
Development of Probiotic-Fermented Black Mulberry (Morus nigra L.) Juice and Its Antioxidant Activity in C2C12 Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Black Mulberry Juice and Inoculum
2.3. Selection of the Fermentation Strains and Determination of their Proportions
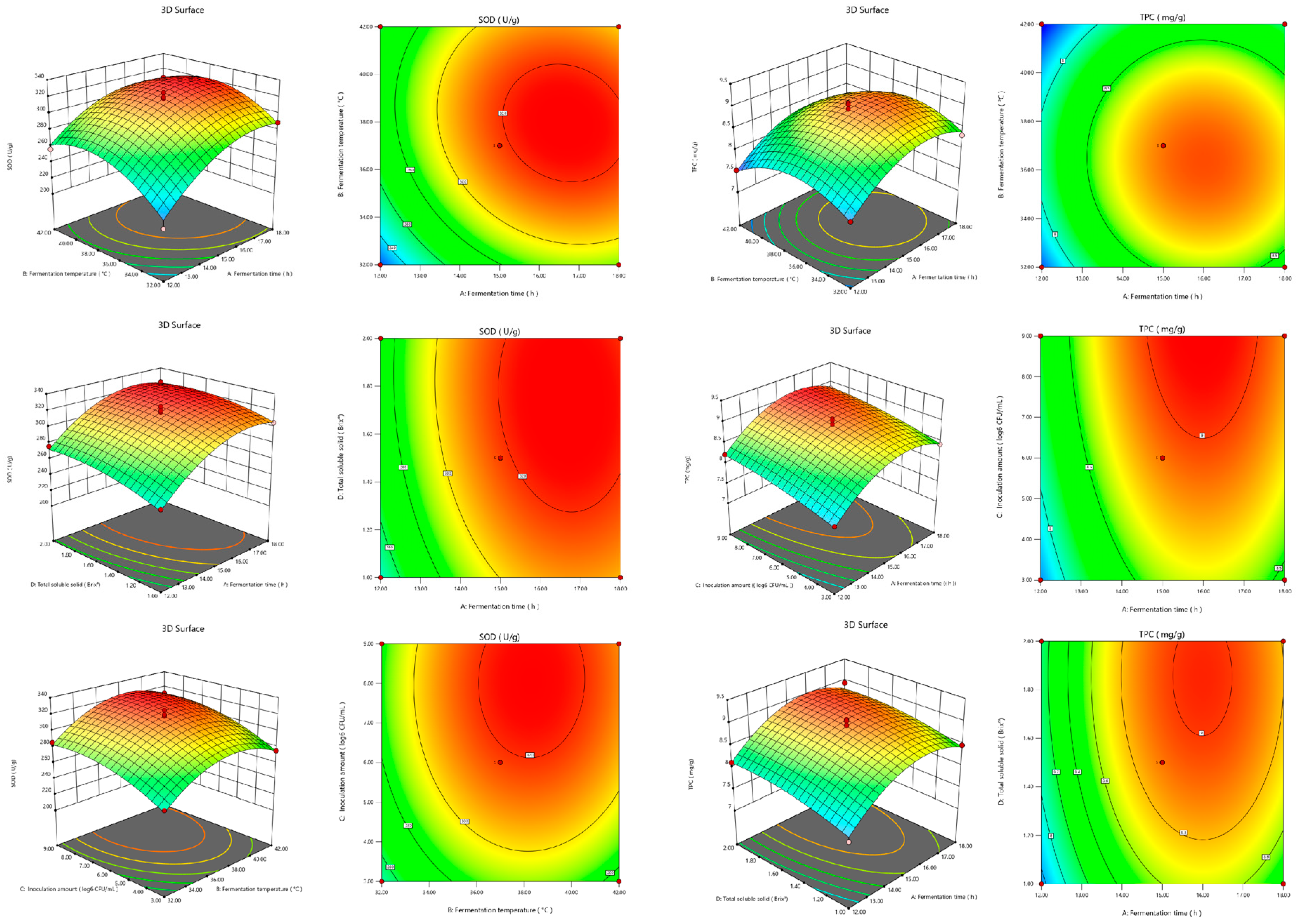
2.4. Optimization of the FBMJ Fermentation Process Using Response Surface Methodology (RSM)
2.5. Measurement of Physicochemical Indicators
2.6. Analysis of Cell Viability with the CCK-8 Assay
2.7. Cell Culture and Treatment
2.8. Evaluation of Antioxidant Activity
2.9. Statistical Analysis
3. Results
3.1. Selection of Fermentation Strains and Determination of the Proportions of Strains
3.2. Response Surface Experimental Results and Analysis of Variance
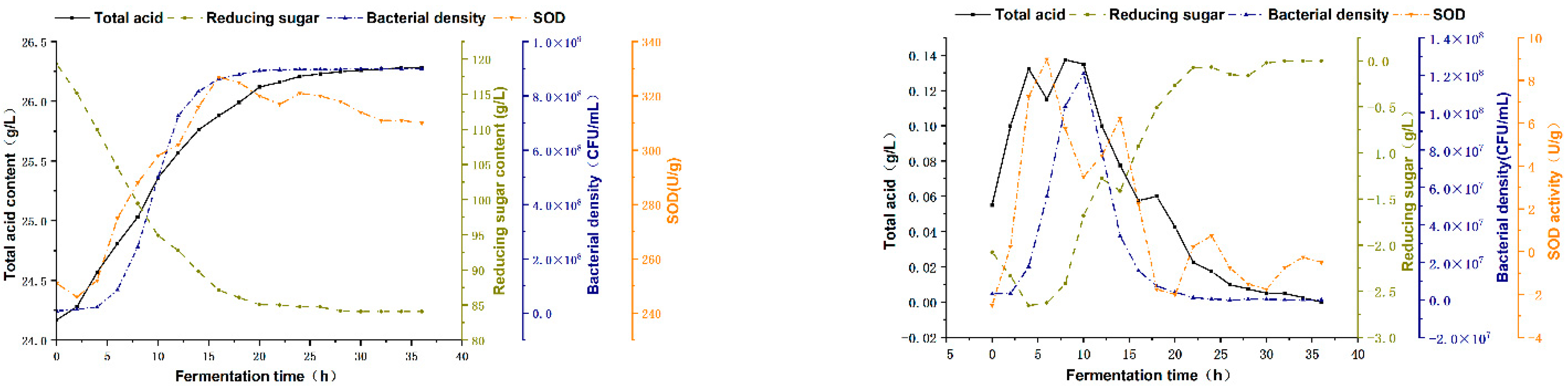
3.3. Fermentation Kinetics and Specific Rate of FBMJ
3.4. Antioxidant Activity
3.4.1. Effects of BMJ and FBMJ on H2O2-Induced Cytotoxicity in C2C12 Cells
3.4.2. Effect of BMJ and FBMJ on Intracellular ROS Levels in C2C12 Cells
3.4.3. Effects of FBMJ on SOD and GSH-Px Activities and MDA Content
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Y.; Nie, W.-J. Chemical properties in fruits of mulberry species from the Xinjiang province of China. Food Chem. 2015, 174, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Jan, B.; Parveen, R.; Zahiruddin, S.; Khan, M.U.; Mohapatra, S.; Ahmad, S. Nutritional constituents of mulberry and their potential applications in food and pharmaceuticals: A review. Saudi, J. Biol. Sci. 2021, 28, 3909–3921. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Long, X.; Liao, S.; Pang, D.; Li, Q.; Zou, Y. Effect of mulberry galacto-oligosaccharide isolated from mulberry on glucose metabolism and gut microbiota in a type 2 diabetic mice. J. Funct. Foods 2021, 87, 104836. [Google Scholar] [CrossRef]
- Huang, X.; Sun, L.; Dong, K.; Wang, G.; Luo, P.; Tang, D.; Huang, Q. Mulberry fruit powder enhanced the antioxidant capacity and gel properties of hammered minced beef: Oxidation degree, rheological, and structure. LWT 2022, 154, 112648. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Q.; Zhu, S.; Liu, B.; Liu, F.; Xu, Y. Mulberry leaf (Morus alba L.): A review of its potential influences in mechanisms of action on metabolic diseases. Pharmacol. Res. 2022, 175, 106029. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Huang, Y.; Xu, J.; He, X.; Wang, Y. Anti-neuroinflammatory and antioxidant phenols from mulberry fruit (Morus alba L.). J. Funct. Foods 2020, 68, 103914. [Google Scholar] [CrossRef]
- Bhattacharjya, D.; Sadat, A.; Dam, P.; Buccini, D.F.; Mondal, R.; Biswas, T.; Biswas, K.; Sarkar, H.; Bhuimali, A.; Kati, A.; et al. Current concepts and prospects of mulberry fruits for nutraceutical and medicinal benefits. Curr. Opin. Food Sci. 2021, 40, 121–135. [Google Scholar] [CrossRef]
- Liu, Y.; Sheng, J.; Li, J.; Zhang, P.; Tang, F.; Shan, C. Influence of lactic acid bacteria on physicochemical indexes, sensory and flavor characteristics of fermented sea buckthorn juice. Food Biosci. 2022, 46, 101519. [Google Scholar] [CrossRef]
- Ranjith, F.H.; Ariffin, S.H.; Muhialdin, B.J.; Yusof, N.L.; Mohammed, N.K.; Marzlan, A.A.; Hussin, A.S.M. Influence of natural antifungal coatings produced by Lacto-fermented antifungal substances on respiration, quality, antioxidant attributes, and shelf life of mango (Mangifera indica L.). Postharvest Biol. Technol. 2022, 189, 111904. [Google Scholar] [CrossRef]
- Tang, R.; Yu, H.; Qi, M.; Yuan, X.; Ruan, Z.; Hu, C.; Xiao, M.; Xue, Y.; Yao, Y.; Liu, Q. Biotransformation of citrus fruits phenolic profiles by mixed probiotics in vitro anaerobic fermentation. LWT 2022, 160, 113087. [Google Scholar] [CrossRef]
- Tang, S.; Cheng, Y.; Wu, T.; Hu, F.; Pan, S.; Xu, X. Effect of Lactobacillus plantarum-fermented mulberry pomace on antioxidant properties and fecal microbial community. LWT 2021, 147, 111651. [Google Scholar] [CrossRef]
- Kim, B.S.; Kim, H.; Kang, S.-S. In vitro anti-bacterial and anti-inflammatory activities of lactic acid bacteria-biotransformed mulberry (Morus alba Linnaeus) fruit extract against Salmonella Typhimurium. Food Control 2019, 106, 106758. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, L. Development of Cistanche deserticola Fermented Juice and Its Repair Effect on Ethanol-Induced WRL68 Cell Damage. Fermentation 2022, 8, 178. [Google Scholar] [CrossRef]
- Maghamian, N.; Goli, M.; Najarian, A. Ultrasound-assisted preparation of double nano-emulsions loaded with glycyrrhizic acid in the internal aqueous phase and skim milk as the external aqueous phase. LWT 2021, 141, 110850. [Google Scholar] [CrossRef]
- Zaghian, N.; Goli, M. Optimization of the production conditions of primary (W1/O) and double (W1/O/W2) nano-emulsions containing vitamin B12 in skim milk using ultrasound wave by response surface methodology. J. Food Meas. Charact. 2020, 14, 3216–3226. [Google Scholar] [CrossRef]
- Sazesh, B.; Goli, M. Quinoa as a wheat substitute to improve the textural properties and minimize the carcinogenic acrylamide content of the biscuit. J. Food Process. Preserv. 2020, 44, e14563. [Google Scholar] [CrossRef]
- Mohd Rosli, N.N.H.; Harun, N.H.; Abdul Rahman, R.; Ngadi, N.; Samsuri, S.; Amran, N.A.; Safiei, N.Z.; Ab Hamid, F.H.; Zakaria, Z.Y.; Jusoh, M. Preservation of total phenolic content (TPC) in cucumber juice concentrate using non-thermal Progressive Freeze Concentration: Quantitative design characteristics and process optimization. J. Clean. Prod. 2022, 330, 129705. [Google Scholar] [CrossRef]
- Kou, G.; Li, Z.; Wu, C.; Liu, Y.; Hu, Y.; Guo, L.; Xu, X. Citrus Tangeretin Improves Skeletal Muscle Mitochondrial Biogenesis via Activating the AMPK-PGC1-α Pathway In Vitro and In Vivo: A Possible Mechanism for Its Beneficial Effect on Physical Performance. J. Agric. Food Chem. 2018, 66, 11917–11925. [Google Scholar] [CrossRef]
- Cai, L.; Wang, W.; Tong, J.; Fang, L.; He, X.; Xue, Q.; Li, Y. Changes of bioactive substances in lactic acid bacteria and yeasts fermented kiwifruit extract during the fermentation. LWT 2022, 164, 113629. [Google Scholar] [CrossRef]
- Salman, S.; Öz, G.; Felek, R.; Haznedar, A.; Turna, T.; Özdemir, F. Effects of fermentation time on phenolic composition, antioxidant and antimicrobial activities of green, oolong, and black teas. Food Biosci. 2022, 49, 101884. [Google Scholar] [CrossRef]
- Yue, Y.; Wang, S.; Lv, X.; Wang, C.; Xu, B.; Ping, L.; Guo, J.; Li, X.; Evivie, S.E.; Liu, F.; et al. Analysis of the complete genome sequence of Lactobacillus delbrueckii ssp. bulgaricus with post-acidification capacity and its influence on yogurt in storage. J. Dairy Sci. 2022, 105, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hu, W.; Xiu, Z.; Ji, Y.; Guan, Y. Interactions between Leu. mesenteroides and L. plantarum in Chinese northeast sauerkraut. LWT 2022, 168, 113901. [Google Scholar] [CrossRef]
- Ceylan, H.G.; Atasoy, A.F. Optimization and characterization of prebiotic concentration of edible films containing Bifidobacterium animalis subsp. lactis BB-12® and its application to block type processed cheese. Int. Dairy J. 2022, 134, 105443. [Google Scholar] [CrossRef]
- Maciel da Silva, R.; Henrique Campelo, P.; Rodrigues, S. In vitro viability of L. Casei B-442 and fructooligosaccharides integrity in Amazonian sapota-do-solimões functional juice. Food Res. Int. 2022, 154, 111036. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, L.; Xing, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Bacterial diversity and fermentation quality of Moringa oleifera leaves silage prepared with lactic acid bacteria inoculants and stored at different temperatures. Bioresour. Technol. 2019, 284, 349–358. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, S.; Han, X.; Zhou, Z.; Mao, J. Combined effects of fermentation temperature and Saccharomyces cerevisiae strains on free amino acids, flavor substances, and undesirable secondary metabolites in huangjiu fermentation. Food Microbiol. 2022, 108, 104091. [Google Scholar] [CrossRef]
- Li, S.; Tao, Y.; Li, D.; Wen, G.; Zhou, J.; Manickam, S.; Han, Y.; Chai, W.S. Fermentation of blueberry juices using autochthonous lactic acid bacteria isolated from fruit environment: Fermentation characteristics and evolution of phenolic profiles. Chemosphere 2021, 276, 130090. [Google Scholar] [CrossRef]
- Ramli, I.; Posadino, A.M.; Zerizer, S.; Spissu, Y.; Barberis, A.; Djeghim, H.; Azara, E.; Bensouici, C.; Kabouche, Z.; Rebbas, K.; et al. Low concentrations of Ambrosia maritima L. phenolic extract protect endothelial cells from oxidative cell death induced by H2O2 and sera from Crohn’s disease patients. J. Ethnopharmacol. 2023, 300, 115722. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Liu, H.; Luo, X.-J.; Zhao, Z.; Zou, Z.-Y.; Li, J.; Lin, X.-J.; Liang, Y. The roles of reactive oxygen species (ROS) and autophagy in the survival and death of leukemia cells. Crit. Rev. Oncol. Hematol. 2017, 112, 21–30. [Google Scholar] [CrossRef]
- Bao, T.; Li, Y.; Xie, J.; Jia, Z.; Chen, W. Systematic evaluation of polyphenols composition and antioxidant activity of mulberry cultivars subjected to gastrointestinal digestion and gut microbiota fermentation. J. Funct. Foods 2019, 58, 338–349. [Google Scholar] [CrossRef]
- Chang, Q.; Cai, H.; Wei, L.; Lan, R. Chitosan oligosaccharides alleviate acute heat stress-induced oxidative damage by activating ERK1/2-mediated HO-1 and GSH-Px gene expression in breast muscle of broilers. Poult. Sci. 2022, 101, 101515. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | L. paracasei X1 (%) | L. casei X2 (%) | L. plantarum X3 (%) | L. delbrueckii X4 (%) | B. animalis X5 (%) | L. fermentum X6 (%) |
|---|---|---|---|---|---|---|
| 1 | 0.15 | 0.02 | 0.08 | 0.08 | 0.13 | 0.34 |
| 2 | 0.16 | 0.04 | 0.11 | 0.13 | 0.09 | 0.33 |
| 3 | 0.17 | 0.06 | 0.14 | 0.07 | 0.16 | 0.32 |
| 4 | 0.18 | 0.08 | 0.06 | 0.12 | 0.12 | 0.31 |
| 5 | 0.19 | 0.10 | 0.09 | 0.06 | 0.08 | 0.30 |
| 6 | 0.20 | 0.01 | 0.12 | 0.11 | 0.15 | 0.29 |
| 7 | 0.21 | 0.03 | 0.15 | 0.05 | 0.11 | 0.28 |
| 8 | 0.22 | 0.05 | 0.07 | 0.10 | 0.07 | 0.27 |
| 9 | 0.23 | 0.07 | 0.10 | 0.04 | 0.14 | 0.26 |
| 10 | 0.24 | 0.09 | 0.13 | 0.09 | 0.10 | 0.25 |
| Factors | Code | Levels | ||||
|---|---|---|---|---|---|---|
| −1 | 0 | 1 | ||||
| Fermentation time (h) | A | 12 | 15 | 18 | ||
| Fermentation temperature (°C) | B | 32 | 34.5 | 37 | ||
| Soluble solid content (°Brix) | C | 3 | 6 | 9 | ||
| Inoculum concentration | D | 19 | 19.5 | 20 | ||
| Run | Fermentation time (h) | Fermentation temperature (°C) | Inoculum concentration (106 CFU/mL) | Total soluble solid content (oBrix) | SOD (U/g) | TPC (mg/g) |
| 1 | 12 | 32 | 6 | 19.5 | 214.88 | 7.68 |
| 2 | 18 | 32 | 6 | 19.5 | 288.90 | 8.31 |
| 3 | 12 | 37 | 6 | 19.5 | 256.23 | 7.53 |
| 4 | 18 | 37 | 6 | 19.5 | 308.28 | 8.07 |
| 5 | 15 | 34.5 | 3 | 18 | 275.44 | 8.23 |
| 6 | 15 | 34.5 | 9 | 18 | 296.78 | 8.78 |
| 7 | 15 | 34.5 | 3 | 20 | 286.20 | 8.62 |
| 8 | 15 | 34.5 | 9 | 20 | 322.79 | 9.09 |
| 9 | 12 | 34.5 | 6 | 18 | 250.56 | 7.65 |
| 10 | 18 | 34.5 | 6 | 18 | 305.23 | 8.47 |
| 11 | 12 | 34.5 | 6 | 20 | 275.89 | 8.12 |
| 12 | 18 | 34.5 | 6 | 20 | 320.98 | 8.93 |
| 13 | 15 | 32 | 3 | 19.5 | 254.11 | 8.17 |
| 14 | 15 | 37 | 3 | 19.5 | 275.36 | 8.16 |
| 15 | 15 | 32 | 9 | 19.5 | 285.56 | 8.75 |
| 16 | 15 | 37 | 9 | 19.5 | 311.11 | 8.39 |
| 17 | 12 | 34.5 | 3 | 19.5 | 243.34 | 7.78 |
| 18 | 18 | 34.5 | 3 | 19.5 | 285.68 | 8.43 |
| 19 | 12 | 34.5 | 9 | 19.5 | 277.60 | 8.22 |
| 20 | 18 | 34.5 | 9 | 19.5 | 325.10 | 8.84 |
| 21 | 15 | 32 | 6 | 18 | 265.78 | 8.31 |
| 22 | 15 | 37 | 6 | 18 | 285.98 | 8.02 |
| 23 | 15 | 32 | 6 | 20 | 280.45 | 8.57 |
| 24 | 15 | 37 | 6 | 20 | 305.80 | 8.31 |
| 25 | 15 | 34.5 | 6 | 19.5 | 317.96 | 8.7 |
| 26 | 15 | 34.5 | 6 | 19.5 | 324.80 | 9.06 |
| 27 | 15 | 34.5 | 6 | 19.5 | 311.23 | 9.03 |
| 28 | 15 | 34.5 | 6 | 19.5 | 315.22 | 8.83 |
| 29 | 15 | 34.5 | 6 | 19.5 | 319.56 | 8.93 |
| Category | Attributes | Score |
|---|---|---|
| Appearance | Red color | 0–10 |
| Violet/black color | 0–10 | |
| Clarity | 0–10 | |
| Flavor | Black mulberry flavor | 0–10 |
| Sour flavor | 0–10 | |
| Wine bouquet | −10–0 | |
| Taste | Sweetness/Sourness (acid-sugar ratio) | 0–10 |
| Astringency | 0–10 | |
| Black mulberry taste | 0–10 | |
| Wine taste | −10–0 | |
| Overall | Acceptability | 0–10 |
| Purchase intention | 0–10 |
| SOD Activity (U/g) | TPC (mg/g) | |||
|---|---|---|---|---|
| Source | F-Value | p-Value | F-Value | p-Value |
| Model | 52.81 | <0.0001 ** | 34.45 | <0.0001 ** |
| A | 286.42 | <0.0001 ** | 126.09 | <0.0001 ** |
| B | 67.36 | <0.0001 ** | 13.06 | 0.0028 * |
| C | 113.61 | <0.0001 ** | 54.67 | <0.0001 ** |
| D | 36.28 | <0.0001 ** | 36.17 | <0.0001 ** |
| AB | 4.16 | 0.0607 | 0.18 | 0.6737 |
| AC | 0.23 | 0.6392 | 0.021 | 0.8881 |
| AD | 0.79 | 0.3887 | 2.28×10−3 | 0.9626 |
| BC | 0.16 | 0.6957 | 2.8 | 0.1166 |
| BD | 0.23 | 0.6399 | 0.021 | 0.8881 |
| CD | 2.01 | 0.1786 | 0.15 | 0.708 |
| A2 | 118.81 | <0.0001 ** | 154.61 | <0.0001 ** |
| B2 | 146.32 | <0.0001 ** | 136.98 | <0.0001 ** |
| C2 | 33.54 | <0.0001 ** | 3.99 | 0.0655 |
| D2 | 14.84 | 0.0018 ** | 9.57 | 0.0079 ** |
| Lack of Fit | 1.19 | 0.4688 Ns | 0.3 | 0.9450 Ns |
| R2 | 0.9814 | - | 0.9718 | - |
| Adj. R2 | 0.9628 | - | 0.9436 | - |
| Pred. R² | 0.9125 | - | 0.9054 | - |
| Adeq Precision | 26.4759 | - | 21.1733 | - |
| C.V. % | 1.86 | - | 1.24 | - |
| No. | Strains | SOD Activity (U/g) | TPC (mg/g) | pH | Sensory Evaluation (Score) | Time to Reach 108 CFU/mL (h) |
|---|---|---|---|---|---|---|
| 1 | Black mulberry juice | 255.19 ± 0.17n | 7.14 ± 0.01g | 3.43 ± 0.01defg | 80.00 ± 0.00ef | / |
| 2 | L. plantarum | 290.70 ± 0.31c | 7.66 ± 0.35de | 3.41 ± 0.01ijk | 88.33 ± 0.58b | 21.03 ± 0.06c |
| 3 | L. casei | 278.60 ± 0.52e | 8.16 ± 0.02b | 3.43 ± 0.01efghi | 81.00 ± 1.00e | 18.17 ± 0.29i |
| 4 | L. paracasei | 305.36 ± 0.39b | 7.89 ± 0.06c | 3.44 ± 0.01cbef | 80.33 ± 0.58e | 19.33 ± 0.29gh |
| 5 | Weissella spp. | 306.59 ± 0.52a | 7.75 ± 0.05cde | 3.38 ± 0.01l | 70.00 ± 1.00ij | 17.33 ± 0.29j |
| 6 | L. fermentum | 275.36 ± 0.45g | 8.51 ± 0.22a | 3.41 ± 0.01ijk | 80.33 ± 0.58e | 17.17 ± 0.29j |
| 7 | L. acidophilus | 280.59 ± 0.55d | 7.65 ± 0.04de | 3.38 ± 0.01l | 70.00 ± 0.00j | 18.50 ± 0.00i |
| 8 | L. reuteri | 257.63 ± 0.55l | 7.59 ± 0.85e | 3.45 ± 0.01abc | 81.00 ± 1.00e | 23.33 ± 0.29b |
| 9 | L. delbrueckii subsp. bulgaricus | 266.63 ± 0.55i | 8.15 ± 0.49b | 3.44 ± 0.01bcd | 85.00 ± 1.00c | 20.17 ± 0.29e |
| 10 | L. helveticus | 263.40 ± 0.62j | 8.09 ± 0.26b | 3.44 ± 0.00cde | 78.33 ± 0.58g | 21.00 ± 0.00d |
| 11 | L. salivarius | 267.07 ± 0.67i | 7.10 ± 0.17g | 3.43 ± 0.01defgh | 81.00 ± 1.00e | 18.17 ± 0.29i |
| 12 | L. brevis | 271.35 ± 0.56h | 7.69 ± 0.16de | 3.42 ± 0.01hijk | 72.33 ± 0.58i | 19.00 ± 0.00h |
| 13 | Leuconostoc lactis | 255.40 ± 0.63n | 7.77 ± 0.02cd | 3.41 ± 0.01ijk | 82.67 ± 0.58d | 21.83 ± 0.29c |
| 14 | P. acidilactici | 279.47 ± 0.45e | 7.16 ± 0.01fg | 3.42 ± 0.01fghij | 80.00 ± 0.00ef | 23.33 ± 0.29b |
| 15 | P. pentosaceus | 275.52 ± 0.46g | 7.15 ± 0.03fg | 3.46 ± 0.01a | 82.67 ± 0.58d | 19.50 ± 0.00fg |
| 16 | L. mesenteroides | 256.53 ± 0.46m | 8.13 ± 0.32b | 3.40 ± 0.01k | 77.67 ± 0.58g | 26.17 ± 0.29a |
| 17 | W. anomalus | 277.26 ± 0.23f | 7.67 ± 0.19de | 3.41 ± 0.01ijk | 65.68 ± 0.58l | 21.50 ± 0.00c |
| 18 | K. lactis | 231.45 ± 0.39p | 8.13 ± 0.32b | 3.41 ± 0.01jk | 67.68 ± 0.58k | 20.83 ± 0.29d |
| 19 | S. lactis | 260.20 ± 0.18k | 7.70 ± 0.26de | 3.42 ± 0.01ghij | 78.67 ± 0.58fg | 19.33 ± 0.29gh |
| 20 | L. johnsonii | 252.48 ± 0.45o | 7.32 ± 0.46f | 3.43 ± 0.01defg | 75.00 ± 0.00h | 19.83 ± 0.29ef |
| 21 | B. animalis subsp. lactis | 271.30 ± 0.34h | 7.17 ± 0.42fg | 3.46 ± 0.01ab | 90.33 ± 0.58a | 21.50 ± 0.00c |
| No. | L. paracasei X1 (%) | L. casei X2 (%) | L. plantarum X3 (%) | L. delbrueckii subsp. bulgaricus X4 (%) | B. animalis subsp. lactis X5 (%) | L. fermentum X6 (%) | SOD Activity (U/g) |
|---|---|---|---|---|---|---|---|
| 1 | 0.15 | 0.02 | 0.08 | 0.08 | 0.13 | 0.34 | 241.13 |
| 2 | 0.16 | 0.04 | 0.11 | 0.13 | 0.09 | 0.33 | 229.08 |
| 3 | 0.17 | 0.06 | 0.14 | 0.07 | 0.16 | 0.32 | 268.01 |
| 4 | 0.18 | 0.08 | 0.06 | 0.12 | 0.12 | 0.31 | 243.12 |
| 5 | 0.19 | 0.10 | 0.09 | 0.06 | 0.08 | 0.30 | 261.06 |
| 6 | 0.20 | 0.01 | 0.12 | 0.11 | 0.15 | 0.29 | 250.73 |
| 7 | 0.21 | 0.03 | 0.15 | 0.05 | 0.11 | 0.28 | 273.12 |
| 8 | 0.22 | 0.05 | 0.07 | 0.10 | 0.07 | 0.27 | 245.21 |
| 9 | 0.23 | 0.07 | 0.10 | 0.04 | 0.14 | 0.26 | 289.14 |
| 10 | 0.24 | 0.09 | 0.13 | 0.09 | 0.10 | 0.25 | 279.15 |
| Group | SOD (U/mL) | MDA (nmol/mg Prot) | GSH-Px (U/mg Prot) |
|---|---|---|---|
| Control group | 52.97 ± 0.32 | 4.57 ± 0.04 | 17.52 ± 0.68 |
| Model group | 44.65 ± 0.26 #### | 5.31 ± 0.03 #### | 9.08 ± 0.72 ### |
| BMJ(1mg/mL) | 47.79 ± 0.11 **** | 4.38 ± 0.03 **** | 22.80 ± 0.46 **** |
| FBMJ(1mg/mL) | 51.41 ± 0.34 **** | 3.63 ± 0.06 *** | 38.12 ± 0.39 **** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, M.; Aihaiti, A.; Liu, X.; Tuerhong, N.; Yang, J.; Chen, K.; Wang, L. Development of Probiotic-Fermented Black Mulberry (Morus nigra L.) Juice and Its Antioxidant Activity in C2C12 Cells. Fermentation 2022, 8, 697. https://doi.org/10.3390/fermentation8120697
Lv M, Aihaiti A, Liu X, Tuerhong N, Yang J, Chen K, Wang L. Development of Probiotic-Fermented Black Mulberry (Morus nigra L.) Juice and Its Antioxidant Activity in C2C12 Cells. Fermentation. 2022; 8(12):697. https://doi.org/10.3390/fermentation8120697
Chicago/Turabian StyleLv, Mingshan, Aihemaitijiang Aihaiti, Xiaolu Liu, Nuersiman Tuerhong, Jiangyong Yang, Keping Chen, and Liang Wang. 2022. "Development of Probiotic-Fermented Black Mulberry (Morus nigra L.) Juice and Its Antioxidant Activity in C2C12 Cells" Fermentation 8, no. 12: 697. https://doi.org/10.3390/fermentation8120697
APA StyleLv, M., Aihaiti, A., Liu, X., Tuerhong, N., Yang, J., Chen, K., & Wang, L. (2022). Development of Probiotic-Fermented Black Mulberry (Morus nigra L.) Juice and Its Antioxidant Activity in C2C12 Cells. Fermentation, 8(12), 697. https://doi.org/10.3390/fermentation8120697