3.3. Optimization of Medium Composition for SOD Production at Flask-Scale
To improve the cell growth and SOD production of
S. cerevisiae TBRC657, an optimization of the fermentation process involving carbon sources, nitrogen sources or inducer types and concentrations were individually investigated at flask-scale. After culturing the selected strain in different carbon-source base media, the cell weight and SOD activity were also different (
Table 3). When molasses was applied as the major carbon-source, the growth of
S. cerevisiae TBRC657 and its SOD production were the highest (1.86 g and 328.24 U/mL, respectively). The second and third most effective carbon-sources for a higher SOD activity were glucose (normal YPD) and sucrose, respectively. In contrast, lower values of SOD production were obtained when culturing
S. cerevisiae TBRC657 with corn starch (18.19 U/mL). According to the results, molasses showed the highest potential for use as a low-cost carbon-source for the enzyme production, which increased the SOD activity up to 1.58-fold when compared to the normal YPD (328.24 and 208.18 U/mL, respectively). In addition, molasses obtained from sugarcane mills represents a low-cost source of waste containing a high concentration of mono- and disaccharides such as glucose, sucrose and fructose (approximately 50%(
w/w)), which microorganisms can utilize for their growth and producing the essential metabolites [
31,
32]. As for the other carbon-sources, fructose and sucrose were also good sources for cell biomass and SOD production due to the ability of the yeast cell to utilize those carbon-sources. On the other hand,
S. cerevisiae TBRC657 could not utilize starch for its growth, which might require additional protocols such as applying starch-degrading enzymes to the system [
33]. Those additional protocols may affect to the overall cost of enzyme production and are not easy to operate at the pilot scale. Thus, the medium consisting of molasses as a major carbon-source gave the greatest values of cell weight and SOD activity and was used in further experiments.
To examine the optimum concentration of molasses as a major carbon-source,
S. cerevisiae TBRC657 was cultured at different concentrations of molasses ranging from 5.0% (
w/w)–30.0% (
w/w). The result showed that the highest cell growth was obtained when culturing
S. cerevisiae TBRC657 with 15.0%(
w/w) molasses, which provided a SOD production of 350.88 U/mL (
Table 4). On the other hand, a molasses concentration of 25.0% (
w/w) provided the highest SOD production of 590.87 U/mL. The second and third highest SOD productions were achieved with 30 (
w/w) and 20.0% (
w/w) molasses, respectively. Thus, 25.0% (
w/w) molasses was selected as the optimum concentration for use as a sole carbon-source for SOD production from
S. cerevisiae TBRC657.
Another essential factor which is involved in cell biomass and enzyme production is the nitrogen source. To improve growth and SOD production from
S. cerevisiae TBRC657, the culturing medium was supplemented with different nitrogen-sources, including organic and inorganic nitrogen sources, to achieve a similar amount of yeast cell and SOD production as obtained from normal YPD, as shown in
Table 5. After
S. cerevisiae TBRC657 was harvested and SOD was extracted inside the cells, yeast extract and soytone gave a similar growth of
S. cerevisiae TBRC657 when compared to normal YPD, as shown in
Table 5 (1.45, 1.30 and 1.28 g for yeast extract, soytone and normal YPD, respectively). According to the results, when applied to yeast extract as a sole nitrogen-source for SOD production,
S. cerevisiae TBRC657 was able to increase SOD production up to 1.24-fold when compared to normal YPD (262.92 and 211.53 U/mL, respectively). Thus, inorganic nitrogen-sources such as urea or sodium nitrate which were used in this study were not suitable nitrogen-sources for the selected strain due to a very low cell and SOD production of
S. cerevisiae TBRC657. Thus, yeast extract is the most common component applied as a nitrogen-source in culture media due to a high content of amino acids and essential nutrients for cell growth [
34,
35]. After varying the concentration of yeast extract ranging from 1.0% (
w/w)–5.0% (
w/w), the cell growth and SOD production were constant once 3.0% (
w/w) yeast extract was applied (data not shown), which would be used for further studies.
After investigating the medium composition for SOD production, molasses and yeast extract were found to be suitable carbon- and nitrogen sources for SOD production from
S. cerevisiae TBRC657. The optimal concentration of molasses and yeast extract were 25.0% (
w/w) and 3% (
w/w), respectively, which were used as the major composition of the optimized medium in
Table 6. To enhance SOD production, some chemical and physical conditions were tested to increase oxidative stress, leading to a higher production of SOD and other antioxidants, which will eliminate the excess free radicals inside the cell. According to previous reports, some chemicals such as hydrogen peroxide, menadione or acetic acid could at the appropriate concentration induce SOD productivity from many microorganisms [
36,
37,
38]. According to the results obtained, the optimized medium having molasses as major component, which consisted of many types of sugar and chemicals at high concentrations, acted as a hypertonic solution which caused osmotic pressure inside the cells [
39]. This condition will generate osmotic and oxidative stresses inside
S. cerevisiae TBRC657, resulting in an increase in SOD production. Before this experiment, varying the concentration of hydrogen peroxide and menadione, which maximize SOD production without any effect on the growth of the selected strain, was examined. The results exhibited that the suitable concentrations of hydrogen peroxide and menadione were 100 mM and 1 mM, respectively (data not shown). As expected, those inducers at the desired concentrations enhanced SOD production up to 1.37-fold compared to the non-induced control when culturing the selected strain in normal YPD. This result indicates that the chemicals used in this study significantly enhanced SOD production from
S. cerevisiae TBRC657. On the other hand, applying those inducers to the cell while culturing in the optimized medium did not enhance SOD production and also effected the growth of the yeast cells due to the decreasing value of total cell weight when compared to the non-induced culture, as shown in
Table 6. In addition, the optimized medium without any chemical induction increased SOD production from
S. cerevisiae TBRC657 up to 3.97-fold when compared to the normal YPD. Thus, the SOD production using the optimized medium provided many advantages, such as being easy to operate and not requiring any inducers, which might reduce the overall cost of enzyme production. According to the results obtained, the optimized medium was used for investigating SOD production in a 5 L bioreactor in further experiments.
3.5. SOD Formulation and Storage Stability
Before a thermal shift assay was conducted, the obtained SOD was partial purified by ammonium sulfate precipitation ranging from 40–60% of saturation, which exhibited the highest enzyme yield and purification factor. A DSF study of 4 different groups of additives—polymers and polyols, salts, sugar alcohols and sugars—at various concentrations, involving differences of conformational structure and biological function of the target enzymes, exhibited that each additive at various concentrations had a different effect on the structure of the partial purified SOD. Thus, the various values of ΔTm were revealed in this study. Positive ΔTm represents additives enabling protection of conformational structure of the target enzyme, whereas negative ΔTm represents the additives disrupting the conformational structure of the target enzyme [
40]. Among these additives, a mixture between the partial purified SOD and 1.0% (
w/v) trehalose exhibited the highest ΔTm of 15.17 °C compared to the unformulated SOD, as shown in
Table 8. The additives that provided the second and third highest ΔTm were 10.0% (
v/v) glycerol and 1.0% (
w/v) ammonium chloride, which exhibited ΔTm values of 14.00 °C and 13.50 °C, respectively, compared to the unformulated SOD. According to the results, most additives used in this study gave positive ΔTm values when compared to the unformulated control, suggesting that those additives were able to protect the obtained SOD and stabilize its conformational structure in a harsh environment. Thus, those additives can enhance the strength of the hydrophobic interactions between non-polar amino acid residues in the protein, leading to a more rigid protein, which promotes enzyme stability in those environments. The different groups of additives which are reported to provide a different function for stabilizing the conformational structure of the target enzyme were examined and mixed with the obtained enzyme. For example, sugars such as fructose, glucose or trehalose can enhance the bonding strength of hydrophobic interaction between the enzyme itself and the others, causing a stable structure of the target enzyme which can protect the enzyme from harsh environments. Salt solutions such as sodium chloride or ammonium chloride can protect the structure of the enzyme via competition in the binding between proteins and water molecules. Thus, water molecules will be removed from the target protein, forming a more compact structure. Some additives can form a hydrophobic interaction with the target enzyme, leading to reduced movement and also reduced water activity of the enzyme, which causes the formulated enzymes to be more stable in the environment [
41,
42,
43,
44]. Thus, these top 3 formulated SODs were selected and further analyzed for their SOD activities and storage stability at 4 °C for 6 months.
The top 3 formulates providing the highest ΔTm—1.0% (
w/v) trehalose, 10.0% (
v/v) glycerol and 1.0% (
w/v) ammonium chloride—were selected and measured for their SOD activities in order to investigate the effect of those additives on biological function. Thus, the suitable additives should provide a high ΔTm with the same level of SOD activity compared to the unformulated SOD. The results show that all 3 formulates exhibited an SOD activity of 971.42 U/mL, 988.14 U/mL and 1002.67 U/mL, respectively, whilst the unformulated SOD showed the same level of SOD activity of 982.44 U/mL (
Table 9). Thus, the results indicated that the selected additives were involved in the conformational structure of the target enzyme but did not affect the activity of the enzyme. The formulated SODs were stored for 6 months and the SOD activity from each formulate was measured every month compared to the unformulated SOD, which was set as 100.00% since the beginning of experiment. The results from all selected formulates stored for up to 6 months, show that all formulates, especially trehalose or glycerol, seemed to have a potential to stabilize SOD activity at storage conditions, resulting in more than 80% of the remaining SOD activity being detected when compared to the unformulated control, which dramatically decreased in activity since month 3 of the experiment. The remarkable increase in storage stability with the selected additives might be explained by preferential exclusion theory, suggesting that the addition of additives—trehalose or glycerol—to the enzyme solution segregates the water molecules away from the enzyme surface, resulting in a reduction of the hydration radius and increasing the compactness of the enzyme molecules which consequently stabilize the target enzyme. Thus, the enhancement of the conformational structure to enhance storage stability might require more than one group of additives which have different functions on the target enzyme [
41,
45,
46]. Mixing different groups of additives might have a synergistic effect, resulting in enhanced efficiency of storage stability of the target enzyme by altering its conformational structure, which can be detected by other biophysical techniques in further studies.
3.6. Cytotoxicity Activity and Protective Effect of SOD on Fibroblast Cells
The obtained SOD is expected to be used as a bioactive ingredient in the cosmetic and personal care product industries. For that reason, the cytotoxicity and proliferation of the obtained SOD from
S. cerevisiae TBRC657 was tested against fibroblast cells, which represent cells that synthesize the extracellular matrix and collagen in connective tissue. After 24 h of treatment, the percentage of living cells treated with an SOD dosage of 50 U and 100 U increased, while dramatically decreasing when treated with higher dosages (
Table 10). This result shows high cell viability and fibroblast cell proliferation when treated with a smaller dosage of SOD, with more than 80% of cell viability, which makes it declarable as a non-toxic substance to fibroblast cells. Then, the inhibitory concentration 50% or IC50, which indicates the concentration which is required to inhibit 50% of the proliferation, was examined, and the result showed that IC50 of the obtained SOD was approximately 2660 U, which is equal to 1.77 mg/mL protein of crude extract (data not shown). Thus, the results obtained indicate that a small dosage of the obtained SOD enhanced a proliferation of fibroblast cells. A treatment at 500 units of SOD to fibroblast cells exhibited a cell viability of 98.77% (
Table 10), which makes it declarable as non-toxic to fibroblast cells because more than 80% of cell viability was detected [
47,
48]. According to the results, the cell viability was predicted to be decreased when treating with higher dosages of crude SOD, indicating the limitations of crude SOD dosages.
Following this, the protective effect of the obtained SOD on fibroblast cells was observed. Firstly, the effect of hydrogen peroxide on cell morphology and viability was studied using SOD-untreated fibroblast cells. The morphology of those fibroblast cells directly treated with the highest concentration of hydrogen peroxide showed shrinking and no attachment to others, which is characteristic of programmed cell death with a viability of 53.50% compared to the control. Hence, adding an exogenous hydrogen peroxide caused the fibroblast cells to generate a high level of ROS inside the cells, followed by a programmed cell death phenomenon [
49]. Thus, the protective effect of SOD-treated cells on ROS was investigated. After incubating fibroblast cells with various dosages of SOD overnight, the cells were treated with exogenous hydrogen peroxide for 1 h, with amounts ranging from 12.5–200.0 µg/mL, causing ROS formation inside the fibroblast cells. Then, cell viability of all treatments was measured by MTT assays and compared to the untreated cells. According to
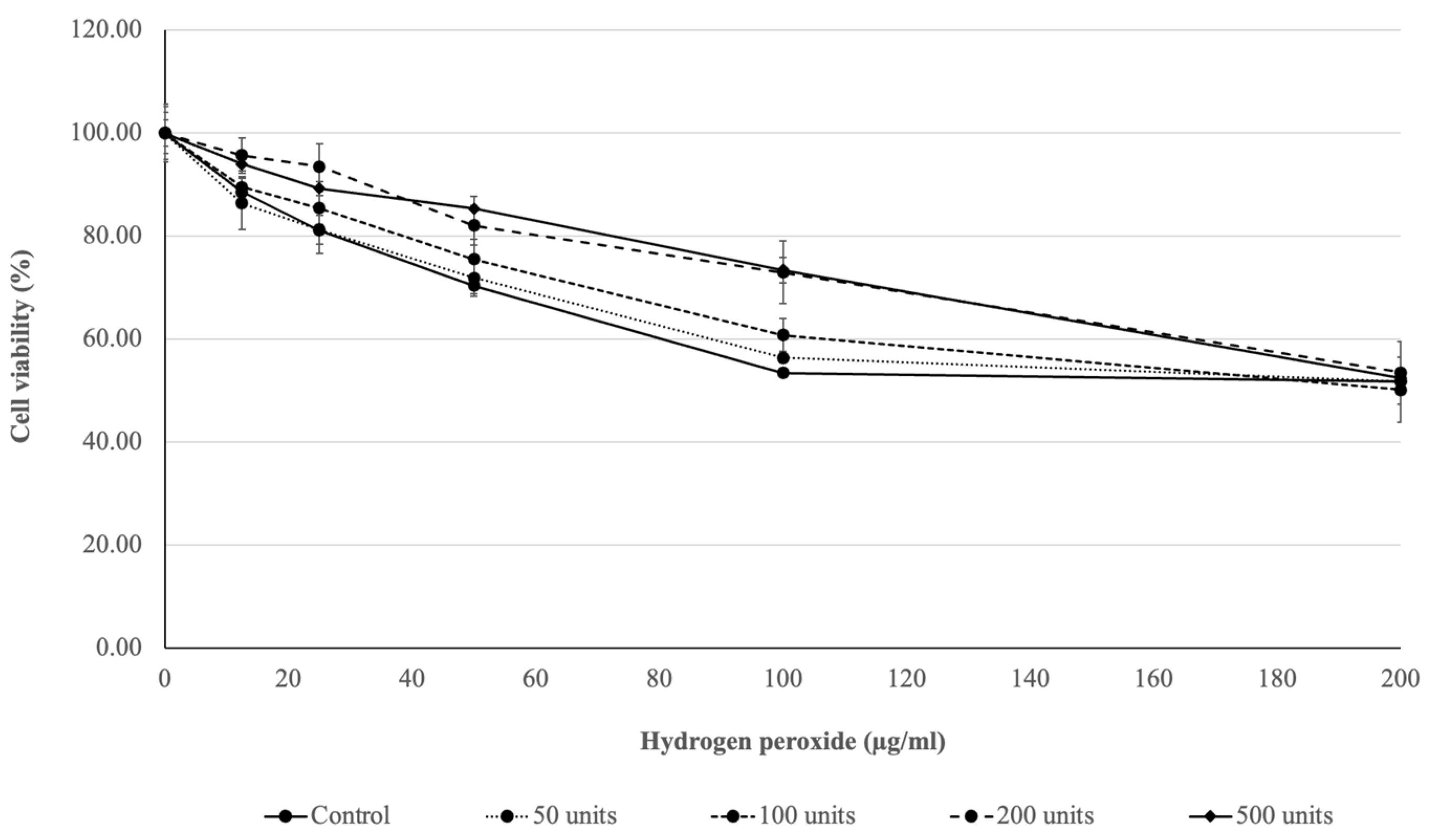
Figure 1, a high dosage of SOD (200 units and 500 units) enhanced the protective effect on ROS inside the cell generated by 100 µg/mL of hydrogen peroxide, with an approximate 20% increase in cell viability compared to the other treatments. This result indicates that the obtained SOD had a protective effect on exogenous hydrogen peroxide [
50]. On the other hand, 200 µg/mL of hydrogen peroxide was an excess concentration which triggered programmed cell death, even though the highest dosage of SOD was not sufficient [
51,
52]. This result indicates that SOD obtained from
S. cerevisiae TBRC657 plays an essential role in oxidative stress protection, with a protective effect on ROS inside the cells which can be used and applied in many industrial applications such as cosmetics and personal care products.


 ,
, 

{kind=link}