
Saudi Traditional Fermented Goat Milk Protects against Experimental Non-Alcoholic Fatty Liver Disease by Hypoglycaemic and Antioxidant Potentials

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diets and Drugs
2.3. Experimental Design
2.4. Collection and Biochemical Analyses of Urine
2.5. Oral Glucose Tolerance Test (OGTT) and Intraperitoneal Insulin Tolerance Test (IPITT)
2.6. Preparation and Biochemical Analyses of the Blood Serum
2.7. Tissue Collection and Processing
2.8. Biochemical Analyses of the Liver
2.9. Histopathological Evaluation
2.10. Statistical Analysis
3. Results
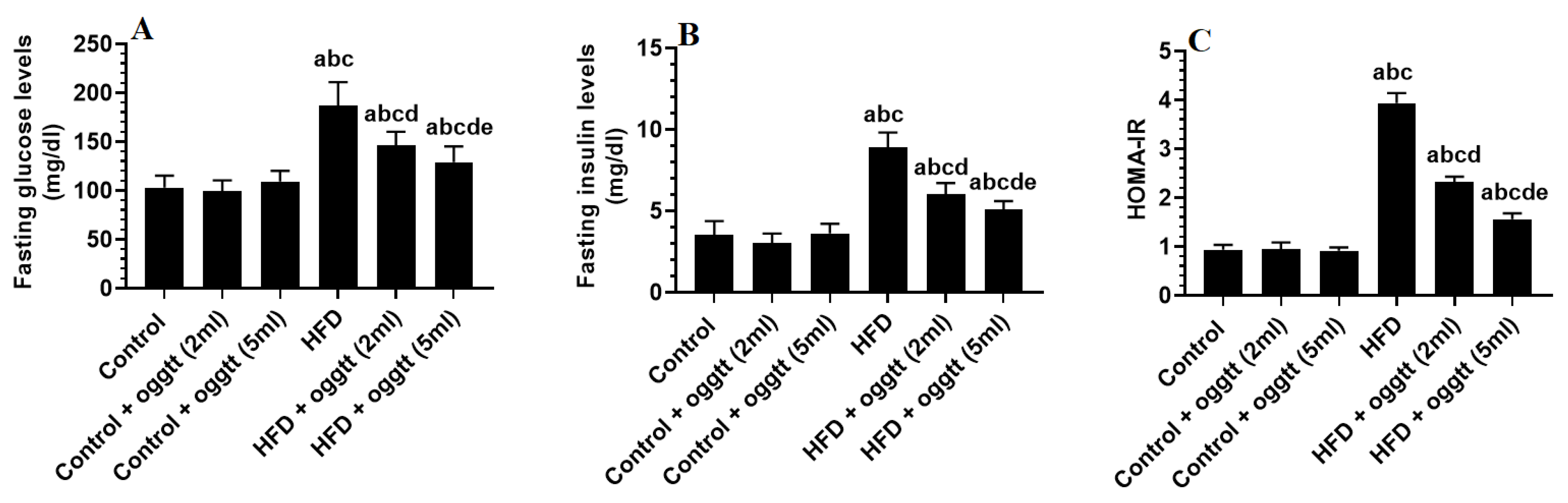
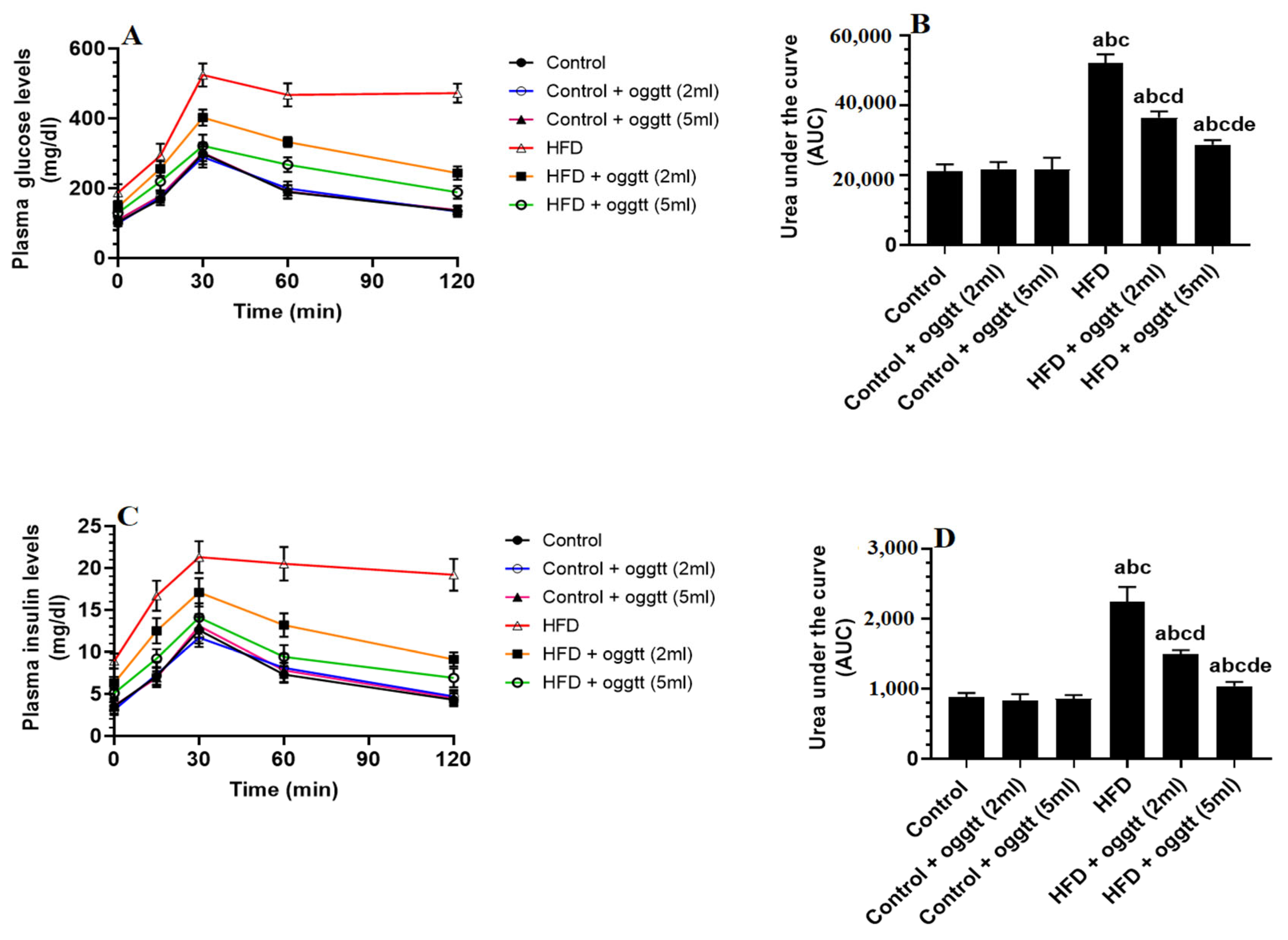
3.1. Oggtt Lowers Glucose and Insulin Levels and Improves HOMA-IR in HFD-Fed Rats
3.2. Oggtt Reduces Body Weights and Has an Anti-Hyperlipidemic Effect in HFd-Fed Rats
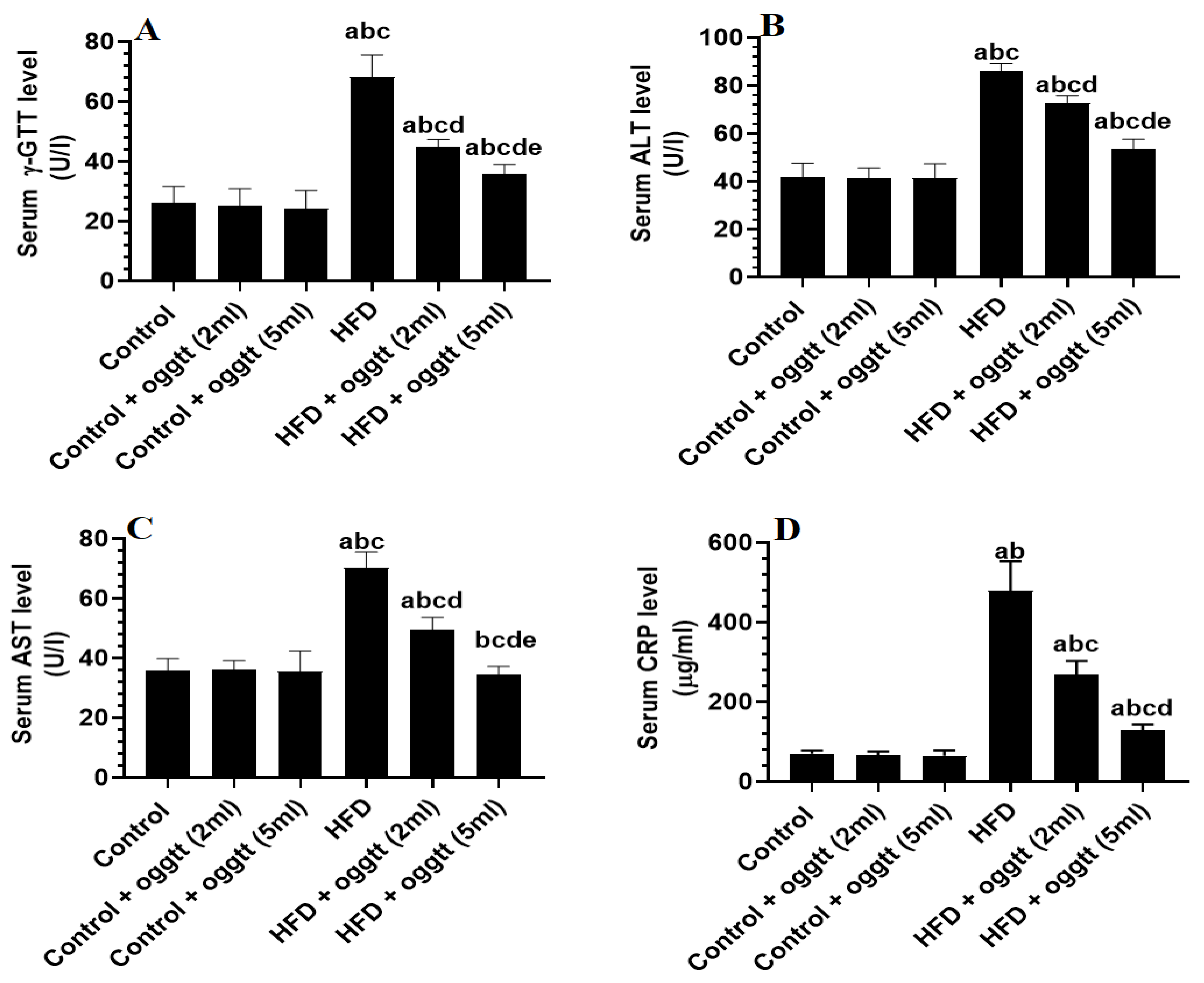
3.3. Oggtt Improves Liver Function in HFD-Fed Rats
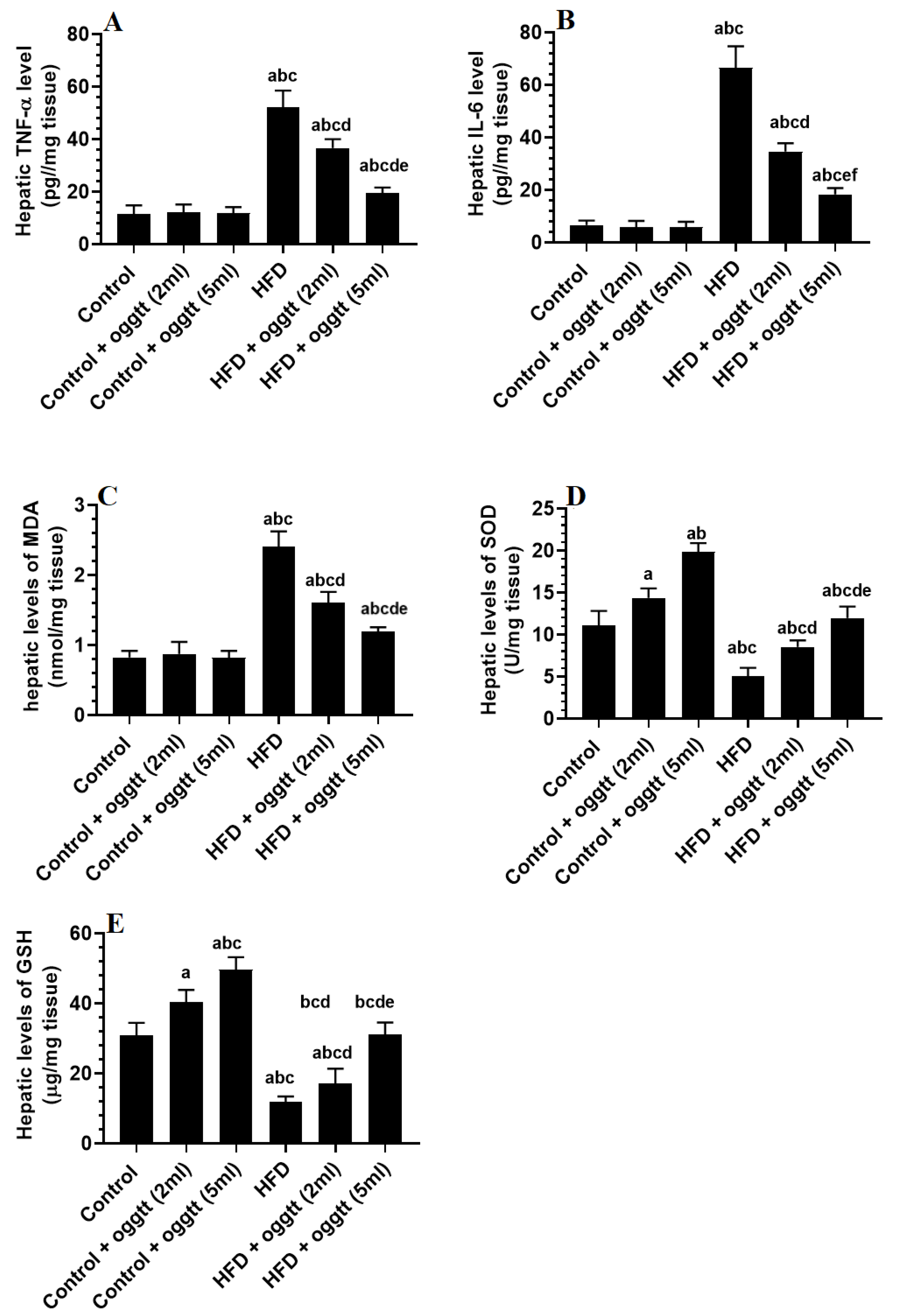
3.4. Oggtt Attenautes Hepatic Oxidaitve Stress and Inflmmation in HFD-Fed Rats
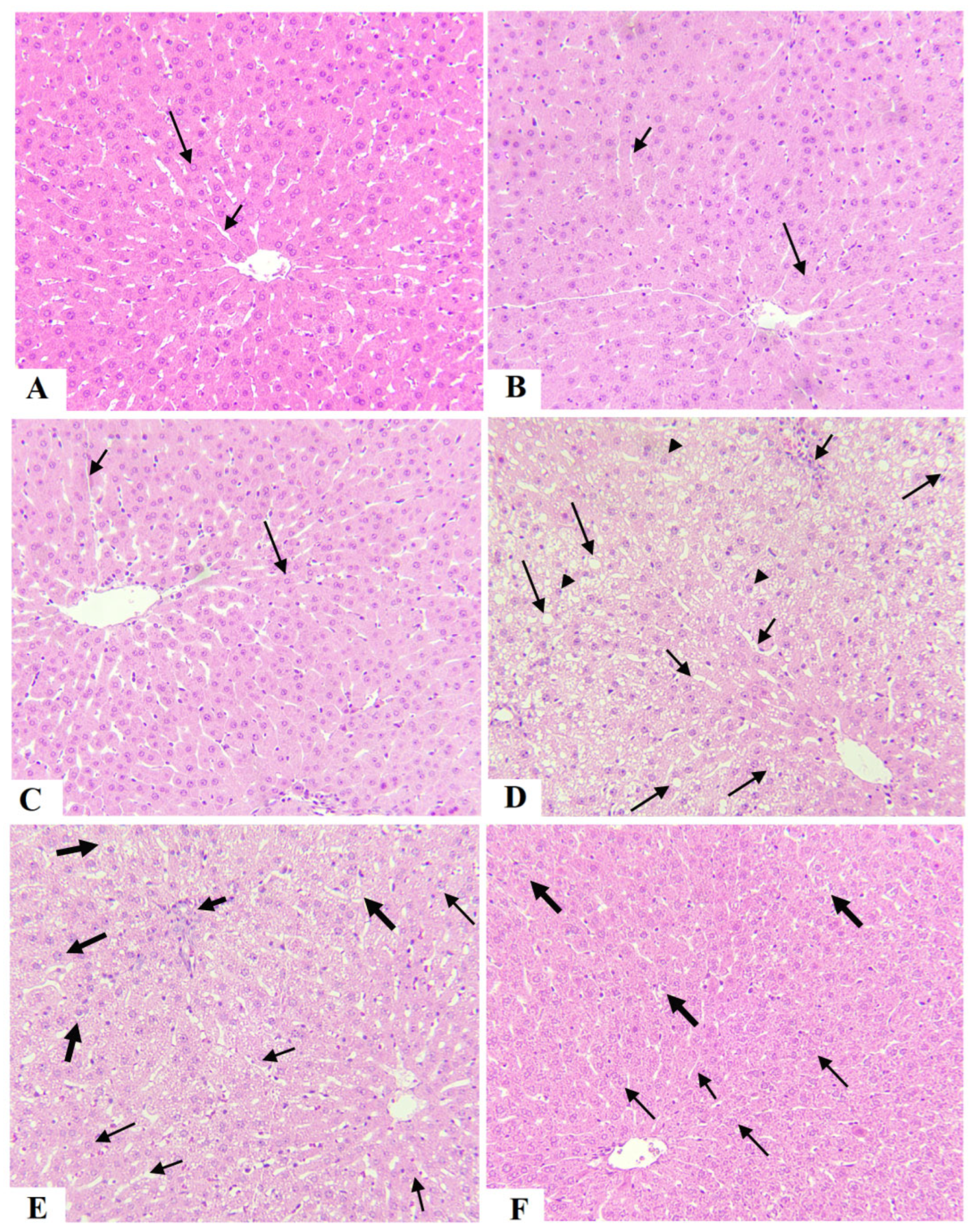
3.5. Oggtt Prevents Hepatic Steatosis in HFD-Fed Rats
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obaroakpo, J.U.; Nan, W.; Hao, L.; Liu, L.; Zhang, S.; Lu, J.; Pang, X.; Lv, J. The hyperglycemic regulatory effect of sprouted quinoa yoghurt in high-fat-diet and streptozotocin-induced type 2 diabetic mice via glucose and lipid homeostasis. Food Funct. 2020, 11, 8354–8368. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Althumiri, N.A.; Basyouni, M.H.; AlMousa, N.; AlJuwaysim, M.F.; Almubark, R.A.; BinDhim, N.F.; Alkhamaali, Z.; Alqahtani, S.A. Obesity in Saudi Arabia in 2020: Prevalence, Distribution, and Its Current Association with Various Health Conditions. Healthcare 2021, 9, 311. [Google Scholar] [CrossRef] [PubMed]
- Luci, C.; Bourinet, M.; Leclère, P.S.; Anty, R.; Gual, P. Chronic Inflammation in Non-Alcoholic Steatohepatitis: Molecular Mechanisms and Therapeutic Strategies. Front. Endocrinol. 2020, 11, 597648. [Google Scholar] [CrossRef]
- Masarone, M.; Rosato, V.; Dallio, M.; Gravina, A.G.; Aglitti, A.; Loguercio, C.; Federico, A.; Persico, M. Role of Oxidative Stress in Pathophysiology of Nonalcoholic Fatty Liver Disease. Oxidative Med. Cell. Longev. 2018, 2018, 9547613. [Google Scholar] [CrossRef]
- Yu, J.; Marsh, S.; Hu, J.; Feng, W.; Wu, C. The Pathogenesis of Nonalcoholic Fatty Liver Disease: Interplay between Diet, Gut Microbiota, and Genetic Background. Gastroenterol. Res. Pract. 2016, 2016, 2862173. [Google Scholar] [CrossRef] [Green Version]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metab. Clin. Exp. 2016, 65, 1038–1048. [Google Scholar]
- Al-Abdulkarim, B.O.; Osman, M.S.; El-Nadeef, M.A. Determination of chemical composition, and storage on dried fermented goat milk product (Oggtt). J. Saudi Soc. Agric. Sci. 2013, 12, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Dragomir, K.; Andreja, K. The Effect of Fatty Acids in Goat Milk on Health, Milk Production—An Up-to-Date Overview of Animal Nutrition, Management and Health 2012. Available online: https://www.intechopen.com/chapters/39464 (accessed on 26 September 2012).
- Hodgkinson, A.J.; Wallace, O.; Boggs, I.; Broadhurst, M.; Prosser, C.G. Gastric digestion of cow and goat milk: Impact of infant and young child in vitro digestion conditions. Food Chem. 2018, 245, 275–281. [Google Scholar] [CrossRef]
- Delgadillo-Puga, C.; Cuchillo-Hilario, M.; León-Ortiz, L.; Ramírez-Rodríguez, A.; Cabiddu, A.; Navarro-Ocaña, A.; Morales-Romero, A.M.; Medina-Campos, O.N.; Pedraza-Chaverri, J. Goats’ Feeding Supplementation with Acacia farnesiana Pods and Their Relationship with Milk Composition: Fatty Acids, Polyphenols, and Antioxidant Activity. Animals 2019, 9, 515. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo-Puga, C.; Noriega, L.G.; Morales-Romero, A.M.; Nieto-Camacho, A.; Granados-Portillo, O.; Rodríguez-López, L.A.; Alemán, G.; Furuzawa-Carballeda, J.; Tovar, A.R.; Cisneros-Zevallos, L.; et al. Goat’s Milk Intake Prevents Obesity, Hepatic Steatosis and Insulin Resistance in Mice Fed A High-Fat Diet by Reducing Inflammatory Markers and Increasing Energy Expenditure and Mitochondrial Content in Skeletal Muscle. Int. J. Mol. Sci. 2020, 21, 5530. [Google Scholar] [CrossRef]
- Ham, J.S.; Shin, J.H.; Jang, A.; Jeong, S.G.; Parkl, K.W.; Kim, H.U.; Kang, S.Y.; Hwang, H.J.; Lee, W.K. Effect of Goat Milk Yogurt Supplemented with Citrus Concentrate on Blood Glucose Hangug chugsan sigpum haghoeji and Serum Lipids in Diabetic Rats. Korean J. Food Sci. Anim. Resour. 2008, 28, 445–450. [Google Scholar] [CrossRef]
- Miglani, S.; Patyar, R.R.; Patyar, S.; Reshi, M.R. Effect of goat milk on hepatotoxicity induced by antitubercular drugs in rats. J. Food Drug Anal. 2016, 24, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, Z.; Huo, D.; Shao, Y. Consumption of goats’ milk protects mice from carbon tetrachloride-induced acute hepatic injury and improves the associated gut microbiota imbalance. Front. Immunol. 2018, 9, 1034. [Google Scholar] [CrossRef]
- Fernandez, M.A.; Marette, A. Novel perspectives on fermented milks and cardiometabolic health with a focus on type 2 diabetes. Nutr. Rev. 2018, 76, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Alsayadi, M.M.; Almowallad, S.A.; Yahya, M.A.; Alsanea, E.; Alfardi, T.; Alhaidari, S. Nutritional, Health-Promoting Properties and Antioxidant Activity of Yemeni Fermented Milk (Laban) and A Laban-Pulicaria Jaubertii Mixture. Turk. J. Agric. Food Sci. Technol. 2020, 8, 2049–2058. [Google Scholar] [CrossRef]
- Korish, A.A.; Arafah, M.M. Camel milk ameliorates steatohepatitis, insulin resistance and lipid peroxidation in experimental non-alcoholic fatty liver disease. BMC Complement. Altern. Med. 2013, 13, 264. [Google Scholar] [CrossRef] [Green Version]
- Fallah, Z.; Feizi, A.; Hashemipour, M.; Kelishadi, R. Positive Effect of Fermented Camel Milk on Liver Enzymes of Adolescents with Metabolic Syndrome: A Double Blind, Randomized, Cross-over Trial. Mater. Socio-Med. 2018, 30, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Tiss, M.; Souiy, Z.; Abdeljelil, N.; Njima, M.; Achour, L.; Hamden, K. Fermented soy milk prepared using kefir grains prevents and ameliorates obesity, type 2 diabetes, hyperlipidemia and Liver-Kidney toxicities in HFFD-rats. J. Funct. Foods 2020, 67, 103869. [Google Scholar] [CrossRef]
- Chen, H.-L.; Tung, Y.-T.; Tsai, C.-L.; Lai, C.-W.; Lai, Z.-L.; Tsai, H.-C.; Lin, Y.-L.; Wang, C.-H.; Chen, C.-M. Kefir improves fatty liver syndrome by inhibiting the lipogenesis pathway in leptin-deficient ob/ob knockout mice. Int. J. Obes. 2014, 38, 1172–1179. [Google Scholar] [CrossRef]
- Punaro, G.R.; Maciel, F.R.; Rodrigues, A.M.; Rogero, M.M.; Bogsan, C.S.B.; Oliveira, M.N.; Ihara, S.S.; Araujo, S.R.R.; Sanches, T.R.C.; Andrade, L.C.; et al. Kefir administration reduced progression of renal injury in STZ-diabetic rats by lowering oxidative stress. Nitric Oxide. Biol. Chem. 2014, 37, 53–60. [Google Scholar]
- Alsayadi, M.M.; Al jawfi, Y.; Belarbi, M.; Sabri, F.Z. Antioxidant potency of water kefir. JMBFS 2013, 2, 2444–2447. [Google Scholar]
- Bashiti, T.A.; Zabut, B.M. Effect of probiotic fermented milk (Kefir) on some blood biochemical parameters among newly diagnosed type 2 diabetic adult males in Gaza governorate. Curr. Res. Nutr. Food Sci. 2019, 7, 568–575. [Google Scholar] [CrossRef]
- de Moura EDias, M.; Dos Reis, S.A.; da Conceição, L.L.; Sediyama, C.; Pereira, S.S.; de Oliveira, L.L.; Gouveia Peluzio, M.D.C.; Martinez, J.A.; Milagro, F.I. Diet-induced obesity in animal models: Points to consider and influence on metabolic markers. Diabetol. Metab. Syndr. 2021, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Yahya, M.A.; Alhaj, O.A.; Al-Khalifah, A.S. Antihypertensive effect of fermented skim camel (Camelus dromedarius) milk on spontaneously hypertensive rats. Nutr. Hosp. 2017, 34, 416–421. [Google Scholar] [CrossRef]
- Nagy, C.; Einwallner, E. Study of In Vivo Glucose Metabolism in High-fat Diet-fed Mice Using Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT). J. Vis. Exp. 2018, 131, 56672. [Google Scholar] [CrossRef]
- Yoon, H.; Jeon, D.J.; Park, C.E.; You, H.S.; Moon, A.E.M. Relationship between homeostasis model assessment of insulin resistance and beta cell function and serum 25-hydroxyvitamin D in non-diabetic Korean adults. J. Clin. Biochem. Nutr. 2016, 59, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Alsayadi, M.; Al Jawfi, Y.; Belarbi, M.; Soualem-Mami, Z.; Merzouk, H.; Sari, D.C.; Sabri, F.; Ghalim, M. Evaluation of Anti-Hyperglycemic and Anti-Hyperlipidemic Activities of Water Kefir as Probiotic on Streptozotocin-Induced Diabetic Wistar Rats. J. Diabetes Mellit. 2014, 4, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Fan, J.; Ding, X.; Qiao, L.; Wang, G. Characterization of high-fat, diet-induced, non-alcoholic steatohepatitis with fibrosis in rats. Dig. Dis. Sci. 2010, 55, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; You, G.-P.; Cui, F.; Chen, L.-F.; Yang, X.-J.; Chen, L.-G.; Lu, H.-D.; Zhang, W.-Q. Effects of a low-fat diet on the hepatic expression of adiponectin and its receptors in rats with NAFLD. Ann. Hepatol. 2015, 14, 108–117. [Google Scholar] [CrossRef]
- Stott, N.; Marino, J. High Fat Rodent Models of Type 2 Diabetes: From Rodent to Human. Nutrients 2020, 21, 3650. [Google Scholar] [CrossRef]
- Song, L.; Qu, D.; Zhang, Q.; Jiang, J.; Zhou, H.; Jiang, R.; Li, Y.; Zhang, Y.; Yan, H. Phytosterol esters attenuate hepatic steatosis in rats with non-alcoholic fatty liver disease rats fed a high-fat diet. Sci. Rep. 2017, 7, srep41604. [Google Scholar] [CrossRef]
- Jensen, V.S.; Hvid, H.; Damgaard, J.; Nygaard, H.; Ingvorsen, C.; Wulff, E.M.; Lykkesfeldt, J.; Fledelius, C. Dietary fat stimulates development of NAFLD more potently than dietary fructose in Sprague–Dawley rats. Diabetol. Metab. Syndr. 2018, 10, 4. [Google Scholar] [CrossRef]
- Carmiel-Haggai, M.; Cederbaum, A.I.; Nieto, N. A high-fat diet leads to the progression of non-alcoholic fatty liver disease in obese rats. FASEB J. 2005, 19, 136–138. [Google Scholar] [CrossRef]
- Jaime, T.J.; Balich, L.L.; Acevedo, G.B.; de la Maza Cave, M.P.; Birn, S.H.; Parada, S.H.; Silva, J.R.; Barnett, D.B. Effect of calorie restriction on energy expenditure in overweight and obese adult women. Nutr. Hosp. 2015, 31, 2428–2436. [Google Scholar] [CrossRef]
- Panahi, S.; Doyon, C.Y.; Després, J.P.; Pérusse, L.; Vohl, M.C.; Drapeau, V.; Tremblay, A. Yogurt consumption, body composition, and metabolic health in the Quebec Family Study. Eur. J. Nutr. 2018, 57, 1591–1603. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G.; Schwedhelm, C.; Kalle-Uhlmann, T.; Missbach, B.; Knüppel, S.; Boeing, H. Consumption of dairy products in relation to changes in anthropometric variables in adult populations: Asystematic review and meta-analysis of cohort studies. PLoS ONE 2016, 11, e0157461. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Hao, T.; Rimm, E.B.; Willett, W.C.; Hu, F.B. Changes in diet and lifestyle and long-term weight gain in women and men. N. Engl. J. Med. 2011, 364, 2392–2404. [Google Scholar] [CrossRef] [Green Version]
- Rautiainen, S.; Wang, L.; Lee, I.M.; Manson, J.E.; Buring, J.E.; Sesso, H.D. Dairy consumption in association with weight change and risk of becoming overweight or obese in middle-aged and older women: A prospective cohort study. Am. J. Clin. Nutr. 2016, 103, 979–988. [Google Scholar] [CrossRef] [Green Version]
- Vincent, H.K.; Bourguignon, C.M.; Weltman, A.L.; Vincent, K.R.; Barrett, E.; Innes, K.E.; Taylor, A.G. Effects of antioxidant supplementation on insulin sensitivity, endothelial adhesion molecules, and oxidative stress in normal-weight and overweight young adults. Metabolism 2009, 58, 254–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoehn, K.L.; Salmon, A.B.; Hohnen-Behrens, C.; Turner, N.; Hoy, A.J.; Maghzal, G.J.; Stocker, R.; Van Remmen, H.; Kraegen, E.W.; Cooney, G.J.; et al. Insulin resistance is a cellular antioxidant defense mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 17787–17792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadrzadeh-Yeganeh, H.; Elmadfa, I.; Djazayery, A.; Jalali, M.; Heshmat, R.; Chamary, M. The effects of probiotic and conventional yoghurt on lipid profile in women. Br. J. Nutr. 2010, 103, 1778–1783. [Google Scholar] [CrossRef]
- Ivey, K.; Hodgson, J.; Kerr, D.; Thompson, P.; Stojceski, B.; Prince, R. The effect of yoghurt and its probiotics on blood pressure and serum lipid profile; a randomised controlled trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivey, K.; Hodgson, J.M.; Kerr, D.; Lewis, J.R.; Thompson, P.L.; Prince, R.L. The effects of probiotic bacteria on glycaemic control in overweight men and women: A randomised controlled trial. Eur. J. Clin. Nutr. 2014, 68, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Aune, D.; Norat, T.; Romundstad, P.R.; Vatten, L.J. Dairy products and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of cohort studies. Am. J. Clin. Nutr. 2013, 98, 1066–1083. [Google Scholar] [CrossRef] [Green Version]
- Gijsbers, L.; Ding, E.L.; Malik, V.S.; De Goede, J.; Geleijnse, J.M.; Soedamah-Muthu, S.S. consumption of dairy foods and diabetes incidence: A dose-response meta-analysis of observational studies. Am. J. Clin. Nutr. 2016, 103, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Bence, K.K.; Birnbaum, M.J. Metabolic drivers of non-alcoholic fatty liver disease. Mol. Metab. 2021, 50, 101143. [Google Scholar] [CrossRef]
- Sekiya, M.; Hiraishi, A.; Touyama, M.; Sakamoto, K. Oxidative stress induced lipid accumulation via SREBP1c activation in HepG2 cells. Biochem. Biophys. Res. Commun. 2008, 375, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Liss, K.H.; Finck, B.N. PPARs and nonalcoholic fatty liver disease. Biochimie 2017, 136, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Kim, H.; Jeong, D.; Kang, I.-B.; Chon, J.-W.; Kim, H.-S.; Song, K.-Y.; Seo, K.-H. Kefir alleviates obesity and hepatic steatosis in high-fat diet-fed mice by modulation of gut microbiota and mycobiota: Targeted and untargeted community analysis with correlation of biomarkers. J. Nutr. Biochem. 2017, 44, 35–43. [Google Scholar] [CrossRef]
- Mohamed, B.; Idam, N. Effect of camel milk on plasma lipid profile of hypercholesteremic rats. OJVRTM 2011, 15, 314–317. [Google Scholar]
- Ziamajidi, N.; Khaghani, S.; Hassanzadeh, G.; Vardasbi, S.; Ahmadian, S.; Nowrouzi, A.; Ghaffari, S.M.; Abdirad, A. Amelioration by chicory seed extract of diabetes- and oleic acid-induced non-alcoholic fatty liver disease (NAFLD)/non-alcoholic steatohepatitis (NASH) via modulation of PPARα and SREBP-1. Food Chem. Toxicol. 2013, 58, 198–209. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Revelo, X.; Malh, H. Pathogenesis of Nonalcoholic Steatohepatitis: An Overview. Hepatol. Commun. 2020, 4, 478–492. [Google Scholar] [CrossRef]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell Inf. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Control + Oggtt (2 mL) | Control + Oggtt (5 mL) | HFD | HFD + Oggtt (2 mL) | HFD + Oggtt (5 mL) | ||
|---|---|---|---|---|---|---|---|
| Final body weights (g) | 422 ± 42.9 | 389 ± 31.9 a | 351 ± 22.3 ab | 568 ± 41.3 abc | 487 ± 36.7 abcd | 422 ± 31.9 abcde | |
| Weekly food intake (g) | 234 ± 24.5 | 219 ± 22.7 | 229 ± 26.5 | 322 ± 29 abc | 339 ± 37.1 abc | 328 ± 32.2 abc | |
| Liver weight | 13.8 ± 1.5 | 12.9 ± 1.3 | 13.7 ± 1.1 | 18.5 ± 1.6 abc | 15.4 ± 1.5 abcd | 12.8 ± 1.1 cde | |
| Serum | TGs (mg/dL) | 53.2 ± 4.9 | 44.6 ± 5.1 a | 36.9 ± 4.1 ab | 104 ± 9.8 abc | 88.5 ± 7.9 abcd | 63.5 ± 5.8 abcde |
| CHOL (mg/dL) | 74.5 ± 5.8 | 66.3 ± 5.5 a | 59.6 ± 4.1 ab | 146 ± 11.4 abc | 103 ± 10.5 abcd | 81.5 ± 7.4 abcde | |
| LDL-c (mg/dL) | 45.6 ± 4.9 | 37.4 ± 4.1 a | 31.1 ± 3.4 ab | 83.4 ± 7.1 abc | 71.2 ± 7.1 abcd | 59.8 ± 6.1 abcde | |
| FFAs (µmol/L) | 512 ± 45.3 | 498 ± 42.2 | 523 ± 51.2 | 1032 ± 115 abc | 723 ± 62.4 abcd | 551 ± 44.8 abcde | |
| Liver | TGs (mg/g) | 4.7 ± 0.38 | 3.7 ± 0.41 a | 3.01 ± 0.29 ab | 7.8 ± 0.91 abc | 6.3 ± 0.54 abcd | 0.52 ± 85 abcde |
| CHOL (mg/g) | 2.7 ± 0.29 | 2.2 ± 0.21 a | 1.7 ± 0.27 ab | 5.1 ± 0.72 abc | 4.1 ± 0.62 abcd | 3.2 ± 0.26 abcde | |
| FFA (µmol/g) | 73.4 ± 8.1 | 78.5 ± 6.9 | 75.3 ± 6.1 | 149 ± 11.6 abc | 101 ± 13.2 abcd | 84.5 ± 7.8 abcde | |
| Faeces | TGs (mg/g) | 4.4 ± 0.35 | 4.7 ± 0.5 | 4.5 ± 0.71 | 12.3 ± 1.43 abc | 13.2 ± 1.93 abc | 11.9 ± 1.6 abc |
| CHOL (mg/g) | 5.8 ± 0.72 | 5.5 ± 0.78 | 6.1 ± 0.82 | 15.4 ± 1.7 abc | 13.8 ± 1.9 abc | 14.7 ± 1.8 abc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altamimy, K.M.; Alshammari, G.M.; Yagoub, A.E.A.; Albekairi, N.A.; Alshehri, S.; Saleh, A.; Yahya, M.A. Saudi Traditional Fermented Goat Milk Protects against Experimental Non-Alcoholic Fatty Liver Disease by Hypoglycaemic and Antioxidant Potentials. Fermentation 2022, 8, 735. https://doi.org/10.3390/fermentation8120735
Altamimy KM, Alshammari GM, Yagoub AEA, Albekairi NA, Alshehri S, Saleh A, Yahya MA. Saudi Traditional Fermented Goat Milk Protects against Experimental Non-Alcoholic Fatty Liver Disease by Hypoglycaemic and Antioxidant Potentials. Fermentation. 2022; 8(12):735. https://doi.org/10.3390/fermentation8120735
Chicago/Turabian StyleAltamimy, Khadijah M., Ghedeir M. Alshammari, Abu ElGasim A. Yagoub, Norah A. Albekairi, Samiyah Alshehri, Ali Saleh, and Mohammed Abdo Yahya. 2022. "Saudi Traditional Fermented Goat Milk Protects against Experimental Non-Alcoholic Fatty Liver Disease by Hypoglycaemic and Antioxidant Potentials" Fermentation 8, no. 12: 735. https://doi.org/10.3390/fermentation8120735
APA StyleAltamimy, K. M., Alshammari, G. M., Yagoub, A. E. A., Albekairi, N. A., Alshehri, S., Saleh, A., & Yahya, M. A. (2022). Saudi Traditional Fermented Goat Milk Protects against Experimental Non-Alcoholic Fatty Liver Disease by Hypoglycaemic and Antioxidant Potentials. Fermentation, 8(12), 735. https://doi.org/10.3390/fermentation8120735