
Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications


Abstract
:1. Introduction
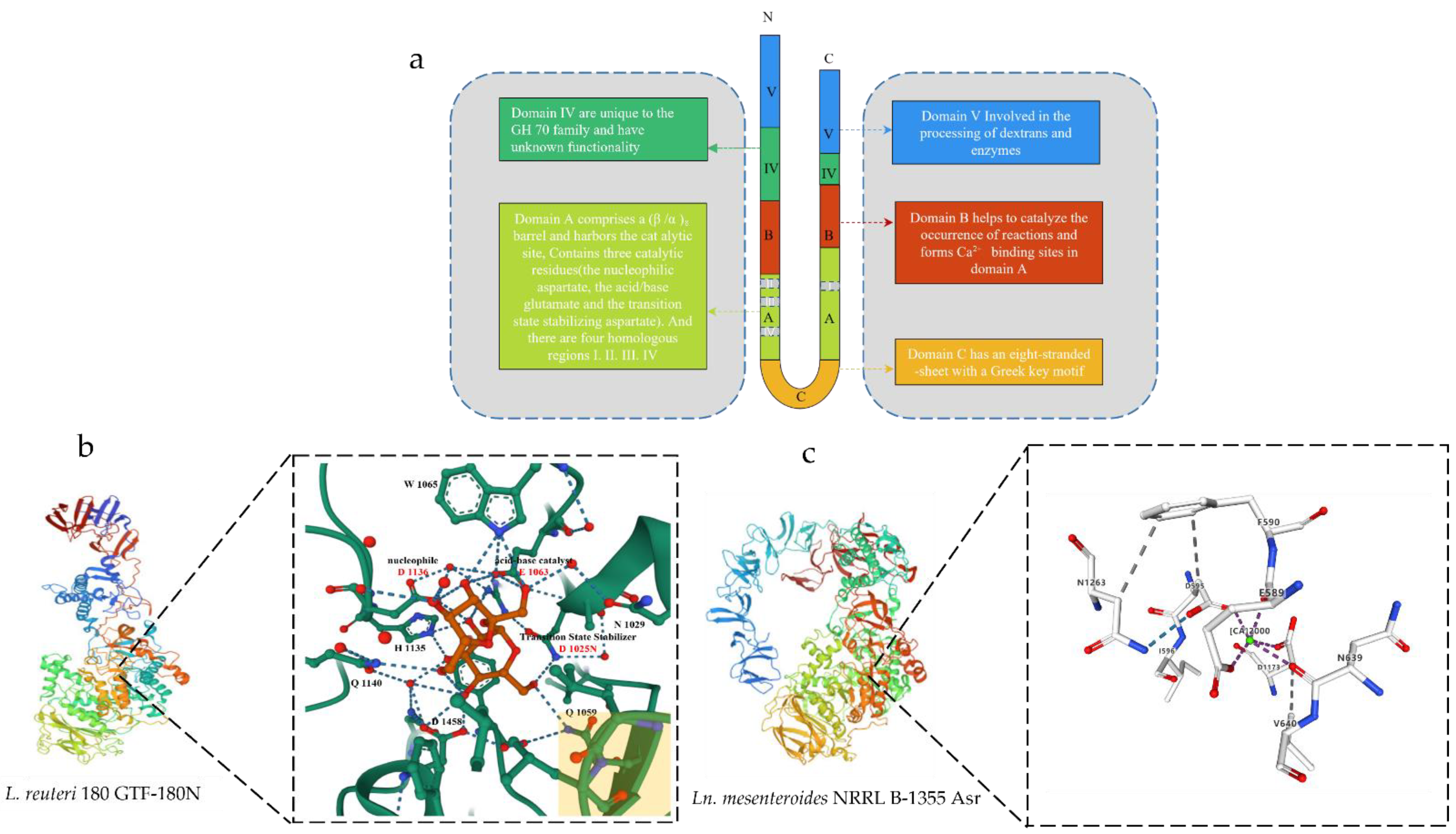
2. Structure of the GS
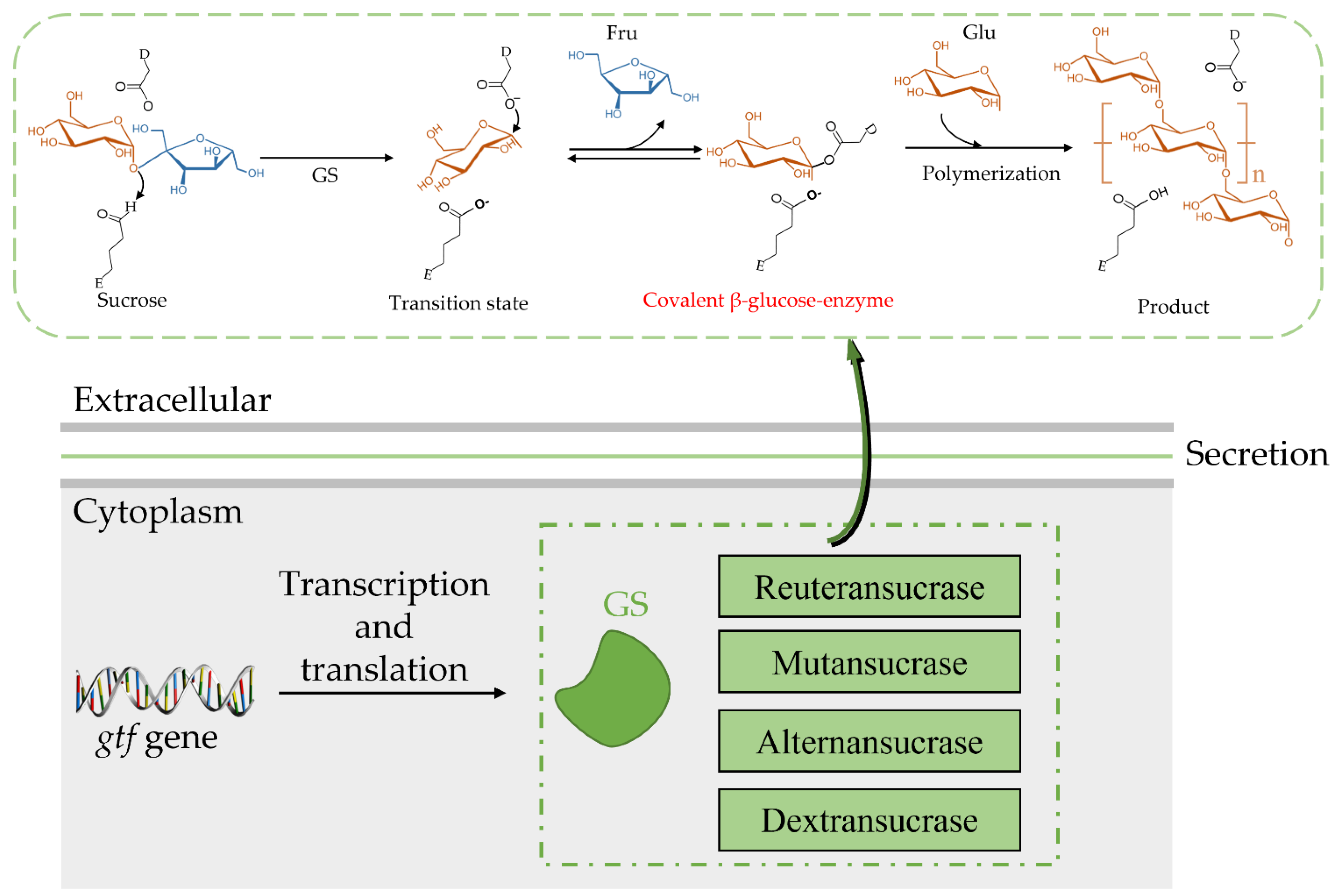
3. Catalytic Mechanism of GS
4. Isolation and Purification of GS
5. Physiological and Biochemical Properties of GS
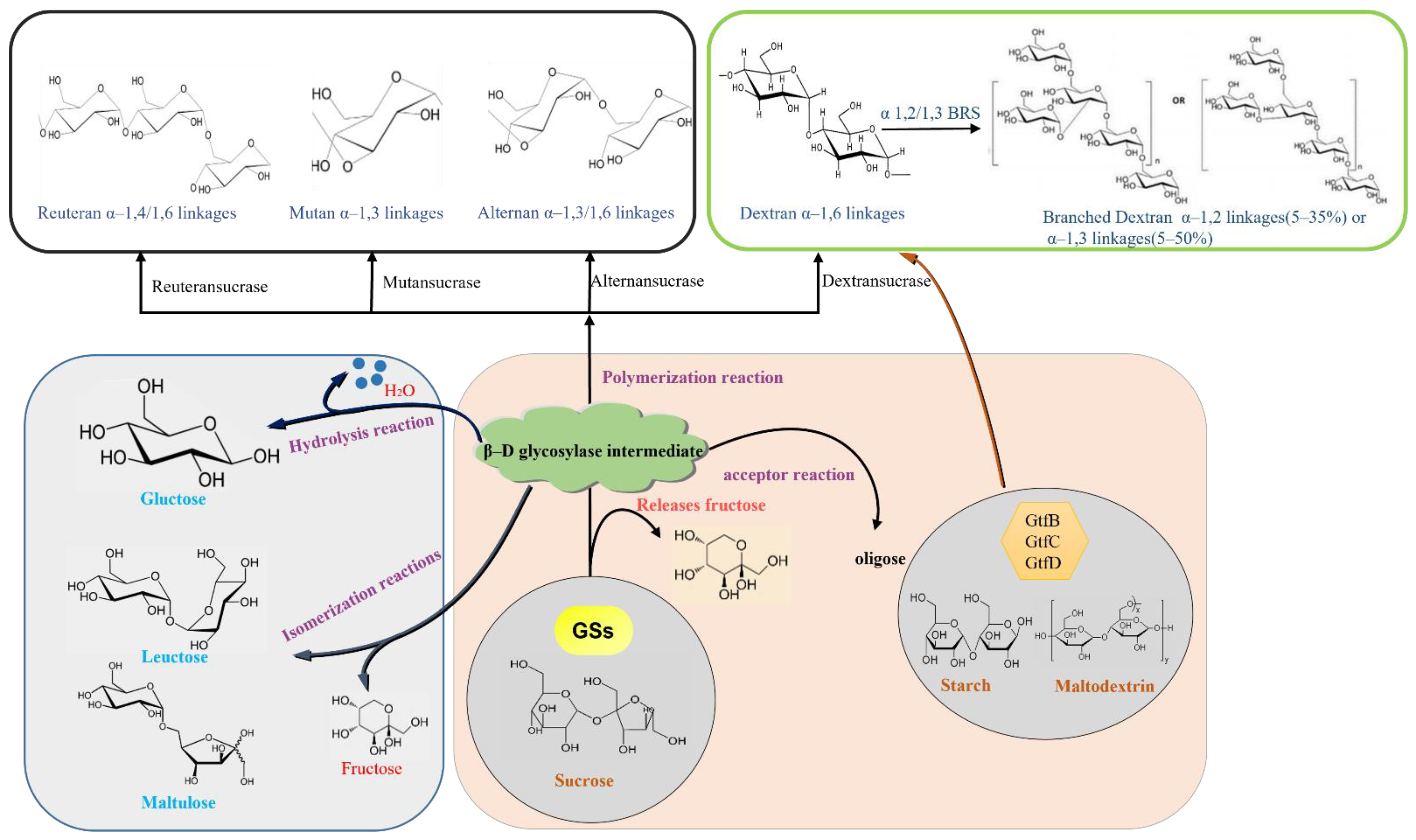
6. EPS Synthesis by GS
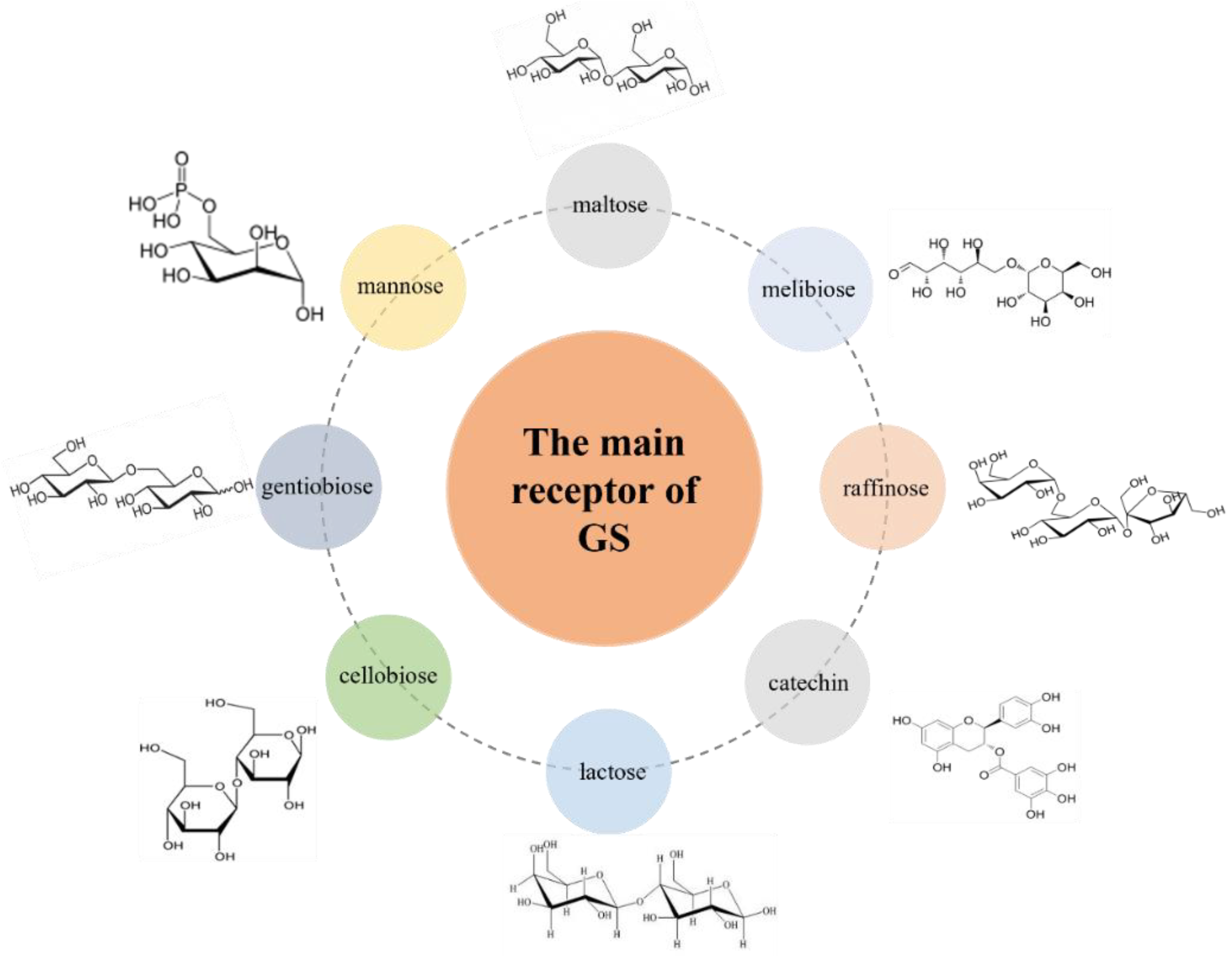
7. Acceptor Reaction
7.1. Acceptor Reaction with Maltose
7.2. Acceptor Reaction with Mannose
7.3. Acceptor Reaction with Lactose
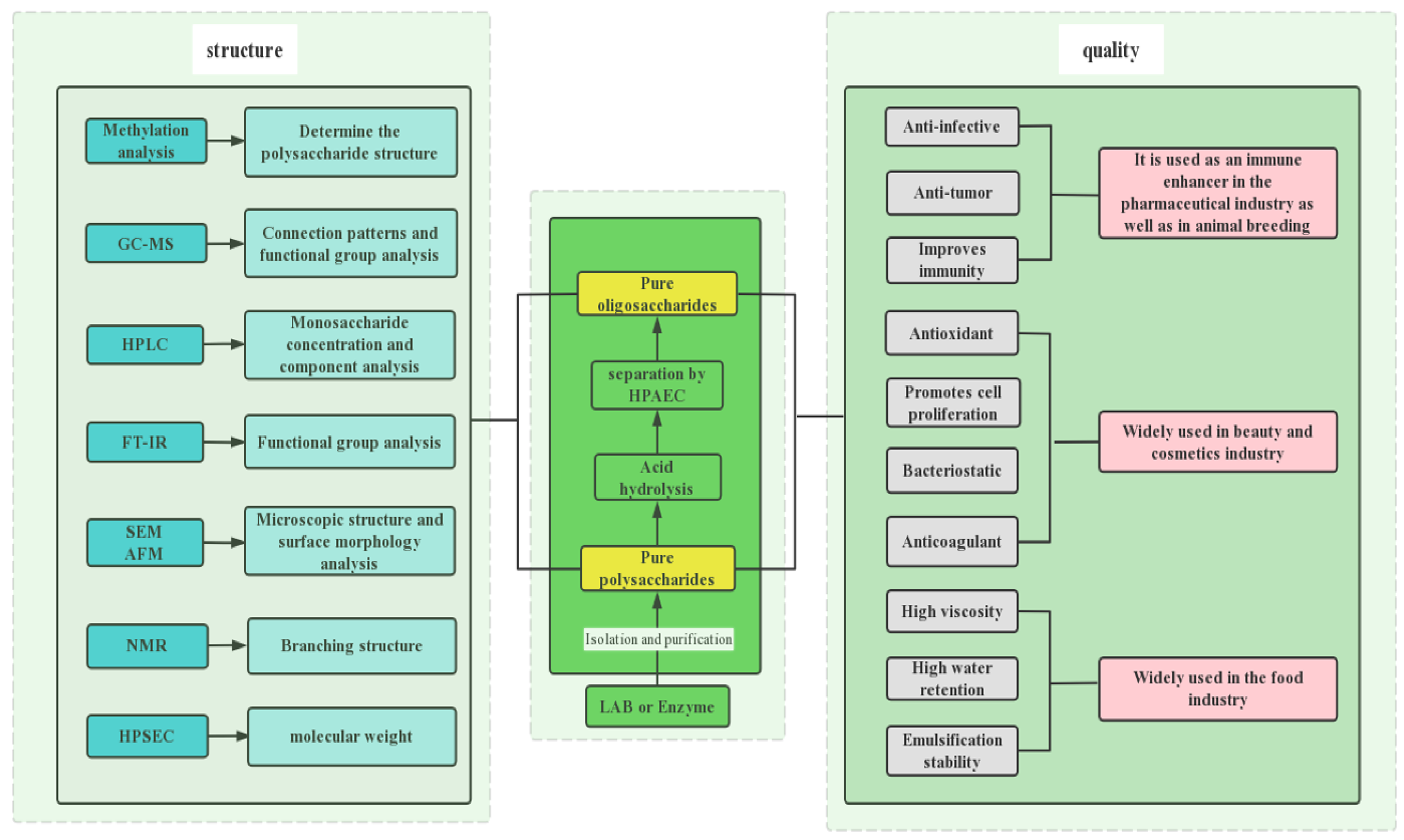
8. Analysis of EPS
8.1. Determination of Monosaccharide Composition in EPS
8.2. Determination of Functional Groups in EPS
8.3. Determination of Morphology in EPS
8.4. Determination of Structure in EPS
8.5. Thermal Analysis of EPS
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, Y.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Exopolysaccharides produced by lactic acid bacteria and Bifidobacteria: Structures, physiochemical functions and applications in the food industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Rana, S.; Upadhyay, L.S.B. Microbial exopolysaccharides: Synthesis pathways, types and their commercial applications. Int. J. Biol. Macromol. 2020, 157, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.; Pijning, T.; Dijkhuizen, L.; van Leeuwen, S.S. Mutational analysis of the role of the glucansucrase Gtf180-ΔN active site residues in product and linkage specificity with lactose as acceptor substrate. J. Agric. Food Chem. 2018, 66, 12544–12554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Pijning, T.; Tietema, M.; Dobruchowska, J.M.; Yin, H.; Gerwig, G.J.; Kralj, S.; Dijkhuizen, L. Characterization of the glucansucrase GTF180 W1065 mutant enzymes producing polysaccharides and oligosaccharides with altered linkage composition. Food Chem. 2017, 217, 81–90. [Google Scholar] [CrossRef]
- Molina, M.; Cioci, G.; Moulis, C.; Séverac, E.; Remaud-Siméon, M. Bacterial α-glucan and branching sucrases from GH70 family: Discovery, structure–function relationship studies and engineering. Microorganisms 2021, 9, 1607. [Google Scholar] [CrossRef]
- Holt, S.M.; Skory, C.; Cote, G. Enzymatic synthesis of artificial polysaccharides. ACS Sustain. Chem. Eng. 2020, 8, 11853–11871. [Google Scholar] [CrossRef]
- Chen, Z.; Ni, D.; Zhang, W.; Stressler, T.; Mu, W. Lactic acid bacteria-derived α-glucans: From enzymatic synthesis to miscellaneous applications. Biotechnol. Adv. 2021, 47, 107708. [Google Scholar] [CrossRef]
- Kim, M.; Jang, J.-K.; Park, Y.-S. Production optimization, structural analysis, and prebiotic-and anti-inflammatory effects of gluco-oligosaccharides produced by Leuconostoc lactis SBC001. Microorganisms 2021, 9, 200. [Google Scholar] [CrossRef]
- Hasselwander, O.; DiCosimo, R.; You, Z.; Cheng, Q.; Rothman, S.C.; Suwannakham, S.; Baer, Z.C.; Roesch, B.M.; Ruebling-Jass, K.D.; Lai, J.P. Development of dietary soluble fibres by enzymatic synthesis and assessment of their digestibility in vitro, animal and randomised clinical trial models. Int. J. Food Sci. Nutr. 2017, 68, 849–864. [Google Scholar] [CrossRef] [Green Version]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Kolida, S.; Naeye, T.; Einerhand, A.W.; Gibson, G.R.; Rastall, R.A. The prebiotic effect of α-1, 2 branched, low molecular weight dextran in the batch and continuous faecal fermentation system. J. Funct. Foods 2013, 5, 1938–1946. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Kolida, S.; Naeye, T.; Einerhand, A.; Brison, Y.; Remaud-Simeon, M.; Monsan, P.; Gibson, G.R.; Rastall, R.A. In vitro fermentation of linear and α-1, 2-branched dextrans by the human fecal microbiota. Appl. Environ. Microbiol. 2011, 77, 5307–5315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daba, G.M.; Elnahas, M.O.; Elkhateeb, W.A. Contributions of exopolysaccharides from lactic acid bacteria as biotechnological tools in food, pharmaceutical, and medical applications. Int. J. Biol. Macromol. 2021, 173, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Korcz, E.; Kerényi, Z.; Varga, L. Dietary fibers, prebiotics, and exopolysaccharides produced by lactic acid bacteria: Potential health benefits with special regard to cholesterol-lowering effects. Food Funct. 2018, 9, 3057–3068. [Google Scholar] [CrossRef]
- Kabli, M.; İspirli, H.; Balubaid, M.; Taylan, O.; Yılmaz, M.T.; Dertli, E. Optimization of lactose derivative hetero-oligosaccharides production using whey as the acceptor molecule by an active glucansucrase. Biocatal. Biotransform. 2022, 40, 9–16. [Google Scholar] [CrossRef]
- Song, L.; Miao, M.; Jiang, B.; Xu, T.; Cui, S.W.; Zhang, T. Leuconostoc citreum SK24.002 glucansucrase: Biochemical characterisation and de novo synthesis of α-glucan. Int. J. Biol. Macromol. 2016, 91, 123–131. [Google Scholar] [CrossRef]
- Guzman, G.Y.F.; Hurtado, G.B.; Ospina, S.A. New dextransucrase purification process of the enzyme produced by Leuconostoc mesenteroides IBUN 91.2. 98 based on binding product and dextranase hydrolysis. J. Biotechnol. 2018, 265, 8–14. [Google Scholar] [CrossRef]
- Vasileva, T.; Bivolarski, V.; Michailova, G.; Salim, A.; Rabadjiev, Y.; Ivanova, I.; Iliev, I. Glucansucrases produced by fructophilic lactic acid bacteria Lactobacillus kunkeei H3 and H25 isolated from honeybees. J. Basic Microbiol. 2017, 57, 68–77. [Google Scholar] [CrossRef]
- Meng, X.; Li, X.; Pijning, T.; Wang, X.; van Leeuwen, S.S.; Dijkhuizen, L.; Chen, G.; Liu, W. Characterization of the (engineered) branching sucrase gtfz-cd2 from Apilactobacillus kunkeei for efficient glucosylation of benzenediol compounds. Appl. Environ. Microbiol. 2022, 88, e01031-22. [Google Scholar] [CrossRef]
- Vujičić-Žagar, A.; Pijning, T.; Kralj, S.; López, C.A.; Eeuwema, W.; Dijkhuizen, L.; Dijkstra, B.W. Crystal structure of a 117 kDa glucansucrase fragment provides insight into evolution and product specificity of GH70 enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 21406–21411. [Google Scholar] [CrossRef]
- Ito, K.; Ito, S.; Shimamura, T.; Weyand, S.; Kawarasaki, Y.; Misaka, T.; Abe, K.; Kobayashi, T.; Cameron, A.D.; Iwata, S. Crystal structure of glucansucrase from the dental caries pathogen Streptococcus mutans. J. Mol. Biol. 2011, 408, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Stam, M.R.; Danchin, E.G.; Rancurel, C.; Coutinho, P.M.; Henrissat, B. Dividing the large glycoside hydrolase family 13 into subfamilies: Towards improved functional annotations of α-amylase-related proteins. Protein Eng. Des. Sel. 2006, 19, 555–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangoiti, J.; Pijning, T.; Dijkhuizen, L. Biotechnological potential of novel glycoside hydrolase family 70 enzymes synthesizing α-glucans from starch and sucrose. Biotechnol. Adv. 2018, 36, 196–207. [Google Scholar] [CrossRef]
- Meng, X.; Gangoiti, J.; Bai, Y.; Pijning, T.; Van Leeuwen, S.S.; Dijkhuizen, L. Structure–function relationships of family GH70 glucansucrase and 4, 6-α-glucanotransferase enzymes, and their evolutionary relationships with family GH13 enzymes. Cell. Mol. Life Sci. 2016, 73, 2681–2706. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.T.; Zhang, H.B.; Huang, L.J.; Hu, X.Q. Purification and characterization of extracellular dextranase from a novel producer, Hypocrea lixii F1002, and its use in oligodextran production. Process Biochem. 2011, 46, 1942–1950. [Google Scholar] [CrossRef]
- Moulis, C.; Vaca Medina, G.; Suwannarangsee, S.; Monsan, P.; Remaud-Simeon, M.; Potocki-Veronese, G. One-step synthesis of isomalto-oligosaccharide syrups and dextrans of controlled size using engineered dextransucrase. Biocatal. Biotransform. 2008, 26, 141–151. [Google Scholar] [CrossRef]
- Leemhuis, H.; Pijning, T.; Dobruchowska, J.M.; van Leeuwen, S.S.; Kralj, S.; Dijkstra, B.W.; Dijkhuizen, L. Glucansucrases: Three-dimensional structures, reactions, mechanism, α-glucan analysis and their implications in biotechnology and food applications. J. Biotechnol. 2013, 163, 250–272. [Google Scholar] [CrossRef] [Green Version]
- van Hijum, S.A.; Kralj, S.; Ozimek, L.K.; Dijkhuizen, L.; van Geel-Schutten, I.G. Structure-function relationships of glucansucrase and fructansucrase enzymes from lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 157–176. [Google Scholar] [CrossRef] [Green Version]
- André, I.; Potocki-Véronese, G.; Morel, S.; Monsan, P.; Remaud-Siméon, M. Sucrose-utilizing transglucosidases for biocatalysis. Carbohydr. Sustain. Dev. I 2010, 294, 25–48. [Google Scholar] [CrossRef]
- Korakli, M.; Vogel, R.F. Structure/function relationship of homopolysaccharide producing glycansucrases and therapeutic potential of their synthesised glycans. Appl. Microbiol. Biotechnol. 2006, 71, 790–803. [Google Scholar] [CrossRef]
- Robyt, J.F.; Yoon, S.H.; Mukerjea, R. Dextransucrase and the mechanism for dextran biosynthesis. Carbohydr. Res. 2008, 343, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Falconer, D.J.; Mukerjea, R.; Robyt, J.F. Biosynthesis of dextrans with different molecular weights by selecting the concentration of Leuconostoc mesenteroides B-512FMC dextransucrase, the sucrose concentration, and the temperature. Carbohydr. Res. 2011, 346, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Robyt, J.F.; Lee, S.Y.; Lee, J.H.; Kim, Y.M. Dextran molecular size and degree of branching as a function of sucrose concentration, pH, and temperature of reaction of Leuconostoc mesenteroides B-512FMCM dextransucrase. Carbohydr. Res. 2003, 338, 1183–1189. [Google Scholar] [CrossRef]
- Kralj, S.; van Geel-Schutten, G.H.; Dondorff, M.M.G.; Kirsanovs, S.; Van Der Maarel, M.; Dijkhuizen, L. Glucan synthesis in the genus Lactobacillus: Isolation and characterization of glucansucrase genes, enzymes and glucan products from six different strains. Microbiology 2004, 150, 3681–3690. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Cho, S.K.; Eom, H.J.; Kim, S.Y.; Kim, T.J.; Han, N.S. Optimized substrate concentrations for production of long-chain isomaltooligosaccharides using dextransucrase of Leuconostoc mesenteroides B-512F. J. Microbiol. Biotechnol. 2008, 18, 1141–1145. [Google Scholar]
- Meng, X.; Dobruchowska, J.M.; Pijning, T.; Gerwig, G.J.; Kamerling, J.P.; Dijkhuizen, L. Truncation of domain V of the multidomain glucansucrase GTF180 of Lactobacillus reuteri 180 heavily impairs its polysaccharide-synthesizing ability. Appl. Microbiol. Biotechnol. 2015, 99, 5885–5894. [Google Scholar] [CrossRef]
- Lawson, C.L.; van Montfort, R.; Strokopytov, B.; Rozeboom, H.J.; Kalk, K.H.; de Vries, G.E.; Penninga, D.; Dijkhuizen, L.; Dijkstra, B.W. Nucleotide sequence and X-ray structure of cyclodextrin glycosyltransferase from Bacillus circulans strain 251 in a maltose-dependent crystal form. J. Mol. Biol. 1994, 236, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Kralj, S.; Eeuwema, W.; Eckhardt, T.H.; Dijkhuizen, L. Role of asparagine 1134 in glucosidic bond and transglycosylation specificity of reuteransucrase from Lactobacillus reuteri 121. FEBS J. 2006, 273, 3735–3742. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, S.S.; Kralj, S.; van Geel-Schutten, I.H.; Gerwig, G.J.; Dijkhuizen, L.; Kamerling, J.P. Structural analysis of the α-D-glucan (EPS180) produced by the Lactobacillus reuteri strain 180 glucansucrase GTF180 enzyme. Carbohydr. Res. 2008, 343, 1237–1250. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, S.; Lona, L.; Franco, T. Effect of phosphate concentration on the production of dextransucrase by Leuconostoc mesenteroides NRRL B512F. Bioprocess Biosyst. Eng. 2003, 26, 57–62. [Google Scholar] [CrossRef]
- Otts, D.; Day, D.F. Dextransucrase secretion in Leuconostoc mesenteroides depends on the presence of a transmembrane proton gradient. J. Bacteriol. 1988, 170, 5006–5011. [Google Scholar] [CrossRef] [PubMed]
- Quirasco, M.; Lopez-Munguia, A.; Remaud-Simeon, M.; Monsan, P.; Farres, A. Induction and transcription studies of the dextransucrase gene in Leuconostoc mesenteroides NRRL B-512F. Appl. Environ. Microbiol. 1999, 65, 5504–5509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, A.; Katiyar, S.S. Fractionation of Leuconostoc mesenteroides NRRL B-512F dextran sucrase by polyethylene glycol: A simple and effective method purification. J. Microbiol. Methods 1994, 20, 225–231. [Google Scholar] [CrossRef]
- Purama, R.K.; Goyal, A. Identification, effective purification and functional characterization of dextransucrase from Leuconostoc mesenteroides NRRL B-640. Bioresour. Technol. 2008, 99, 3635–3642. [Google Scholar] [CrossRef]
- Kobayashi, M.; Matsuda, K. Electrophoretic analysis of the multiple forms of dextransucrase from Leuconostoc mesenteroides. J. Biochem. 1986, 100, 615–621. [Google Scholar] [CrossRef]
- Miller, A.W.; Eklund, S.H.; Robyt, J.F. Milligram to gram scale purification and characterization of dextransucrase from Leuconostoc mesenteroides NRRL B-512F. Carbohydr. Res. 1986, 147, 119–133. [Google Scholar] [CrossRef]
- Miller, A.W.; Robyt, J.F. Stabilization of dextransucrase from Leuconostoc mesenteroides NRRL B-512F by nonionic detergents, poly(ethylene glycol) and high-molecular-weight dextran. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1984, 785, 89–96. [Google Scholar] [CrossRef]
- Kitaoka, M.; Robyt, J.F. Use of a microtiter plate screening method for obtaining Leuconostoc mesenteroides mutants constitutive for glucansucrase. Enzym. Microb. Technol. 1998, 22, 527–531. [Google Scholar] [CrossRef]
- Monsan, P.; Lopez, A. On the production of dextran by free and immobilized dextransucrase. Biotechnol. Bioeng. 1981, 23, 2027–2037. [Google Scholar] [CrossRef]
- Neubauer, H.; Bauche, A.; Mollet, B. Molecular characterization and expression analysis of the dextransucrase DsrD of Leuconostoc mesenteroides Lcc4 in homologous and heterologous Lactococcus lactis cultures. Microbiology 2003, 149, 973–982. [Google Scholar] [CrossRef]
- Robyt, J.F.; Walseth, T.F. Production, purification, and properties of dextransucrase from Leuconostoc mesenteroides NRRL B-512F. Carbohydr. Res. 1979, 68, 95–111. [Google Scholar] [CrossRef]
- Miao, M.; Ma, Y.; Jiang, B.; Cui, S.W.; Jin, Z.; Zhang, T. Characterisations of Lactobacillus reuteri SK24. 003 glucansucrase: Implications for α-gluco-poly and oligosaccharides biosynthesis. Food Chem. 2017, 222, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Nigam, M.; Goyal, A.; Katiyar, S.S. High yield purification of dextransucrase from Leuconostoc mesenteroides NRRL B-512F by phase partitioning. J. Food Biochem. 2006, 30, 12–20. [Google Scholar] [CrossRef]
- Amari, M.; Arango, L.; Gabriel, V.; Robert, H.; Morel, S.; Moulis, C.; Gabriel, B.; Remaud-Siméon, M.; Fontagné-Faucher, C. Characterization of a novel dextransucrase from Weissella confusa isolated from sourdough. Appl. Microbiol. Biotechnol. 2013, 97, 5413–5422. [Google Scholar] [CrossRef]
- Wang, C.; Chen, S.; Zhang, H.B.; Li, Y.; Hu, X.Q. Characterization of the inserted mutagenesis dextransucrases from Leuconostoc mesenteroides 0326 to produce hyperbranched dextran. Int. J. Biol. Macromol. 2018, 112, 584–590. [Google Scholar] [CrossRef]
- Kim, Y.M.; Yeon, M.J.; Choi, N.S.; Chang, Y.H.; Jung, M.Y.; Song, J.J.; Kim, J.S. Purification and characterization of a novel glucansucrase from Leuconostoc lactis EG001. Microbiol. Res. 2010, 165, 384–391. [Google Scholar] [CrossRef]
- Rühmkorf, C.; Bork, C.; Mischnick, P.; Rübsam, H.; Becker, T.; Vogel, R.F. Identification of Lactobacillus curvatus TMW 1.624 dextransucrase and comparative characterization with Lactobacillus reuteri TMW 1.106 and Lactobacillus animalis TMW 1.971 dextransucrases. Food Microbiol. 2013, 34, 52–61. [Google Scholar] [CrossRef]
- Vidal, R.F.; Martínez, A.; Moulis, C.; Escalier, P.; Morel, S.; Remaud-Simeon, M.; Monsan, P. A novel dextransucrase is produced by Leuconostoc citreum strain B/110-1-2: An isolate used for the industrial production of dextran and dextran derivatives. J. Ind. Microbiol. Biotechnol. 2011, 38, 1499–1506. [Google Scholar] [CrossRef]
- Yi, A.R.; Lee, S.R.; Jang, M.U.; Park, J.M.; Eom, H.J.; Han, N.S.; Kim, T.J. Cloning of dextransucrase gene from Leuconostoc citreum HJ-P4 and its high-level expression in E. coli by low temperature induction. J. Microbiol. Biotechnol. 2009, 19, 829–835. [Google Scholar] [CrossRef]
- Ko, J.A.; Jeong, H.J.; Ryu, Y.B.; Park, S.J.; Wee, Y.J.; Kim, D.; Kim, Y.M.; Lee, W.S. Large increase in Leuconostoc citreum KM20 dextransucrase activity achieved by changing the strain/inducer combination in an E. coli expression system. J. Microbiol. Biotechnol. 2012, 22, 510–515. [Google Scholar] [CrossRef]
- Passerini, D.; Vuillemin, M.; Ufarté, L.; Morel, S.; Loux, V.; Fontagné-Faucher, C.; Monsan, P.; Remaud-Siméon, M.; Moulis, C. Inventory of the GH70 enzymes encoded by Leuconostoc citreum NRRL B-1299-identification of three novel α-transglucosylases. FEBS J. 2015, 282, 2115–2130. [Google Scholar] [CrossRef] [PubMed]
- Monchois, V.; Remaud-Simeon, M.; Monsan, P.; Willemot, R.M. Cloning and sequencing of a gene coding for an extracellular dextransucrase (DSRB) from Leuconostoc mesenteroides NRRL B-1299 synthesizing only a alpha (1-6) glucan. FEMS Microbiol. Lett. 1998, 159, 307–315. [Google Scholar] [CrossRef]
- Bozonnet, S.; Dols-Laffargue, M.; Fabre, E.; Pizzut, S.; Remaud-Simeon, M.; Monsan, P.; Willemot, R.M. Molecular characterization of DSR-E, an alpha-1,2 linkage-synthesizing dextransucrase with two catalytic domains. J. Bacteriol. 2002, 184, 5753–5761. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hu, Y.; Zhu, C.; Zhu, B.; Wang, Y. Cloning, sequencing and expression of a dextransucrase gene (dexYG) from Leuconostoc mesenteroides. Biotechnol. Lett. 2008, 30, 1441–1446. [Google Scholar] [CrossRef]
- Kang, H.K.; Kim, Y.M.; Kim, D.M. Functional, genetic, and bioinformatic characterization of dextransucrase (DSRBCB4) gene in Leuconostoc mesenteroides B-1299 CB4. J. Microbiol. Biotechnol. 2008, 18, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Fulton, D.B.; Robyt, J.F. Enzymatic synthesis of L-DOPA alpha-glycosides by reaction with sucrose catalyzed by four different glucansucrases from four strains of Leuconostoc mesenteroides. Carbohydr. Res. 2010, 345, 1730–1735. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Bruce Fulton, D.; Robyt, J.F. Enzymatic synthesis of two salicin analogues by reaction of salicyl alcohol with Bacillus macerans cyclomaltodextrin glucanyltransferase and Leuconostoc mesenteroides B-742CB dextransucrase. Carbohydr. Res. 2004, 339, 1517–1529. [Google Scholar] [CrossRef]
- Chellapandian, M.; Larios, C.; Sanchez-Gonzalez, M.; Lopez-Munguia, A. Production and properties of a dextransucrase from Leuconostoc mesenteroides IBT-PQ isolated from ‘pulque’, a traditional Aztec alcoholic beverage. J. Ind. Microbiol. Biotechnol. 1998, 21, 51–56. [Google Scholar] [CrossRef]
- Siddiqui, N.N.; Aman, A.; Qader, S.A. Mutational analysis and characterization of dextran synthesizing enzyme from wild and mutant strain of Leuconostoc mesenteroides. Carbohydr. Polym. 2013, 91, 209–216. [Google Scholar] [CrossRef]
- Yalin, Y.; Jin, L.; Jianhua, W.; Da, T.; Zigang, T. Expression and characterization of dextransucrase gene dsrX from Leuconostoc mesenteroides in Escherichia coli. J. Biotechnol. 2008, 133, 505–512. [Google Scholar] [CrossRef]
- Monchois, V.; Remaud-Simeon, M.; Russell, R.R.; Monsan, P.; Willemot, R.M. Characterization of Leuconostoc mesenteroides NRRL B-512F dextransucrase (DSRS) and identification of amino-acid residues playing a key role in enzyme activity. Appl. Microbiol. Biotechnol. 1997, 48, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Funane, K.; Ishii, T.; Matsushita, M.; Hori, K.; Mizuno, K.; Takahara, H.; Kitamura, Y.; Kobayashi, M. Water-soluble and water-insoluble glucans produced by Escherichia coli recombinant dextransucrases from Leuconostoc mesenteroides NRRL B-512F. Carbohydr. Res. 2001, 334, 19–25. [Google Scholar] [CrossRef]
- Gilmore, K.S.; Russell, R.R.; Ferretti, J.J. Analysis of the Streptococcus downei gtfS gene, which specifies a glucosyltransferase that synthesizes soluble glucans. Infect. Immun. 1990, 58, 2452–2458. [Google Scholar] [CrossRef] [Green Version]
- Hanada, N.; Fukushima, K.; Nomura, Y.; Senpuku, H.; Hayakawa, M.; Mukasa, H.; Shiroza, T.; Abiko, Y. Cloning and nucleotide sequence analysis of the Streptococcus sobrinus gtfU gene that produces a highly branched water-soluble glucan. Biochim. Biophys. Acta 2002, 1570, 75–79. [Google Scholar] [CrossRef]
- Tsumori, H.; Minami, T.; Kuramitsu, H.K. Identification of essential amino acids in the Streptococcus mutans glucosyltransferases. J. Bacteriol. 1997, 179, 3391–3396. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, A.; Nakano, Y.J.; Mukasa, H.; Kuramitsu, H.K. Identification of amino acid residues in Streptococcus mutans glucosyltransferases influencing the structure of the glucan product. J. Bacteriol. 1994, 176, 4845–4850. [Google Scholar] [CrossRef] [Green Version]
- Bounaix, M.S.; Robert, H.; Gabriel, V.; Morel, S.; Remaud-Siméon, M.; Gabriel, B.; Fontagné-Faucher, C. Characterization of dextran-producing Weissella strains isolated from sourdoughs and evidence of constitutive dextransucrase expression. FEMS Microbiol. Lett. 2010, 311, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Shi, Q.; Maina, N.H.; Juvonen, M.; Goyal, A. Weissella confusa Cab3 dextransucrase: Properties and in vitro synthesis of dextran and glucooligosaccharides. Carbohydr. Polym. 2014, 101, 554–564. [Google Scholar] [CrossRef]
- Kajala, I.; Shi, Q.; Nyyssölä, A.; Maina, N.H.; Hou, Y.; Katina, K.; Tenkanen, M.; Juvonen, R. Cloning and characterization of a Weissella confusa dextransucrase and its application in high fibre baking. PLoS ONE 2015, 10, e0116418. [Google Scholar] [CrossRef] [Green Version]
- Wangpaiboon, K.; Padungros, P.; Nakapong, S.; Charoenwongpaiboon, T.; Rejzek, M.; Field, R.A.; Pichyangkura, R. An α-1,6-and α-1,3-linked glucan produced by Leuconostoc citreum ABK-1 alternansucrase with nanoparticle and film-forming properties. Sci. Rep. 2018, 8, 8340. [Google Scholar] [CrossRef]
- Miao, M.; Ma, Y.; Jiang, B.; Huang, C.; Li, X.; Cui, S.W.; Zhang, T. Structural investigation of a neutral extracellular glucan from Lactobacillus reuteri SK24.003. Carbohydr. Polym. 2014, 106, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Argüello-Morales, M.A.; Remaud-Simeon, M.; Pizzut, S.; Sarçabal, P.; Willemot, R.; Monsan, P. Sequence analysis of the gene encoding alternansucrase, a sucrose glucosyltransferase from Leuconostoc mesenteroides NRRL B-1355. FEMS Microbiol. Lett. 2000, 182, 81–85. [Google Scholar] [CrossRef]
- Kralj, S.; Stripling, E.; Sanders, P.; van Geel-Schutten, G.H.; Dijkhuizen, L. Highly hydrolytic reuteransucrase from probiotic Lactobacillus reuteri strain ATCC 55730. Appl. Environ. Microbiol. 2005, 71, 3942–3950. [Google Scholar] [CrossRef] [Green Version]
- Miao, M.; Ma, Y.; Huang, C.; Jiang, B.; Cui, S.W.; Zhang, T. Physicochemical properties of a water soluble extracellular homopolysaccharide from Lactobacillus reuteri SK24.003. Carbohydr. Polym. 2015, 131, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Côté, G.L.; Skory, C.D. Cloning, expression, and characterization of an insoluble glucan-producing glucansucrase from Leuconostoc mesenteroides NRRL B-1118. Appl. Microbiol. Biotechnol. 2012, 93, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- Shiroza, T.; Ueda, S.; Kuramitsu, H.K. Sequence analysis of the gtfB gene from Streptococcus mutans. J. Bacteriol. 1987, 169, 4263–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, S.; Shiroza, T.; Kuramitsu, H.K. Sequence analysis of the gtfC gene from Streptococcus mutans GS-5. Gene 1988, 69, 101–109. [Google Scholar] [CrossRef]
- Gangoiti, J.; van Leeuwen, S.S.; Gerwig, G.J.; Duboux, S.; Vafiadi, C.; Pijning, T.; Dijkhuizen, L. 4,3-α-Glucanotransferase, a novel reaction specificity in glycoside hydrolase family 70 and clan GH-H. Sci. Rep. 2017, 7, 39761. [Google Scholar] [CrossRef] [Green Version]
- Dobruchowska, J.M.; Gerwig, G.J.; Kralj, S.; Grijpstra, P.; Leemhuis, H.; Dijkhuizen, L.; Kamerling, J.P. Structural characterization of linear isomalto-/malto-oligomer products synthesized by the novel GTFB 4,6-α-glucanotransferase enzyme from Lactobacillus reuteri 121. Glycobiology 2012, 22, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Leemhuis, H.; Dijkman, W.P.; Dobruchowska, J.M.; Pijning, T.; Grijpstra, P.; Kralj, S.; Kamerling, J.P.; Dijkhuizen, L. 4,6-α-Glucanotransferase activity occurs more widespread in Lactobacillus strains and constitutes a separate GH70 subfamily. Appl. Microbiol. Biotechnol. 2013, 97, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Pijning, T.; Gangoiti, J.; Te Poele, E.M.; Börner, T.; Dijkhuizen, L. Insights into Broad-Specificity Starch Modification from the Crystal Structure of Limosilactobacillus Reuteri NCC 2613 4,6-α-Glucanotransferase GtfB. J. Agric. Food Chem. 2021, 69, 13235–13245. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Oh, J.S.; Kim, D. Molecular characterization and expression analysis of the glucansucrase DSRWC from Weissella cibaria synthesizing a alpha (1→6) glucan. FEMS Microbiol. Lett. 2009, 292, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangoiti, J.; van Leeuwen, S.S.; Vafiadi, C.; Dijkhuizen, L. The Gram-negative bacterium Azotobacter chroococcum NCIMB 8003 employs a new glycoside hydrolase family 70 4,6-α-glucanotransferase enzyme (GtfD) to synthesize a reuteran like polymer from maltodextrins and starch. Biochim. Biophys. Acta 2016, 1860, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, J.; Pijning, T.; Dijkhuizen, L. The Exiguobacterium sibiricum 255-15 GtfC Enzyme Represents a Novel Glycoside Hydrolase 70 Subfamily of 4,6-α-Glucanotransferase Enzymes. Appl. Environ. Microbiol. 2016, 82, 756–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, H.; Koepsell, H.; Corman, J.; Bryant, G.; Bogard, M.; Feger, V.; Jackson, R. The effect of certain cultural factors on production of dextransucrase by Leuconostoc mesenteroides. J. Bacteriol. 1952, 64, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, R.; Zhao, F.; Pan, L.; Han, Y.; Xiao, H.; Zhou, Z. Optimization and purification of glucansucrase produced by Leuconostoc mesenteroides DRP2-19 isolated from Chinese Sauerkraut. Prep. Biochem. Biotechnol. 2018, 48, 465–473. [Google Scholar] [CrossRef]
- Molina, M.; Moulis, C.; Monties, N.; Pizzut-Serin, S.; Guieysse, D.; Morel, S.; Cioci, G.; Remaud-Simeéon, M. Deciphering an undecided enzyme: Investigations of the structural determinants involved in the linkage specificity of alternansucrase. ACS Catal. 2019, 9, 2222–2237. [Google Scholar] [CrossRef]
- Qader, S.A.U.; Aman, A.; Bano, S.; Syed, N.; Azhar, A. The effect of calcium ions and temperature on the production, activity and stability of dextransucrase from the newly isolated strain Leuconostoc mesenteroides PCSIR-4. Rom. J. Biochem. 2008, 45, 159–168. [Google Scholar] [CrossRef]
- Bai, Y.; van der Kaaij, R.M.; Leemhuis, H.; Pijning, T.; van Leeuwen, S.S.; Jin, Z.; Dijkhuizen, L. Biochemical characterization of the Lactobacillus reuteri glycoside hydrolase family 70 GTFB type of 4, 6-α-glucanotransferase enzymes that synthesize soluble dietary starch fibers. Appl. Environ. Microbiol. 2015, 81, 7223–7232. [Google Scholar] [CrossRef] [Green Version]
- Côté, G.L.; Robyt, J.F. Isolation and partial characterization of an extracellular glucansucrase from Leuconostoc mesenteroides NRRL B-1355 that synthesizes an alternating (1→6), (1→3)-α-D-glucan. Carbohydr. Res. 1982, 101, 57–74. [Google Scholar] [CrossRef]
- Du, R.; Qiao, X.; Wang, Y.; Zhao, B.; Han, Y.; Zhou, Z. Determination of glucansucrase encoding gene in Leuconostoc mesenteroides. Int. J. Biol. Macromol. 2019, 137, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Su, L.; Wang, L.; Wu, J.; Chen, S. Alpha-glucanotransferase from the glycoside hydrolase family synthesizes α (1–6)-linked products from starch: Features and synthesis pathways of the products. Trends Food Sci. Technol. 2022, 128, 160–172. [Google Scholar] [CrossRef]
- Díaz-Montes, E. Dextran: Sources, structures, and properties. Polysaccharides 2021, 2, 554–565. [Google Scholar] [CrossRef]
- Kothari, D.; Goyal, A. Structural characterization of enzymatically synthesized dextran and oligosaccharides from Leuconostoc mesenteroides NRRL B-1426 dextransucrase. Biochemistry 2013, 78, 1164–1170. [Google Scholar] [CrossRef]
- Wu, J.; Han, X.; Ye, M.; Li, Y.; Wang, X.; Zhong, Q. Exopolysaccharides synthesized by lactic acid bacteria: Biosynthesis pathway, structure-function relationship, structural modification and applicability. Crit. Rev. Food Sci. Nutr. 2022, 25, 1–22. [Google Scholar] [CrossRef]
- Guggentheim, B. Enzymatic hydrolysis and structure of water-insoluble glucan produced by glucosyltransferase from a strain of Streptococcus mutans. Helv. Odontol. Acta 1970, 5, 89–108. [Google Scholar]
- Kralj, S.; van Geel-Schutten, G.; Rahaoui, H.; Leer, R.; Faber, E.; Van Der Maarel, M.; Dijkhuizen, L. Molecular characterization of a novel glucosyltransferase from Lactobacillus reuteri strain 121 synthesizing a unique, highly branched glucan with α-(1→4) and α-(1→6) glucosidic bonds. Appl. Environ. Microbiol. 2002, 68, 4283–4291. [Google Scholar] [CrossRef] [Green Version]
- İspirli, H.; Yüzer, M.O.; Skory, C.; Colquhoun, I.J.; Sağdıç, O.; Dertli, E. Characterization of a glucansucrase from Lactobacillus reuteri E81 and production of malto-oligosaccharides. Biocatal. Biotransform. 2019, 37, 421–430. [Google Scholar] [CrossRef]
- Cui, L.; Hu, J.; Li, C.C.; Wang, C.M.; Zhang, C.Y. An electrochemical biosensor based on the enhanced quasi-reversible redox signal of prussian blue generated by self-sacrificial label of iron metal-organic framework. Biosens. Bioelectron. 2018, 122, 168–174. [Google Scholar] [CrossRef]
- İspirli, H.; Bowman, M.J.; Skory, C.D.; Dertli, E. Synthesis and characterization of Bifidogenic raffinose-derived oligosaccharides via acceptor reactions of glucansucrase E81. LWT 2021, 147, 111525. [Google Scholar] [CrossRef]
- İspirli, H.; Dertli, E. Production of lactose derivative hetero-oligosaccharides from whey by glucansucrase E81 and determination of prebiotic functions. LWT 2021, 137, 110471. [Google Scholar] [CrossRef]
- İspirli, H.; Dertli, E. Production of mannose-containing oligosaccharides by glucansucrase E81 and determination of their functional characteristics. Biocatal. Biotransform. 2020, 38, 202–209. [Google Scholar] [CrossRef]
- Gangoiti, J.; Corwin, S.F.; Lamothe, L.M.; Vafiadi, C.; Hamaker, B.R.; Dijkhuizen, L. Synthesis of novel α-glucans with potential health benefits through controlled glucose release in the human gastrointestinal tract. Crit. Rev. Food Sci. Nutr. 2020, 60, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.M.; Teresi, J.; Cote, G. Influence of alternansucrase-derived oligosaccharides and other carbohydrates on α-galactosidase and α-glucosidase activity in Bifidobacterium adolescentis. Lett. Appl. Microbiol. 2008, 46, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miller-Fosmore, C.; Côté, G. Growth of various intestinal bacteria on alternansucrase-derived oligosaccharides. Lett. Appl. Microbiol. 2005, 40, 385–390. [Google Scholar] [CrossRef]
- Sanz, M.L.; Côté, G.L.; Gibson, G.R.; Rastall, R.A. Prebiotic properties of alternansucrase maltose-acceptor oligosaccharides. J. Agric. Food Chem. 2005, 53, 5911–5916. [Google Scholar] [CrossRef]
- Bivolarski, V.; Vasileva, T.; Gabriel, V.; Iliev, I. Synthesis of glucooligosaccharides with prebiotic potential by glucansucrase URE 13–300 acceptor reactions with maltose, raffinose and lactose. Eng. Life Sci. 2018, 18, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Robyt, J.F.; Eklund, S.H. Relative, quantitative effects of acceptors in the reaction of Leuconostoc mesenteroides B-512F dextransucrase. Carbohydr. Res. 1983, 121, 279–286. [Google Scholar] [CrossRef]
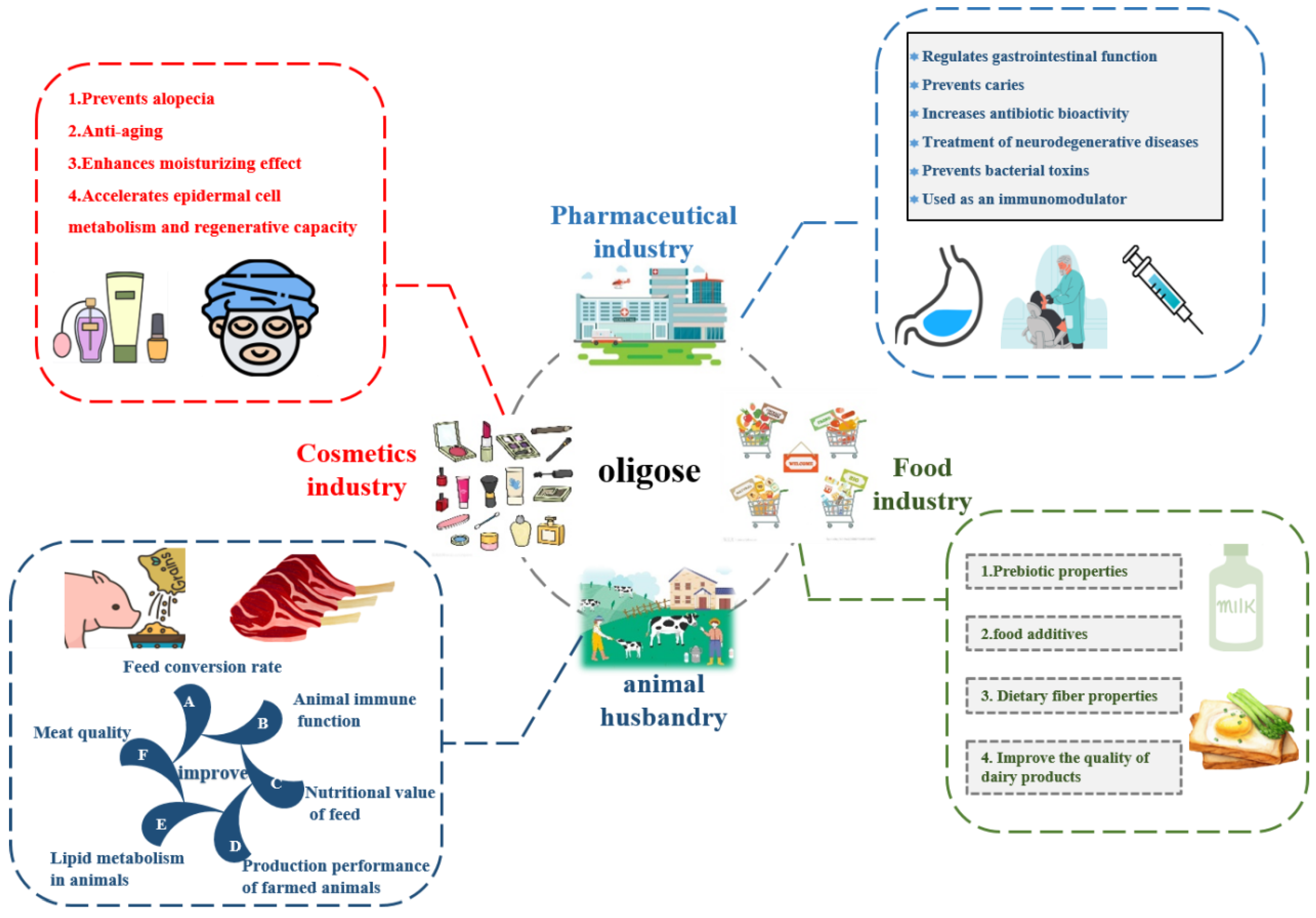
- Dahiya, D.K.; Puniya, M.; Shandilya, U.K.; Dhewa, T.; Kumar, N.; Kumar, S.; Puniya, A.K.; Shukla, P. Gut microbiota modulation and its relationship with obesity using prebiotic fibers and probiotics: A review. Front. Microbiol. 2017, 8, 563. [Google Scholar] [CrossRef]
- Chen, X.Y.; Gänzle, M.G. Lactose and lactose-derived oligosaccharides: More than prebiotics? Int. Dairy J. 2017, 67, 61–72. [Google Scholar] [CrossRef]
- CK Rajendran, S.R.; Okolie, C.L.; Udenigwe, C.C.; Mason, B. Structural features underlying prebiotic activity of conventional and potential prebiotic oligosaccharides in food and health. J. Food Biochem. 2017, 41, e12389. [Google Scholar] [CrossRef]
- Li, W.; Xia, X.; Tang, W.; Ji, J.; Rui, X.; Chen, X.; Jiang, M.; Zhou, J.; Zhang, Q.; Dong, M. Structural characterization and anticancer activity of cell-bound exopolysaccharide from Lactobacillus helveticus MB2-1. J. Agric. Food Chem. 2015, 63, 3454–3463. [Google Scholar] [CrossRef] [PubMed]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Dhanya, B.E.; Prabhu, A.; Rekha, P.D. Extraction and characterization of an exopolysaccharide from a Marine bacterium. Int. Microbiol. 2022, 25, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; Rathour, R.; Singh, R.; Thakur, I.S. Production and characterization of extracellular polymeric substances (EPS) generated by a carbofuran degrading strain Cupriavidus sp. ISTL7. Bioresour. Technol. 2019, 282, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Bejar, W.; Gabriel, V.; Amari, M.; Morel, S.; Mezghani, M.; Maguin, E.; Fontagné-Faucher, C.; Bejar, S.; Chouayekh, H. Characterization of glucansucrase and dextran from Weissella sp. TN610 with potential as safe food additives. Int. J. Biol. Macromol. 2013, 52, 125–132. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Liu, P.; Ahmed, Z.; Xiao, P.; Bai, X. Physical characterization of exopolysaccharide produced by Lactobacillus plantarum KF5 isolated from Tibet Kefir. Carbohydr. Polym. 2010, 82, 895–903. [Google Scholar] [CrossRef]
- Feng, F.; Zhou, Q.; Yang, Y.; Zhao, F.; Du, R.; Han, Y.; Xiao, H.; Zhou, Z. Characterization of highly branched dextran produced by Leuconostoc citreum B-2 from pineapple fermented product. Int. J. Biol. Macromol. 2018, 113, 45–50. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, X.; Min, W.; Yang, Z. Characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus plantarum JLK0142 isolated from fermented dairy tofu. Int. J. Biol. Macromol. 2018, 115, 985–993. [Google Scholar] [CrossRef]
- Tyagi, B.; Gupta, B.; Thakur, I.S. Biosorption of Cr (VI) from aqueous solution by extracellular polymeric substances (EPS) produced by Parapedobacter sp. ISTM3 strain isolated from Mawsmai cave, Meghalaya, India. Environ. Res. 2020, 191, 110064. [Google Scholar] [CrossRef]
- Zhu, J.; Tan, Z.; Zhang, Z.; Shi, X. Characterization on structure and bioactivities of an exopolysaccharide from Lactobacillus curvatus SJTUF 62116. Int. J. Biol. Macromol. 2022, 210, 504–517. [Google Scholar] [CrossRef]
- Sims, I.M.; Carnachan, S.M.; Bell, T.J.; Hinkley, S.F. Methylation analysis of polysaccharides: Technical advice. Carbohydr. Polym. 2018, 188, 1–7. [Google Scholar] [CrossRef]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Gerwig, G.J.; te Poele, E.M.; Dijkhuizen, L.; Kamerling, J.P. Structural analysis of rebaudioside A derivatives obtained by Lactobacillus reuteri 180 glucansucrase-catalyzed trans-alpha-glucosylation. Carbohydr. Res. 2017, 440–441, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Dertli, E.; Colquhoun, I.J.; Côté, G.L.; Le Gall, G.; Narbad, A. Structural analysis of the α-D-glucan produced by the sourdough isolate Lactobacillus brevis E25. Food Chem. 2018, 242, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Du, R.; Yu, L.; Yu, N.; Ping, W.; Song, G.; Ge, J. Characterization of exopolysaccharide produced by Levilactobacillus brevis HDE-9 and evaluation of its potential use in dairy products. Int. J. Biol. Macromol. 2022, 217, 303–311. [Google Scholar] [CrossRef]
- Zhao, D.; Cao, H.Y.; Sun, M.; Yu, L.S.; Du, R.P. Isolation, purification and structural properties analysis of exopolysaccharide from Leuconostoc pseudointestinalis HDL-3. Sci. Technol. Food Ind. 2022, 43, 115–122. [Google Scholar] [CrossRef]
- Chen, Z.; Ni, D.; Cheng, M.; Zhu, Y.; Mu, W. Comparative study of physicochemical properties of dextran and reuteran synthesised by two glucansucrases that are highly similar in amino acid sequence. Int. J. Food Sci. Technol. 2021, 56, 6674–6684. [Google Scholar] [CrossRef]
- İspirli, H.; Sagdic, O.; Yılmaz, M.T.; Dertli, E. Physicochemical characterisation of an α-glucan from Lactobacillus reuteri E81 as a potential exopolysaccharide suitable for food applications. Process Biochem. 2019, 79, 91–96. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | MASS | Organism | Genebank | Length | Reference |
|---|---|---|---|---|---|
| Gtf1624 | 183 kDa | Latilactobacillus curvatus TMW 1.624 | CCK33643.1 | 1697 aa | [57] |
| DSR-F | 170 kDa | Ln. citreum B/110-1-2 | ACY92456.2 | 1527 aa | [58] |
| LcDS | 165 kDa | Ln. citreum HJ-P4 | BAF96719.1 | 1477 aa | [59] |
| DexT | 167 kDa | Ln citreum KM20 | ACA83218.1 | 1495 aa | [60] |
| DSR-A | 145 kDa | Ln. citreum NRRL B-1299 | CDX67012.1 | 1290 aa | [61] |
| DSR-B | 168 kDa | Ln. citreum NRRL B-1299 | AAB95453.1 | 1508 aa | [62] |
| DSR-E | 313 kDa | Ln. citreum NRRL B-1299 | CDX66820.1 | 2836 aa | [63] |
| DSR-DP | - | Ln. citreum NRRL B-1299 | CDX66641.1 | 1278 aa | [63] |
| DSR-M | 144 kDa | Ln. citreum NRRL B-1299 | CDX66895.1 | 1293 aa | [61] |
| DexYG | 170 kDa | Ln. mesenteroides 0326 | ABC75033.1 | 1527 aa | [64] |
| DsrBCB4 | 168 kDa | Ln. mesenteroides B-1299CB4 | ABF85832.1 | 1505 aa | [65] |
| DsrC | 165 kDa | Ln. mesenteroides B-1355 | CAB76565.1 | 1477 aa | [66] |
| Dsrb74 | 169 kDa | Ln. mesenteroides B-742CB | AAG38021.1 | 1508 aa | [67] |
| DsrP | 161 kDa | Ln. mesenteroides IBT-PQ | AAS79426.1 | 1454 aa | [68] |
| DsrN | 169 kDa | Ln. mesenteroides KIBGE-IB-22 | AFP53921.1 | 1527 aa | [69] |
| DsrX | 169 kDa | Ln. mesenteroides L0309 | AAQ98615.2 | 1522 aa | [70] |
| DsrD | 169 kDa | Ln. mesenteroides LCC4 | AAG61158.1 | 1527 aa | [50] |
| DSR-S | 169 kDa | Ln. mesenteroides NRRL B-512F | AAD10952.1 | 1527 aa | [71] |
| DSR-T | 110 kDa | Ln. mesenteroides NRRL B-512F | BAA90527.1 | 1016 aa | [72] |
| Gtf1971 | 178 kDa | Ligilactobacillus animalis TMW 1.971 | CCK33644.1 | 1585 aa | [57] |
| Gtf106A | 199 kDa | L. reuteri TMW 1.106 | ABP88726.1 | 1782 aa | [57] |
| GTF-S | 151 kDa | Streptococcus downei MFE 28 | AAA26898.1 | 1365 aa | [73] |
| GTF-U | 176 kDa | Streptococcus sobrinus | BAA14241.1 | 1592 aa | [74] |
| GTF-B | 166 kDa | Streptococcus mutans GS 5 | AAA88588.1 | 1476 aa | [75] |
| GTF-D | 163 kDa | S. mutans GS 5 | AAA26895.1 | 1462 aa | [76] |
| DsrK39 | 158 kDa | Weissella cibaria LBAE-K39 | ADB43097.3 | 1445 aa | [77] |
| WcCab3-DSR | 154 kDa | Weissella confusa Cab3 | AKE50934.1 | 1401 aa | [78] |
| DSR-C39-2 | 155 kDa | W. confusa LBAE C39-2 | CCF30682.1 | 1412 aa | [54] |
| DSR | 156 kDa | W. confusa VTT E-90392 | AHU88292.1 | 1418 aa | [79] |
| LcALT | 229 kDa | Ln. citreum ABK-1 | AIM52834.1 | 2057 aa | [80] |
| GtfB-SK2 | - | Ln. citreum SK24.002 | - | - | [81] |
| ASR | 229 kDa | Ln. mesenteroides NRRL B-1355 | CAB65910.2 | 2057 aa | [82] |
| GtfO | 197 kDa | L. reuteri ATCC 55730 | AAY86923.1 | 1781 aa | [83] |
| Gtf-SK3 | - | L. reuteri SK24.003 | - | - | [84] |
| GtfML1 | - | L. reuteri ML1 | - | - | [34] |
| DSRI | - | Ln. mesenteroides NRRL B-1118 | - | - | [85] |
| GtfB | - | S. mutans GS5 | - | - | [86] |
| GtfC | - | S. mutans GS5 | - | - | [87] |
| GTF-Kg15 | 174 kDa | Latilactobacillus sakei KG15 | AAU08011.1 | 1595 aa | [34] |
| GTF-33 | 172 kDa | Lentilactobacillus parabuchneri 33 | AAU08006.1 | 1561 aa | [34] |
| - | 164 kDa | Leuconostoc lactis EG001 | ACT20911.1 | 1500 aa | [56] |
| GTF-Kg3 | 161 kDa | Limosilactobacillus fermentum KG3 | AAU08008.1 | 1463 aa | [34] |
| GtfB | 179 kDa | L. f ermentum NCC2970 | AOR73699.1 | 1593 aa | [88] |
| GtfB | 179 kDa | L. reuteri 121 | AAU08014.2 | 1619 aa | [89] |
| GtfML1 | 196 kDa | L. reuteri ML1 | AAU08004.1 | 1772 aa | [34] |
| GtfML4 | 180 kDa | L. reuteri ML1 | AAU08003.2 | 1620 aa | [90] |
| GtfB | 196 kDa | L. reuteri NCC2613 | ASA47879.1 | 1662 aa | [91] |
| GtfC | 163 kDa | S. mutans | BAA26114.1 | 1455 aa | [89] |
| DsrwC | 162 kDa | W. cibaria CMU | ACK38203.1 | 1472 aa | [92] |
| GtfD | 87 kDa | Azotobacter chroococcum NCIMB 8003 | AJE22990.1 | 780 aa | [93] |
| GtfC | 99 kDa | Exiguobacterium sibiricum 255-15 | ACB62096.1 | 893 aa | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Qian, Z.; Ge, J.; Du, R. Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications. Fermentation 2022, 8, 629. https://doi.org/10.3390/fermentation8110629
Yu L, Qian Z, Ge J, Du R. Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications. Fermentation. 2022; 8(11):629. https://doi.org/10.3390/fermentation8110629
Chicago/Turabian StyleYu, Liansheng, Zhigang Qian, Jingping Ge, and Renpeng Du. 2022. "Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications" Fermentation 8, no. 11: 629. https://doi.org/10.3390/fermentation8110629
APA StyleYu, L., Qian, Z., Ge, J., & Du, R. (2022). Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications. Fermentation, 8(11), 629. https://doi.org/10.3390/fermentation8110629