterium degerlachei sp. nov., Flavobacterium frigoris sp. nov. and Flavobacterium micromati sp. nov., novel psychrophilic bacteria isolated from microbial mats in Antarctic lakes. Int. J. Syst. Evol. Microbiol. 2004, 54, 85–92. [Google Scholar] [CrossRef]
- Aslam, Z.; Im, W.T.; Kim, M.K.; Lee, S.T. Flavobacterium granuli sp. nov., isolated from granules used in a wastewater treatment plant. Int. J. Syst. Evol. Microbiol. 2005, 55, 747–751. [Google Scholar] [CrossRef] [Green Version]
- McCammon, S.A.; Bowman, J.P. Taxonomy of Antarctic Flavobacterium species: Description of Flavobacterium gillisiae sp. nov., Flavobacterium tegetincola sp. nov., and Flavobacterium xanthum sp. nov., nom. rev. and reclassification of [Flavobacterium] salegens as Salegentibacter salegens gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 3, 1055–1063. [Google Scholar] [CrossRef]
- Lee, Y.; Jeon, C.O. Flavobacterium alvei sp. nov., isolated from a freshwater river. Int. J. Syst. Evol. Microbiol. 2018, 68, 1919–1924. [Google Scholar] [CrossRef]
- Dong, K.; Liu, H.; Zhang, J.; Zhou, Y.; Xin, Y. Flavobacterium xueshanense sp. nov. and Flavobacterium urumqiense sp. nov., two psychrophilic bacteria isolated from glacier ice. Int. J. Syst. Evol. Microbiol. 2012, 62, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Heim, R.; Strehler, E.E. Cloning an Escherichia coli gene encoding a protein remarkably similar to mammalian aldehyde dehydrogenases. Gene 1991, 99, 15–23. [Google Scholar] [CrossRef]
- Hayes, K.; Noor, M.R.; Djeghader, A.; Armshaw, P.; Pembroke, T.; Tofail, S.A.M.; Soulimane, T. The quaternary structure of Thermus thermophilus aldehyde dehydrogenase is stabilized by an evolutionary distinct C-terminal arm extension. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Hao, L.; Wang, R.; Liu, B. Molecular characterization of a thermostable aldehyde dehydrogenase (ALDH) from the hyperthermophilic archaeon Sulfolobus tokodaii strain 7. Extremophiles 2013, 17, 181–190. [Google Scholar] [CrossRef]
- d Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Hempel, J.; Lindahl, R.; Perozich, J.; Wang, B.; Kuo, I.; Nicholas, H. Beyond the catalytic core of ALDH: A web of important residues begins to emerge. Chem. Biol. Interact. 2001, 130–132, 39–46. [Google Scholar] [CrossRef]
- Moore, S.A.; Baker, H.M.; Blythe, T.J.; Kitson, K.E.; Kitson, T.M.; Baker, E.N. Sheep liver cytosolic aldehyde dehydrogenase: The structure reveals the basis for the retinal specificity of class 1 aldehyde dehydrogenases. Structure 1998, 6, 1541–1551. [Google Scholar] [CrossRef] [Green Version]
- Perozich, J.; Nicholas, H.; Wang, B.C.; Lindahl, R.; Hempel, J. Relationships within the aldehyde dehydrogenase extended family. Protein Sci. 1999, 8, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Clares, R.A.; Díaz-Sánchez, A.G.; González-Segura, L.; Montiel, C. Kinetic and structural features of betaine aldehyde dehydrogenases: Mechanistic and regulatory implications. Arch. Biochem. Biophys. 2010, 493, 71–81. [Google Scholar] [CrossRef]
- Halavaty, A.S.; Rich, R.L.; Chen, C.; Joo, J.C.; Minasov, G.; Dubrovska, I.; Winsor, J.R.; Myszka, D.G.; Duban, M.; Shuvalova, L.; et al. Structural and functional analysis of betaine aldehyde dehydrogenase from Staphylococcus aureus. Acta Crystallogr. D Biol. Crystallogr. 2015, 71, 1159–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, R. Aldehyde dehydrogenases and their role in carcinogenesis. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 283–335. [Google Scholar] [CrossRef]
- Shortall, K.; Djeghader, A.; Magner, E.; Soulimane, T. Insights into Aldehyde Dehydrogenase Enzymes: A Structural Perspective. Front. Mol. Biosci. 2021, 8, 410. [Google Scholar] [CrossRef] [PubMed]
- Smit, B.; Engels, W.; Smit, G. Branched chain aldehydes: Production and breakdown pathways and relevance for flavour in foods. Appl. Microbiol. Biotechnol. 2008, 81, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, A.; Taunton, J. Lysine-Targeted Inhibitors and Chemoproteomic Probes. Annu. Rev. Biochem. 2019, 88, 365–381. [Google Scholar] [CrossRef]
- Reichardt, P.; Schreiber, A.; Wichmann, G.; Metzner, G.; Efer, J.; Raabe, F. Identification and quantification of in vitro adduct formation between protein reactive xenobiotics and a lysine-containing model peptide. Environ. Toxicol. 2003, 18, 29–36. [Google Scholar] [CrossRef]
- Giebułtowicz, J.; Wolinowska, R.; Sztybor, A.; Pietrzak, M.; Wroczyński, P.; Wierzchowski, J. Salivary aldehyde dehydrogenase: Activity towards aromatic aldehydes and comparison with recombinant ALDH3A1. Molecules 2009, 14, 2363–2372. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Zavala, J.S.; Allali-Hassani, A.; Weiner, H. Characterization of E. coli tetrameric aldehyde dehydrogenases with atypical properties compared to other aldehyde dehydrogenases. Protein Sci. 2006, 15, 1387–1396. [Google Scholar] [CrossRef] [Green Version]
- Van Ophem, P.W.; Duine, J.A. Different types of formaldehyde-oxidizing dehydrogenases in Nocardia species 239: Purification and characterization of an NAD-dependent aldehyde dehydrogenase. Arch. Biochem. Biophys. 1990, 282, 248–253. [Google Scholar] [CrossRef]
- Brocker, C.; Lassen, N.; Estey, T.; Pappa, A.; Cantore, M.; Orlova, V.V.; Chavakis, T.; Kavanagh, K.L.; Oppermann, U.; Vasiliou, V. Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel enzyme involved in cellular defense against hyperosmotic stress. J. Biol. Chem. 2010, 285, 18452–18463. [Google Scholar] [CrossRef] [Green Version]
- McLeish, M.J.; Kneen, M.M.; Gopalakrishna, K.N.; Koo, C.W.; Babbitt, P.C.; Gerlt, J.A.; Kenyon, G.L. Identification and Characterization of a Mandelamide Hydrolase and an NAD(P)+-Dependent Benzaldehyde Dehydrogenase from Pseudomonas putida ATCC 12633. J. Bacteriol. 2003, 185, 2451–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagos, D.; Chen, Y.; Brocker, C.; Donald, E.; Jackson, B.C.; Orlicky, D.J.; Thompson, D.C.; Vasiliou, V. Aldehyde dehydrogenase 1B1: Molecular cloning and characterization of a novel mitochondrial acetaldehyde-metabolizing enzyme. Drug Metab. Dispos. 2010, 38, 1679–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klyosov, A.A. Kinetics and Specificity of Human Liver Aldehyde Dehydrogenases toward Aliphatic, Aromatic, and Fused Polycyclic Aldehydes. Biochemistry 1996, 35, 4457–4467. [Google Scholar] [CrossRef]
- Lindahl, R.; Evces, S. Rat liver aldehyde dehydrogenase. I. Isolation and characterization of four high Km normal liver isozymes. J. Biol. Chem. 1984, 259, 11986–11990. [Google Scholar] [CrossRef]
- Eggert, M.W.; Byrne, M.E.; Chambers, R.P. Kinetic involvement of acetaldehyde substrate inhibition on the rate equation of yeast aldehyde dehydrogenase. Appl. Biochem. Biotechnol. 2012, 168, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Tuck, L.R.; Altenbach, K.; Ang, T.F.; Crawshaw, A.D.; Campopiano, D.J.; Clarke, D.J.; Marles-Wright, J. Insight into Coenzyme A cofactor binding and the mechanism of acyl-transfer in an acylating aldehyde dehydrogenase from Clostridium phytofermentans. Sci. Rep. 2016, 6, 22108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedino, S.; Testore, G.; Obert, F. Initial characterization of aldehyde dehydrogenase from rat testis cytosol. Biol. Chem. Hoppe Seyler 1990, 371, 95–101. [Google Scholar] [CrossRef]
- Titoiu, A.M.; Necula-Petrareanu, G.; Visinescu, D.; Dinca, V.; Bonciu, A.; Mihailescu, C.N.; Purcarea, C.; Boukherroub, R.; Szunerits, S.; Vasilescu, A. Flow injection enzymatic biosensor for aldehydes based on a Meldola Blue-Ni complex electrochemical mediator. Microchim. Acta 2020, 187, 550. [Google Scholar] [CrossRef]
- Bucur, B.; Munteanu, F.; Marty, J.-L.; Vasilescu, A. Advances in Enzyme-Based Biosensors for Pesticide Detection. Biosensors 2018, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Fanjul-Bolado, P.; Fogel, R.; Limson, J.; Purcarea, C.; Vasilescu, A. Advances in the Detection of Dithiocarbamate Fungicides: Opportunities for Biosensors. Biosensors 2021, 11, 12. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, R.A.; Basso, A.; Brady, D. New frontiers in enzyme immobilisation: Robust biocatalysts for a circular bio-based economy. Chem. Soc. Rev. 2021, 50, 5850–5862. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-J.; Li, R.-F.; Li, X.-Y.; Zhang, Y.-W. One-step selective affinity purification and immobilization of His-tagged enzyme by recyclable magnetic nanoparticles. Eng. Life Sci. 2021, 21, 364–373. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Li, G.; Zhang, Y.; Zhang, T.; Liu, X.; Gao, F.; Peng, M.; He, Y.; Fan, H. Recent Advances in Enzyme-Nanostructure Biocatalysts with Enhanced Activity. Catalysts 2020, 10, 338. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, Y.; Yang, C.; Ma, C.; Tang, J. Enzyme-inorganic hybrid nanoflowers: Classification, synthesis, functionalization and potential applications. Chem. Eng. J. 2021, 415, 129075. [Google Scholar] [CrossRef]
- Wang, X.; Saba, T.; Yiu, H.H.P.; Howe, R.F.; Anderson, J.A.; Jiafu, S. Cofactor NAD(P)H Regeneration Inspired by Heterogeneous Pathways. Chem 2017, 2, 621–654. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ENZYME | Identity (%) | ||||||
| F-ALDH | Ff-ALDH | Ec-ALDH | Sa-ALDH | Tt-ALDH | St-ALDH | ||
| F-ALDH | Similarity (%) | 89 | 38 | 38 | 39 | 30 | |
| Ff-ALDH | 94 | 38 | 38 | 39 | 30 | ||
| Ec-ALDH | 56 | 57 | 37 | 42 | 33 | ||
| Sa-ALDH | 60 | 59 | 58 | 37 | 34 | ||
| Tt-ALDH | 58 | 57 | 60 | 57 | 33 | ||
| St-ALDH | 52 | 52 | 56 | 58 | 54 | ||
| Compound | Relative Activity (%) |
|---|---|
| None | 100 |
| KCl (100 mM) | 104.5 |
| NaCl (100 mM) | 102.2 |
| KCl (100 mM) * | 99.6 |
| NaCl (100 mM) * | 111.3 |
| CaCl2 (1 mM) | 93.5 |
| MgSO4 (1 mM) | 91.2 |
| MgCl2 (1 mM) | 84.4 |
| NiCl2 (1 mM) | 78.6 |
| HgCl2 (0.5 mM) | 0 |
| HgCl2 (1 mM) | 0 |
| Ethanol (1%) | 85.0 |
| Ethanol (10%) | 144.7 |
| EDTA (2 mM) | 81.7 |
| Triton X-100 | 90.1 |
| β-ME (1 mM) | 116.1 |
| β-ME (10 mM) | 124.1 |
| Protease inhibitors (1X) | 78.2 |
| Variable Substrate | Second Substrate | KM (µM) | Vmax (µmol min−1 mg−1) | kcat (s−1) | Ki (mM) | kcat / KM (μM−1 s−1) |
|---|---|---|---|---|---|---|
| Isovaleraldehyde | NAD+ | 197.0 ± 39.1 | 36.35 ± 3.26 | 100.04 ± 8.98 | 2.08 ± 0.40 | 0.50 ± 0.22 |
| NADP+ | 70.6 ± 6.8 | 12.40 ± 0.39 | 34.11± 1.08 | 2.96 ± 0.28 | 0.48 ± 0.15 | |
| Benzaldehyde | NAD+ | 136.8 ± 15.9 | 12.63 ± 0.40 | 34.74 ± 1.12 | - | 0.25 ± 0.07 |
| NADP+ | 57.5 ± 14.8 | 2.90 ± 0.20 | 10.72 ± 0.68 | 15.33 ± 5.7 | 0.14 ± 0.08 | |
| NAD+ | Isovaleraldehyde | 74.0 ± 6.2 | 21.32 ± 0.32 | 78.7 ± 1.53 | - | 1.06 ± 0.24 |
| Benzaldehyde | 32.5 ± 3.2 | 7.97 ± 0.15 | 29.24 ± 0.54 | - | 0.89 + 0.17 | |
| NADP+ | Isovaleraldehyde | 1419.0 ± 139.3 | 19.13 ± 0.75 | 70.16 ± 2.75 | - | 0.05 ± 0.02 |
| Benzaldehyde | 430.8 ± 105.0 | 2.40 ± 0.14 | 8.96 ± 0.54 | - | 0.02 + 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Necula-Petrareanu, G.; Lavin, P.; Paun, V.I.; Gheorghita, G.R.; Vasilescu, A.; Purcarea, C. Highly Stable, Cold-Active Aldehyde Dehydrogenase from the Marine Antarctic Flavobacterium sp. PL002. Fermentation 2022, 8, 7. https://doi.org/10.3390/fermentation8010007
Necula-Petrareanu G, Lavin P, Paun VI, Gheorghita GR, Vasilescu A, Purcarea C. Highly Stable, Cold-Active Aldehyde Dehydrogenase from the Marine Antarctic Flavobacterium sp. PL002. Fermentation. 2022; 8(1):7. https://doi.org/10.3390/fermentation8010007
Chicago/Turabian StyleNecula-Petrareanu, Georgiana, Paris Lavin, Victoria Ioana Paun, Giulia Roxana Gheorghita, Alina Vasilescu, and Cristina Purcarea. 2022. "Highly Stable, Cold-Active Aldehyde Dehydrogenase from the Marine Antarctic Flavobacterium sp. PL002" Fermentation 8, no. 1: 7. https://doi.org/10.3390/fermentation8010007
APA StyleNecula-Petrareanu, G., Lavin, P., Paun, V. I., Gheorghita, G. R., Vasilescu, A., & Purcarea, C. (2022). Highly Stable, Cold-Active Aldehyde Dehydrogenase from the Marine Antarctic Flavobacterium sp. PL002. Fermentation, 8(1), 7. https://doi.org/10.3390/fermentation8010007