
Improvement of the Nutritional Quality of Psophocarpus tetragonolobus Tubers by Fermentation with Ruminal Crabtree-Negative Yeasts on the In Vitro Digestibility and Fermentation in Rumen Fluid

 , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location, Winged Bean Tubers, and Yeast
2.2. Yeast-Fermented Winged Bean Tubers
2.3. Evaluation of Microorganism Count, Fermentation Product, and Chemical Composition
2.4. Inoculum Preparation In Vitro Gas Technique
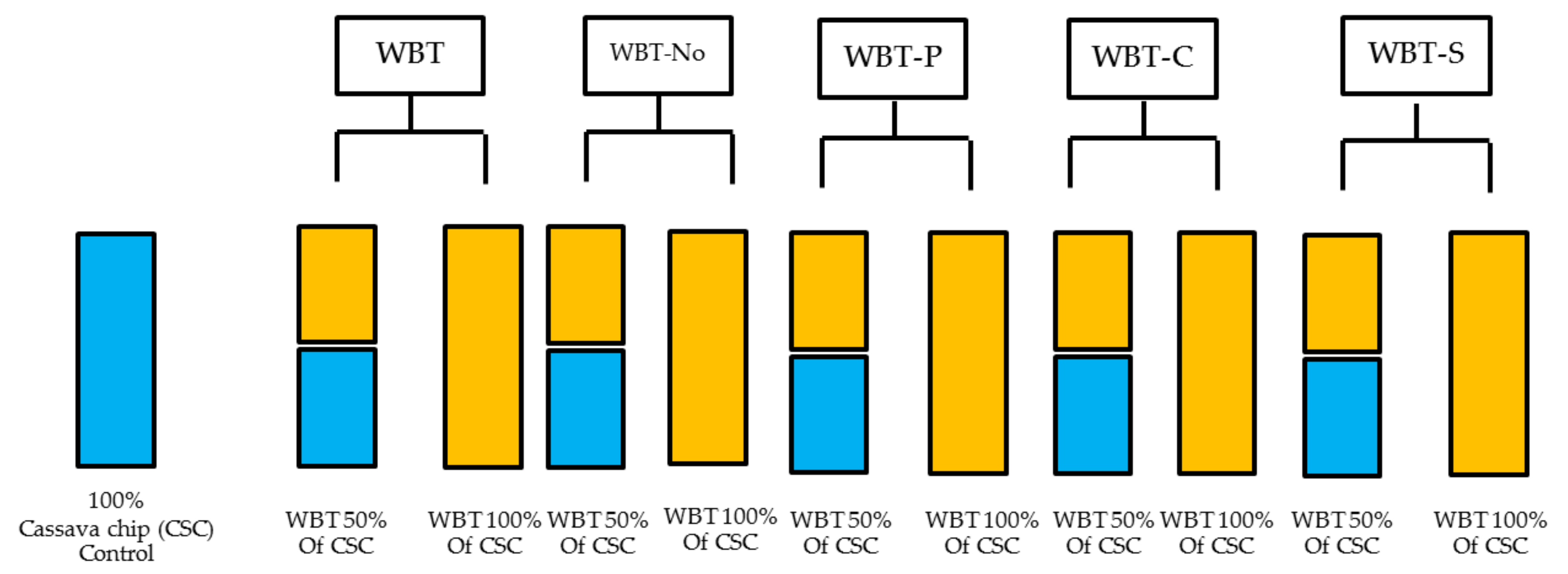
2.5. Dietary Treatments and Design
2.6. In Vitro Gas Production and Fermentation Characteristics
2.7. Statistical Analyses
3. Results
3.1. Chemical Contents of Fermented Winged Bean Tubers
3.2. Fermentation Characteristics and Microbial Counts in Products
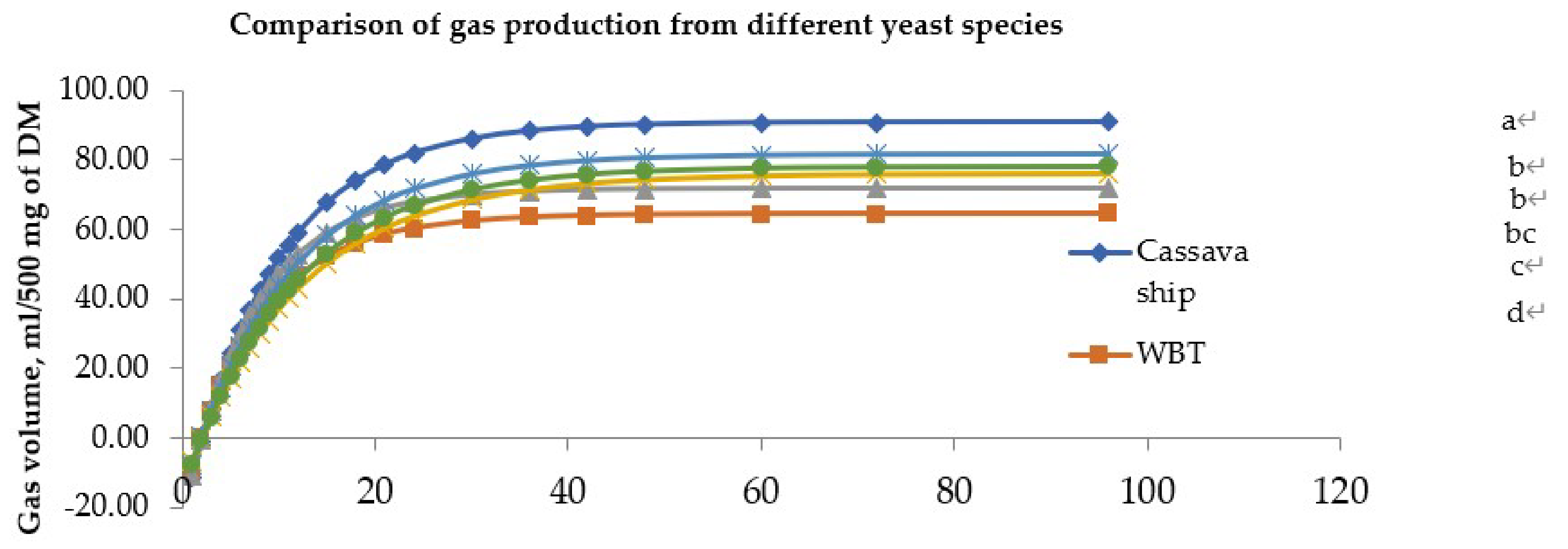
3.3. In Vitro Gas Production and the Model Parameters
3.4. In Vitro Degradability
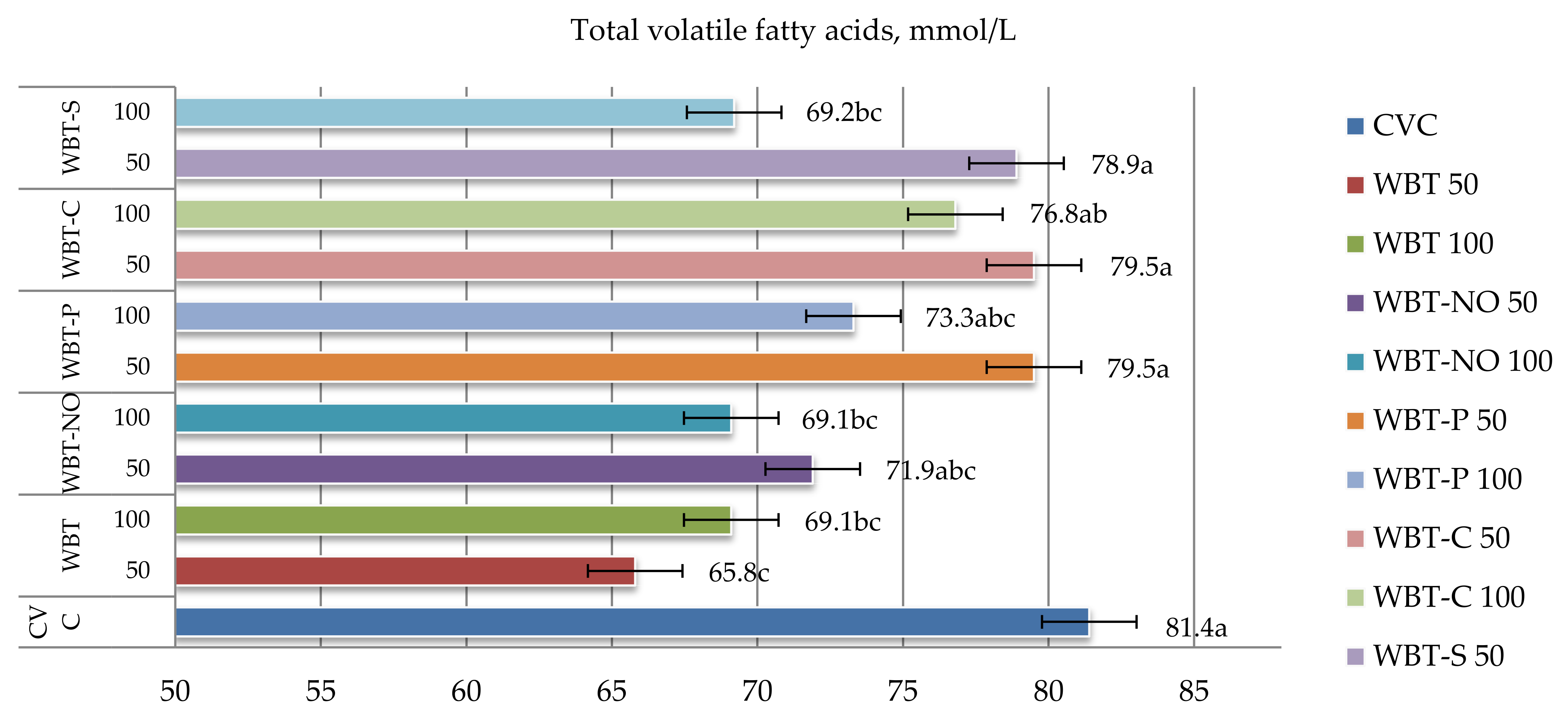
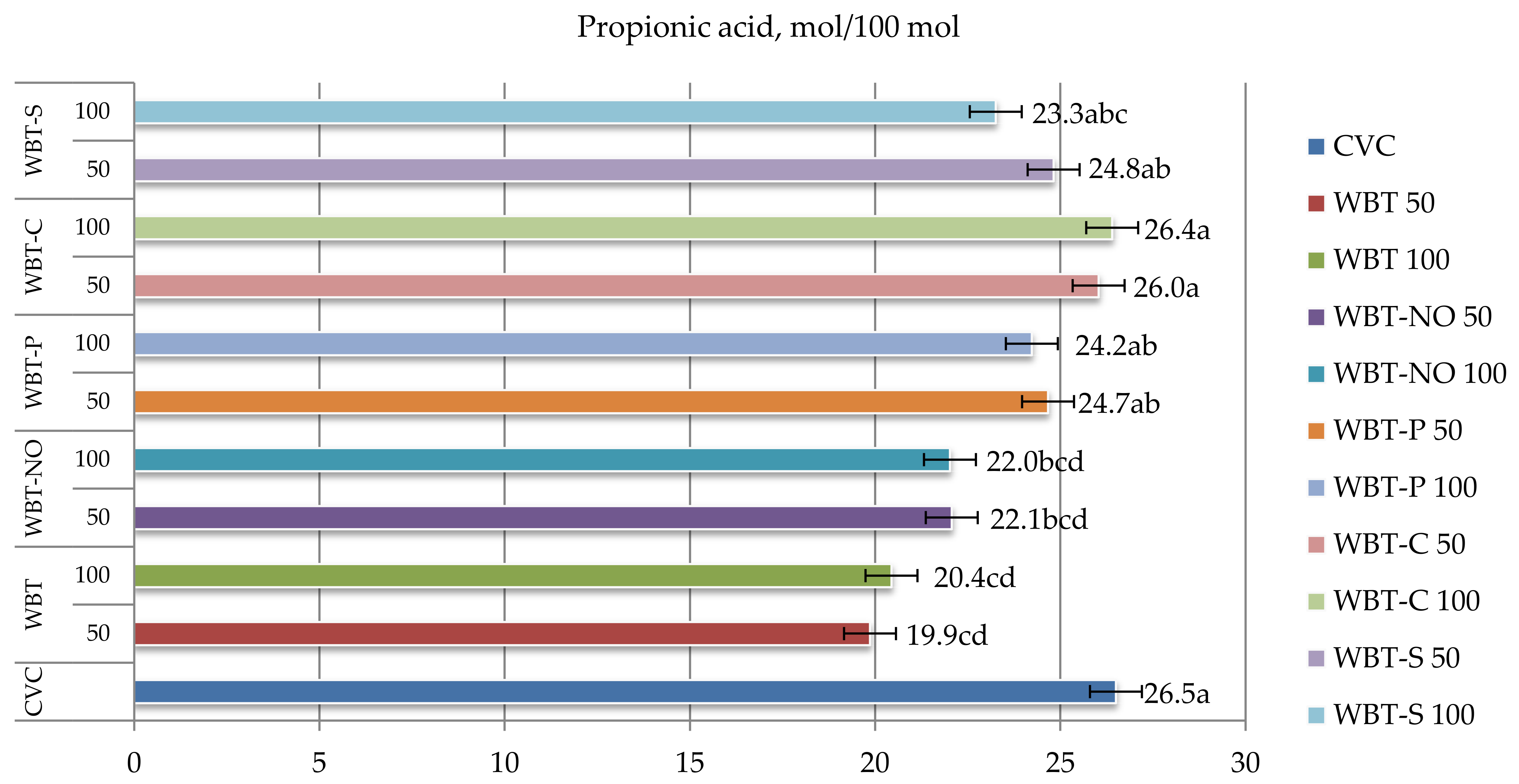
3.5. In Vitro Volatile Fatty Acid
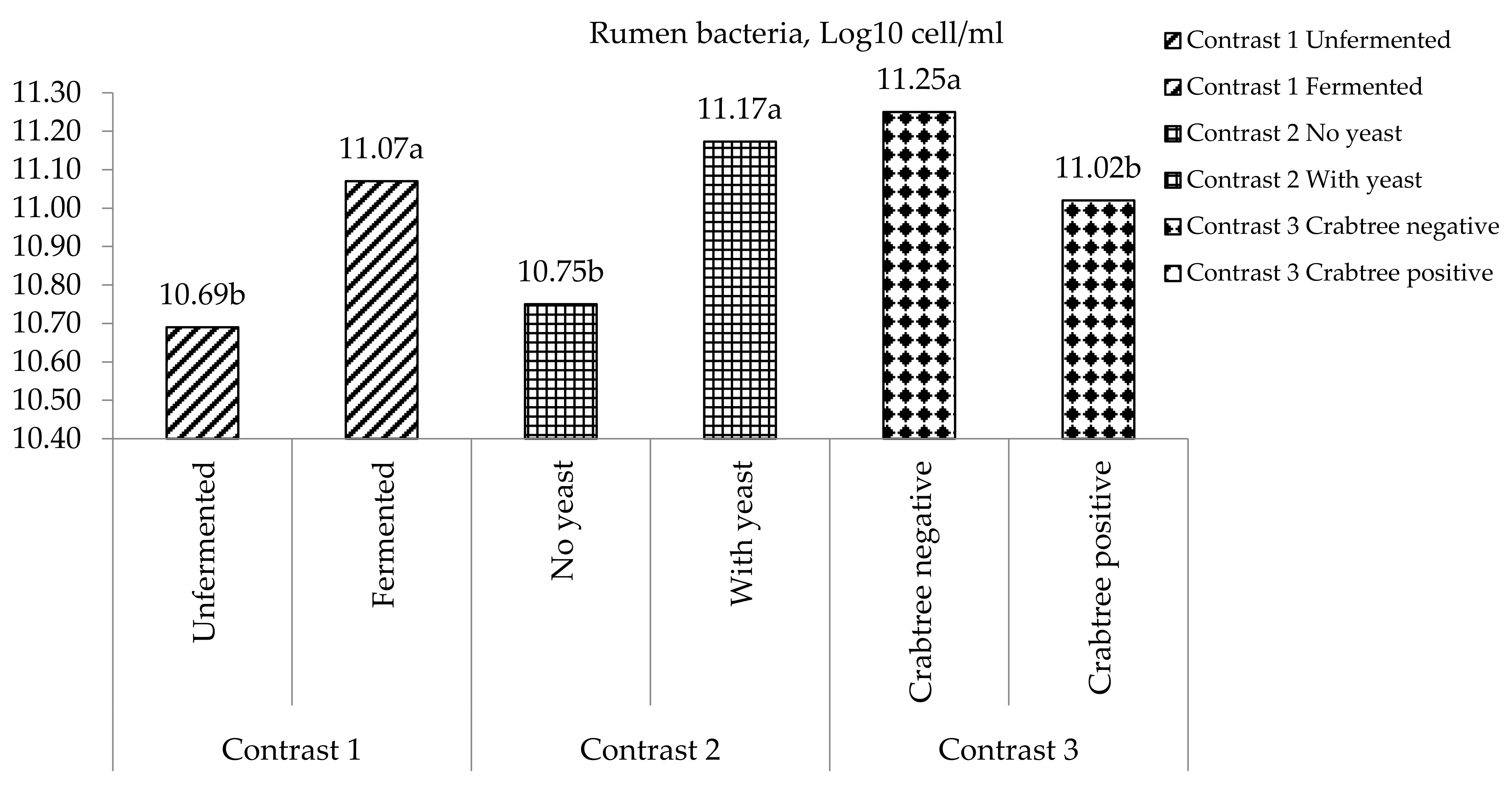
3.6. pH, Ammonia-Nitrogen and Bacteria Populations in the Rumen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wanapat, M.; Kang, S. Cassava chip (Manihot esculenta Crantz) as an energy source for ruminant feeding. Anim. Nutr. 2015, 1, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Aparecido, L.E.D.O.; Moraes, J.R.D.S.C.D.; de Meneses, K.C.; Lorençone, P.A.; Lorençone, J.A.; Souza, G.H.D.O.; Torsoni, G.B. Agricultural zoning as tool for expansion of cassava in climate change scenarios. Theor. Appl. Climatol. 2020, 142, 1085–1095. [Google Scholar] [CrossRef]
- Howeler, R. Cassava in Asia: Trends in cassava production, processing and marketing. In Proceedings of the Workshop on Partnership in Modern Science to Develop a Strong Cassava Commercial Sector in Africa and Appropriate Varieties by 2020, Bellagio, Italy, 2–6 May 2006. [Google Scholar]
- Trakarnpaiboon, S.; Srisuk, N.; Piyachomkwan, K.; Sakai, K.; Kitpreechavanich, V. Enhanced production of raw starch degrading enzyme using agro-industrial waste mixtures by thermotolerant Rhizopus microsporus for raw cassava chip saccharification in ethanol production. Prep. Biochem. Biotechnol. 2017, 47, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Tanzi, A.S.; Eagleton, G.E.; Ho, W.K.; Wong, Q.N.; Mayes, S.; Massawe, F. Winged bean (Psophocarpus tetragonolobus (L.) DC.) for food and nutritional security: Synthesis of past research and future direction. Planta 2019, 250, 911–931. [Google Scholar] [CrossRef] [Green Version]
- Kuswanto, A.N.; Saptadi, D.; Waluyo, B. Evaluation and selection on local strains of winged bean in Brawijaya University Indonesia. Trans. Persat. Genet. Malays. 2016, 3, 51–55. [Google Scholar]
- Lepcha, P.; Egan, A.N.; Doyle, J.J.; Sathyanarayana, N. A review on current status and future prospects of winged bean (Psophocarpus tetragonolobus) in Tropical agriculture. Plant Foods Hum. Nutr. 2017, 72, 225–235. [Google Scholar] [CrossRef]
- Sriwichai, S.; Monkham, T.; Sanitchon, J.; Jogloy, S.; Chankaew, S. Dual-purpose of the winged bean (Psophocarpus tetragonolobus (L.) DC.), the neglected Tropical legume, based on pod and tuber yields. Plants 2021, 10, 1746. [Google Scholar] [CrossRef]
- Wu, J.F. Energy value of cassava for young swine. J. Anim. Sci. 1991, 69, 1349–1353. [Google Scholar] [CrossRef] [Green Version]
- Kantha, S.S.; Erdman, J.W. The winged bean as an oil and protein source: A review. J. Am. Oil Chem. Soc. 1984, 61, 515–525. [Google Scholar] [CrossRef]
- Tan, N.H.; Wong, K.C. Thermal stability of trypsin inhibitor activity in winged bean (Psophocarpus tetragonolobus). J. Agric. Food Chem. 1982, 30, 1140–1143. [Google Scholar] [CrossRef]
- Pitakarnop, N. Production of pellets from winged bean tubers and seed cake for animal feed. In Proceedings of the 2nd International Symposium on Winged Bean, Peradeniya, Sri Lanka, 19–23 January 1981. [Google Scholar]
- Promkot, C.; Wanapat, M.; Mansathit, J. Effects of yeast fermented-cassava chip protein (YEFECAP) on dietary intake and milk production of Holstein crossbred heifers and cows during pre- and post-partum period. Livest. Sci. 2013, 154, 112–116. [Google Scholar] [CrossRef]
- Polyorach, S.; Wanapat, M.; Cherdthong, A. Influence of yeast fermented cassava chip protein (YEFECAP) and roughage to concentrate ratio on ruminal fermentation and microorganisms using in vitro gas production technique. Asian-Australas. J. Anim. Sci. 2014, 27, 36–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khampa, S.; Polsit, K.; Chuelong, S.; Siriuthane, T.; Ittarat, S.; Koatedoke, U.; Cherdthong, A. Supplementation of cassava and durian hull fermented yeast (Saccharomyces cerevisiae) on rumen fermentation and average daily gain in crossbred native cattle. Pak. J. Nutr. 2011, 10, 1121–1125. [Google Scholar] [CrossRef] [Green Version]
- Khampa, S.; Chuelong, S.; Siriuthane, T.; Polsit, K.; Ittharat, S.; Koatdoke, U.; Cherdthong, A. Supplementation levels of palm oil in yeast (Saccharomyces cerevisiae) culture fermented cassava pulp on rumen fermentation and average daily gain in crossbred native cattle. Pak. J. Nutr. 2011, 10, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Pattarajinda, V.; Sunato, S.; Lowilai, P.; Nontaso, N. Effect of yeast fermented ethanol waste on feed utilization and digestion in dairy cattle. Pak. J. Nutr. 2015, 14, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Shen, C.; Liu, Q.; Zhang, S.-L.; Shao, T.; Wang, C.; Wang, Y.X.; Xu, Q.F.; Huo, W.J. The effect of lactic acid bacteria inoculums on in vitro rumen fermentation, methane production, ruminal cellulolytic bacteria populations and cellulase activities of corn stover silage. J. Integr. Agric. 2020, 19, 838–847. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Walker, N.; Bach, A. Effects of active dry yeasts on the rumen microbial ecosystem: Past, present and future. Anim. Feed Sci. Technol. 2008, 145, 5–26. [Google Scholar] [CrossRef]
- Crabtree, H.G. Observations on the carbohydrate metabolism of tumours. Biochem. J. 1929, 23, 536–545. [Google Scholar] [CrossRef]
- De Deken, R. The Crabtree effect: A regulatory system in yeast. Microbiology 1966, 44, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Van Urk, H.; Voll, W.L.; Scheffers, W.A.; Van Dijken, J.P. Transient-state analysis of metabolic fluxes in Crabtree-positive and Crabtree-negative yeasts. Appl. Environ. Microbiol. 1990, 56, 281–287.23. [Google Scholar] [CrossRef] [Green Version]
- Suntara, C.; Cherdthong, A.; Wanapat, M.; Uriyapongson, S.; Leelavatcharamas, V.; Sawaengkaew, J.; Chanjula, P.; Foiklang, S. Isolation and characterization of yeasts from rumen fluids for potential use as additives in ruminant feeding. Vet. Sci. 2021, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Suntara, C.; Cherdthong, A.; Uriyapongson, S.; Wanapat, M.; Chanjula, P. Comparison effects of ruminal Crabtree-negative yeasts and Crabtree-positive yeasts for improving ensiled rice straw quality and ruminal digestion using in vitro gas production. J. Fungi. 2020, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Suntara, C.; Cherdthong, A.; Uriyapongson, S.; Wanapat, M.; Chanjula, P. Novel Crabtree negative yeast from rumen fluids can improve rumen fermentation and milk quality. Sci. Rep. 2021, 11, 6236. [Google Scholar] [CrossRef] [PubMed]
- Yitbarek, M.B.; Tamir, B. Silage additives. Open J. Appl. Sci. 2014, 4, 258–274. [Google Scholar] [CrossRef] [Green Version]
- Kozaki, M.; Uchimura, T.; Okada, S. Experimental Manual of Lactic Acid Bacteria; Asakurasyoten: Tokyo, Japan, 1992; pp. 34–37. [Google Scholar]
- Fawcett, J.K.; Scott, J.E. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- The Working Committee of Thai Feeding Standard for Ruminant (WTSR). Nutrient Requirements of Beef Cattle in Indochinese Peninsula, 1st ed.; Klungnanavitthaya Press: Khon Kaen, Thailand, 2010. [Google Scholar]
- Menke, K.H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Dairy Cattle, 7th ed.; National Research Council; The National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Xia, C.; Rahman, M.A.U.; Yang, H.; Shao, T.; Qiu, Q.; Su, H.; Cao, B. Effect of increased dietary crude protein levels on production performance, nitrogen utilisation, blood metabolites and ruminal fermentation of Holstein bulls. Asian-Australas. J. Anim. Sci. 2018, 31, 1643–1653. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemist (AOAC). The Official Methods of Analysis of the Association of Official Analytical Chemist, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1998. [Google Scholar]
- Tilley, J.; Terry, R. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994; p. 476. [Google Scholar] [CrossRef]
- Schofield, P.; Pitt, R.E.; Pell, A.N. Kinetics of fiber digestion from in vitro gas production. J. Anim. Sci. 1994, 72, 2980–2991. [Google Scholar] [CrossRef]
- Statistical Analysis Systems (SAS). SAS/STAT User’s Guide. In Statistical Analysis Systems Institute, 2nd ed.; Version 9; SAS Institute Inc.: Cary, NC, USA, 2002. [Google Scholar]
- Steel, R.G.; Torrie, J.H. Principles and Procedures of Statistics; McGraw-Hill Book Co. Inc.: New York, NY, USA, 1980; p. 633. [Google Scholar] [CrossRef]
- Kang, S.; Wanapat, M.; Nunoi, A. Effect of urea and molasses supplementation on quality of cassava top silage. J. Anim. Feed Sci. 2018, 27, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Wanapat, M.; Polyorach, S.; Boonnop, K.; Mapato, C.; Cherdthong, A. Effects of treating rice straw with urea or urea and calcium hydroxide upon intake, digestibility, rumen fermentation and milk yield of dairy cows. Livest. Sci. 2009, 125, 238–243. [Google Scholar] [CrossRef]
- Khampa, S.; Chaowarat, P.; Singhalert, R.; Wanapat, M. Supplementation of yeast fermented cassava chip as a replacement concentrate on rumen fermentation efficiency and digestibility of nutrients in cattle. Asian J. Anim. Sci. 2009, 3, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Broach, J.R. Nutritional control of growth and development in yeast. Genetics 2012, 192, 73–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhou, P.; Chen, Y.; Cao, Q.; Liu, X.; Li, D. The production of single cell protein from biogas slurry with high ammonia-nitrogen content by screened Nectaromyces rattus. Poult. Sci. 2021, 100, 101334. [Google Scholar] [CrossRef]
- Pronk, J.T.; Yde Steensma, H.; van Dijken, J.P. Pyruvate metabolism in Saccharomyces cerevisiae. Yeast 1996, 12, 1607–1633. [Google Scholar] [CrossRef]
- Wardrop, F.; Liti, G.; Cardinali, G.; Walker, G. Physiological responses of Crabtree positive and Crabtree negative yeasts to glucose upshifts in a chemostat. Ann. Microbiol. 2004, 54, 103–114. [Google Scholar]
- McDonald, P.; Henderson, A.; Heron, S. The Biochemistry of Silage; Chalcombe Publications: Southampton, UK, 1991. [Google Scholar]
- Cherdthong, A.; Suntara, C.; Khota, W.; Wanapat, M. Feed utilization and rumen fermentation characteristics of Thai-indigenous beef cattle fed ensiled rice straw with Lactobacillus casei TH14, molasses, and cellulase enzymes. Livest. Sci. 2021, 245, 104405. [Google Scholar] [CrossRef]
- Jianxin, L.; Jun, G. Ensiling crop residues. In FAO Animal Production and Health Paper (FAO); Publishing and Multimedia Service, Information Division, Food and Agriculture Organization of the United Nations: Rome, Italy, 2002. [Google Scholar]
- Habeeb, A.A.M. Importance of yeast in ruminants feeding on production and reproduction. Ecol. Evol. 2017, 2, 49. [Google Scholar] [CrossRef]
- Sommart, K.; Parker, D.S.; Rowlinson, P.; Wanapat, M. Fermentation characteristics and microbial protein synthesis in an in vitro system using cassava, rice straw and dried Ruzi grass as substrates. Asian-Australas. J. Anim. Sci. 2000, 13, 1084–1093. [Google Scholar] [CrossRef]
- Makkar, H.P. Recent advances in the in vitro gas method for evaluation of nutritional quality of feed resources. Assessing quality and safety of animal feeds. FAO Anim. Prod. Health Pap. 2004, 160, 55–88. [Google Scholar]
- Wang, Z.; He, Z.; Beauchemin, K.A.; Tang, S.; Zhou, C.; Han, X.; Wang, M.; Kang, J.; Odongo, N.E.; Tan, Z. Evaluation of Different Yeast Species for Improving In vitro Fermentation of Cereal Straws. Asian-Australas. J. Anim. Sci. 2016, 29, 230–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaucheyras-Durand, F.; Fonty, G. Yeasts in ruminant nutrition. Experiences with a live yeast product. Kraftfutter 2002, 85, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Cherdthong, A.; Prachumchai, R.; Supapong, C.; Khonkhaeng, B.; Wanapat, M.; Foiklang, S.; Milintawisamai, N.; Gunun, N.; Gunun, P.; Chanjula, P.; et al. Inclusion of yeast waste as a protein source to replace soybean meal in concentrate mixture on ruminal fermentation and gas kinetics using in vitro gas production technique. Anim. Prod. Sci. 2018, 59, 1682–1688. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Chevaux, E.; Martin, C.; Forano, E. Use of yeast probiotics in ruminants: Effects and mechanisms of action on rumen pH, fibre degradation, and microbiota according to the diet. In Biology; IntechOpen: London, UK, 2012; pp. 119–152. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, R.; Chaudhry, A.S. Methods to study degradation of ruminant feeds. Nutr. Res. Rev. 2008, 21, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef]
- Mutsvangwa, T.; Edwards, I.E.; Topps, J.H.; Paterson, G.F.M. The effect of dietary inclusion of yeast culture (Yea-Sacc) on patterns of rumen fermentation, food intake and growth of intensively fed bulls. Anim. Sci. 1992, 55, 35–40. [Google Scholar] [CrossRef]
- Křížová, L.; Richter, M.; Trináctý, J.; Říha, J.; Kumprechtova, D. The effect of feeding live yeast cultures on ruminal pH and redox potential in dry cows as continuously measured by a new wireless device. Czech J. Anim. Sci. 2011, 56, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Elanthamil, R.; Bandeswaran, C.; Bharthidhasan, A.; Valli, C. Effect of Saccharomyces cerevisiae supplementation to paddy straw based substrate on in vitro rumen fermentation parameters. Indian J. Anim. Nutr. 2018, 35, 121–124. [Google Scholar] [CrossRef]
- Ren, H.; Su, X.; Bai, H.; Yang, Y.; Wang, H.; Dan, Z.; Lu, J.; Wu, S.; Cai, C.; Cao, Y.; et al. Specific enrichment of microbes and increased ruminal propionate production: The potential mechanism underlying the high energy efficiency of Holstein heifers fed steam-flaked corn. AMB Express 2019, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Yang, C.; Dong, L.; Diao, Q.; Si, B.; Ma, J.; Tu, Y. Rumen fermentation characteristics in pre- and post-weaning calves upon feeding with mulberry leaf flavonoids and Candida tropicalis individually or in combination as a supplement. Animals 2019, 9, 990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, T.R. Tropical Animal Feeding. A Manual for Research Workers; Paper No. 126; FAO Animal Production and Health: Rome, Italy, 1995; p. 305. [Google Scholar]
- Wanapat, M.; Boonnop, K.; Promkot, C.; Cherdthong, A. Effects of alternative protein sources on rumen microbes and productivity of dairy cows. Maejo Int. J. Sci. 2011, 5, 13. [Google Scholar]
- Leng, R.A. Factors affecting the utilization of ‘poor-quality’ forages by ruminants particularly under Tropical conditions. Nutr. Res. Rev. 1990, 3, 277–303. [Google Scholar] [CrossRef]
- Anantasook, N.; Wanapat, M.; Cherdthong, A.; Gunun, P. Effect of plants containing secondary compounds with palm oil on feed intake, digestibility, microbial protein synthesis and microbial population in dairy cows. Asian-Australas. J. Anim. Sci. 2013, 26, 820–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherdthong, A.; Khonkhaeng, B.; Seankamsorn, A.; Supapong, C.; Wanapat, M.; Gunun, N.; Gunun, P.; Chanjula, P.; Polyorach, S. Effects of feeding fresh cassava root with high-sulfur feed block on feed utilization, rumen fermentation, and blood metabolites in Thai native cattle. Trop. Anim. Health Prod. 2018, 50, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Anantasook, N.; Wanapat, M.; Cherdthong, A.; Gunun, P. Changes of microbial population in the rumen of dairy steers as influenced by plant containing tannins and saponins and roughage to concentrate ratio. Asian-Australas. J. Anim. Sci. 2013, 26, 1583–1591. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Treatments | Process | Additives | Yeast | SEM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WBT | WBT-NO | WBT-P | WBT-C | WBT-S | UWBT | FWBT | No | Yeast | C- | C+ | |||
| Dry matter, g/kg | 435.2 a | 296.7 b | 258.6 c | 266.3 c | 262.5 c | 435.2 a | 271.0 b | 296.7 a | 262.4 b | 262.43 | 262.45 | 5.36 | p < 0.01 |
| Nutrient content, g/kg DM | |||||||||||||
| Organic matter | 963.2 a | 954.5 c | 960.3 b | 955.5 c | 952.8 d | 963.2 a | 955.8 b | 954.5 b | 956.2 a | 957.9 a | 952.8 b | 0.33 | p < 0.01 |
| Ether extract | 3.79 c | 5.43 b | 5.46 b | 6.69 a | 5.51 b | 3.79 b | 5.77 a | 5.43 | 5.88 | 6.07 | 5.51 | 0.22 | p < 0.01 |
| Crude protein | 199.1 b | 208.1 ab | 213.7 a | 217.7 a | 212.7 a | 199.1 b | 213.1 a | 208.1 | 214.7 | 215.7 | 212.7 | 3.46 | 0.03 |
| Neutral detergent fiber | 180.3 | 180.7 | 173.1 | 176.5 | 181.6 | 180.3 | 178.0 | 180.7 | 177.1 | 174.8 | 181.6 | 2.35 | 0.19 |
| Acid detergent fiber | 65.2 | 63.6 | 56.9 | 62.4 | 63.2 | 65.2 | 61.6 | 63.6 | 60.9 | 59.7 | 63.2 | 2.48 | 0.30 |
| Items | Treatments | Additives | Yeast | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| WBT-NO | WBT-P | WBT-C | WBT-S | No | Yeast | C- | C+ | |||
| pH | 4.62 b | 4.28 d | 4.49 c | 5.56 a | 4.62 b | 4.78 a | 4.39 b | 5.56 a | 0.05 | p < 0.01 |
| Organic acids content (g/kg DM) | ||||||||||
| Ammonia nitrogen | 0.82 c | 0.73 c | 0.96 b | 1.29 a | 0.82 b | 0.99 a | 0.85 b | 1.29 a | 0.02 | p < 0.01 |
| Lactic acid | 4.87 ab | 5.63 a | 4.33 b | 4.21 b | 4.87 | 4.73 | 4.98 | 4.21 | 0.23 | 0.038 |
| Acetic acid | 2.05 | 1.70 | 1.64 | 1.67 | 2.05 a | 1.67 b | 1.67 | 1.67 | 0.08 | 0.053 |
| Propionic acid | 0.08 b | 0.08 b | ND | 0.31 a | 0.08 b | 0.13 a | 0.04 b | 0.31 a | 0.01 | p < 0.01 |
| Microbial count Log10 (cfu/g fresh matter) | ||||||||||
| Lactic acid bacteria | 8.67 a | 8.72 a | 8.23 b | 8.33 b | 8.67 a | 8.43 b | 8.48 a | 8.33 b | 0.03 | p < 0.01 |
| Yeast | 3.32 b | 5.59 a | 4.95 a | 4.91 a | 3.32 b | 5.15 a | 5.27 | 4.91 | 0.19 | p < 0.01 |
| Aerobic bacteria | 3.05 a | 1.09 b | 1.26 b | 1.32 b | 3.05 a | 1.22 b | 1.17 | 1.32 | 0.06 | p < 0.01 |
| Coliform | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Yeast Species | WBT Level | Gas Kinetics 1 | ||
|---|---|---|---|---|
| Vf | k | L | ||
| Cassava chips | 100 | 86.0 cd | 0.110 | 2.09 |
| WBT | 50 | 61.5 f | 0.134 a | 2.00 |
| 100 | 52.3 g | 0.131 a | 2.00 | |
| WBT-NO | 50 | 88.0 bcd | 0.124 ab | 2.07 |
| 100 | 55.9 fg | 0.144 a | 2.03 | |
| WBT-P | 50 | 91.7 abcd | 0.074 c | 1.97 |
| 100 | 96.4 a | 0.096 bc | 2.05 | |
| WBT-C | 50 | 85.3 c | 0.102b c | 2.00 |
| 100 | 75.0 e | 0.089 c | 1.85 | |
| WBT-S | 50 | 95.6 ab | 0.091 c | 2.13 |
| 100 | 93.4 abc | 0.085 c | 2.07 | |
| SEM | 2.50 | 0.01 | 0.04 | |
| Cassava chips vs. Others | 0.0214 | 0.96 | 0.07 | |
| Interaction | p < 0.01 | p < 0.01 | 0.47 | |
| Comparison | ||||
| Yeast species | WBT | 56.9 | 0.133 | 2.00 bc |
| WBT-NO | 72.0 | 0.134 | 2.05 ab | |
| WBT-P | 94.1 | 0.085 | 2.01 b | |
| WBT-C | 80.2 | 0.096 | 1.93 c | |
| WBT-SC | 94.5 | 0.088 | 2.10 a | |
| WBT level | 50 | 84.4 | 0.105 | 2.03 |
| 100 | 74.6 | 0.109 | 2.00 | |
| Contrast 1 | Unfermented | 56.9 b | 0.133 a | 2.00 |
| Fermented | 85.2 a | 0.101 b | 2.02 | |
| Contrast 2 | No yeast | 71.9 b | 0.134 a | 2.05 |
| With yeast | 89.6 a | 0.089 b | 2.01 | |
| Contrast 3 | Crabtree-negative | 87.1 b | 0.090 | 1.97 b |
| Crabtree-positive | 94.5 a | 0.088 | 2.10 a | |
| Contrast 4 | P. kudriazevii KKU20 | 94.1 a | 0.085 | 2.01 a |
| C. tropicalis KKU20 | 80.2 b | 0.096 | 1.93 b | |
| Yeast Species | WBT Level | Degradability at 12 h (g/kg Dry Matter) | Degradability at 24 h (g/kg Dry Matter) | ||||
|---|---|---|---|---|---|---|---|
| IVDMD | IVOMD | IVNDFD | IVDMD | IVOMD | IVNDFD | ||
| Cassava chips | 100 | 541.5 | 589.2 | 469.9 b | 712.8 | 767.3 | 609.8 |
| WBT | 50 | 543.1 de | 566.1 bc | 522.9 a | 686.4 d | 729.1 | 633.0 |
| 100 | 528.5 e | 563.5 c | 523.0 a | 706.1 bc | 743.8 | 634.5 | |
| WBT-NO | 50 | 548.5 de | 571.6 bc | 538.0 a | 698.6 cd | 744.0 | 640.8 |
| 100 | 593.9 abc | 592.2 ab | 511.6 a | 698.1 cd | 748.9 | 628.8 | |
| WBT-P | 50 | 617.9 a | 604.2 a | 531.7 a | 714.5 abc | 767.6 | 614.9 |
| 100 | 574.3 abcd | 598.7 a | 533.6 a | 723.3 ab | 764.1 | 663.3 | |
| WBT-C | 50 | 611.0 ab | 586.9 abc | 541.3 a | 721.1 ab | 770.5 | 645.9 |
| 100 | 570.5 abcd | 587.6 abc | 520.5 a | 713.7 abc | 750.2 | 642.2 | |
| WBT-S | 50 | 542.5 de | 584.6 abc | 523.5 a | 725.3 a | 737.6 | 625.1 |
| 100 | 550.3 de | 580.8 abc | 517.5 a | 700.7 cd | 749.7 | 640.0 | |
| SEM | 12.5 | 9.36 | 15.1 | 7.1 | 13.5 | 19.8 | |
| Cassava chips vs. Others | 0.07 | 0.58 | p < 0.01 | 0.60 | 0.27 | 0.37 | |
| Interaction | p < 0.01 | p < 0.01 | 0.74 | p < 0.05 | 0.19 | 0.81 | |
| Comparison | |||||||
| Yeast species | WBT | 535.8 | 564.8 | 523.0 | 696.2 | 737.5 | 633.8 |
| WBT-NO | 571.2 | 581.9 | 524.8 | 698.3 | 736.4 | 634.8 | |
| WBT-P | 596.1 | 601.4 | 532.6 | 718.9 | 765.8 | 639.1 | |
| WBT-C | 590.8 | 587.3 | 530.9 | 717.4 | 760.4 | 644.0 | |
| WBT-SC | 546.4 | 582.7 | 520.5 | 713.0 | 743.7 | 632.5 | |
| WBT level | 50 | 572.6 | 582.7 | 531.5 | 709.2 | 749.8 | 631.9 |
| 100 | 563.5 | 584.6 | 521.2 | 708.4 | 751.3 | 641.8 | |
| Contrast 1 | Unfermented | 535.8 b | 564.8 b | 523.0 | 696.2 b | 736.5 | 633.8 |
| Fermented | 576.1 a | 588.3 a | 527.2 | 711.9 a | 754.1 | 637.6 | |
| Contrast 2 | No yeast | 571.2 | 581.9 | 524.8 | 698.3 b | 746.4 | 634.8 |
| With yeast | 577.8 | 590.5 | 528.0 | 716.4 a | 756.6 | 638.6 | |
| Contrast 3 | Crabtree-negative | 593.5 a | 594.3 | 531.7 | 718.1 | 763.1 | 641.6 |
| Crabtree-positive | 546.4 b | 582.7 | 520.5 | 713.0 | 743.7 | 632.5 | |
| Contrast 4 | P. kudriazevii KKU20 | 596.1 | 601.4 | 532.6 | 718.9 | 765.8 | 639.1 |
| C. tropicalis KKU20 | 590.8 | 587.3 | 530.9 | 717.4 | 760.4 | 644.0 | |
| Yeast Species | WBT Level | Acetic Acid, mol/100 mol | Butyric Acid, mol/100 mol | ||||
|---|---|---|---|---|---|---|---|
| 4 h | 8 h | Mean | 4 h | 8 h | Mean | ||
| Cassava chips | 100 | 63.2 | 65.2 | 64.2 | 10.3 | 8.3 | 9.3 |
| WBT | 50 | 66.5 | 70.8 a | 68.6 | 13.3 a | 9.7 | 11.5 ab |
| 100 | 65.6 | 71.6 a | 68.6 | 12.1 ab | 9.8 | 10.9 abc | |
| WBT-NO | 50 | 65.2 | 65.1 b | 65.2 | 13.1 a | 12.4 | 12.8 a |
| 100 | 62.2 | 70.7 a | 66.4 | 13.8 a | 9.3 | 11.6 ab | |
| WBT-P | 50 | 66.3 | 65.3 b | 65.8 | 10.2 bcd | 8.9 | 9.5 bc |
| 100 | 69.6 | 64.1 b | 66.8 | 8.7 d | 9.2 | 8.9 c | |
| WBT-C | 50 | 64.4 | 65.1 b | 64.7 | 9.9 cd | 8.5 | 9.2 bc |
| 100 | 65.1 | 64.9 b | 65.0 | 8.5 d | 8.8 | 8.6 c | |
| WBT-S | 50 | 66.0 | 64.8 b | 65.4 | 11.1 bc | 8.5 | 9.b c |
| 100 | 67.2 | 67.6 ab | 67.4 | 8.d | 9.9 | 9.3 bc | |
| SEM | 1.24 | 1.33 | 1.02 | 0.96 | 1.04 | 0.68 | |
| Cassava chips vs. Others | 0.12 | 0.22 | 0.06 | 0.29 | 0.30 | 0.21 | |
| Interaction | 0.82 | p < 0.01 | 0.18 | p < 0.01 | 0.69 | p < 0.05 | |
| Comparison | |||||||
| Process | WBT | 66.0 | 71.2 | 68.6 | 12.7 | 9.8 | 11.2 |
| WBT-NO | 63.7 | 67.9 | 65.8 | 13.5 | 10.9 | 12.2 | |
| WBT-P | 68.0 | 64.7 | 66.3 | 9.4 | 9.0 | 9.2 | |
| WBT-C | 64.7 | 65.0 | 64.9 | 9.2 | 8.6 | 8.9 | |
| WBT-SC | 66.6 | 66.2 | 66.4 | 9.9 | 9.2 | 9.6 | |
| WBT level | 50 | 65.7 | 66.2 | 65.9 | 11.5 | 9.6 | 10.6 |
| 100 | 65.9 | 67.8 | 66.9 | 10.4 | 9.4 | 9.9 | |
| Contrast 1 | Unfermented | 66.0 | 71.2 a | 68.6 a | 12.7 a | 9.8 | 11.2 a |
| Fermented | 65.8 | 65.9 b | 65.9 b | 10.5 b | 9.4 | 9.9 b | |
| Contrast 2 | No yeast | 63.7 b | 67.9 | 65.8 | 13.5 a | 10.9 a | 12.2 a |
| With yeast | 66.4 a | 65.3 | 65.9 | 9.5b | 8.9 b | 9.2 b | |
| Contrast 3 | Crabtree-negative | 66.4 | 64.8 | 65.6 | 9.3 | 8.8 | 9.1 |
| Crabtree-positive | 66.6 | 66.2 | 66.4 | 9.9 | 9.2 | 9.6 | |
| Contrast 4 | P. kudriazevii KKU20 | 68.0 | 64.7 | 66.3 | 9.4 | 9.0 | 9.2 |
| C. tropicalis KKU20 | 64.7 | 65.0 | 64.9 | 9.2 | 8.6 | 8.9 | |
| Yeast Species | WBT Level | pH | Ammonia Nitrogen mg/dL | ||||
|---|---|---|---|---|---|---|---|
| 4 h | 8 h | Mean | 4 h | 8 h | Mean | ||
| Control | 100 | 7.12 | 7.00 | 7.06 | 14.71 d | 17.95 c | 16.33 c |
| WBT | 50 | 7.11 ab | 7.01 | 7.06 | 15.23 d | 18.70 bc | 16.96 c |
| 100 | 7.13 a | 7.00 | 7.06 | 16.06 cd | 18.85 bc | 17.46 b | |
| WBT-NO | 50 | 7.09 ab | 7.00 | 7.04 | 15.83 d | 20.04 abc | 16.86 c |
| 100 | 7.08 b | 7.00 | 7.06 | 18.95 abc | 21.29 abc | 20.64 ab | |
| WBT-P | 50 | 7.10 ab | 7.06 | 7.08 | 15.71 d | 22.17 ab | 20.81 ab |
| 100 | 7.07 b | 7.01 | 7.04 | 19.22 ab | 22.71 a | 20.96 ab | |
| WBT-C | 50 | 7.11 ab | 6.99 | 7.05 | 16.75 bcd | 20.53 abc | 18.64 abc |
| 100 | 7.08 ab | 7.00 | 7.04 | 20.59 a | 21.37 abc | 20.98 ab | |
| WBT-S | 50 | 7.08 ab | 6.99 | 7.04 | 17.05 bcd | 20.52 abc | 18.78 abc |
| 100 | 7.08 ab | 7.02 | 7.05 | 20.68 a | 22.47 ab | 21.50 a | |
| SEM | 0.01 | 0.01 | 0.01 | 0.76 | 0.91 | 0.99 | |
| Control vs. Others | 0.06 | 0.69 | 0.45 | p < 0.01 | 0.022 | 0.016 | |
| Interaction | p < 0.01 | 0.70 | 0.11 | p < 0.01 | 0.022 | 0.016 | |
| Comparison | |||||||
| Process | WBT | 7.12 | 7.00 | 7.06 | 15.65 | 18.77 | 17.21 |
| WBT-NO | 7.08 | 7.00 | 7.05 | 17.39 | 20.67 | 18.75 | |
| WBT-P | 7.08 | 7.03 | 7.06 | 17.46 | 22.44 | 20.89 | |
| WBT-C | 7.10 | 6.99 | 7.04 | 18.67 | 20.95 | 19.81 | |
| WBT-SC | 7.08 | 7.01 | 7.04 | 18.87 | 21.49 | 20.14 | |
| Level of WBT | 50 | 7.10 | 7.01 | 7.05 | 16.11 | 20.39 | 18.41 |
| 100 | 7.09 | 7.01 | 7.05 | 19.10 | 21.34 | 20.31 | |
| Contrast 1 | Unfermented | 7.12 a | 7.00 | 7.06 | 15.65 b | 18.77 b | 17.21 b |
| Fermented | 7.08 b | 7.01 | 7.05 | 18.10 a | 21.39 a | 19.90 a | |
| Contrast 2 | No yeast | 7.08 | 7.00 | 7.05 | 17.39 | 20.67 | 18.75 |
| With yeast | 7.09 | 7.01 | 7.05 | 18.33 | 21.63 | 20.28 | |
| Contrast 3 | Crabtree-negative | 7.09 | 7.01 | 7.05 | 18.07 | 21.70 | 20.35 |
| Crabtree-positive | 7.08 | 7.01 | 7.04 | 18.87 | 21.49 | 20.14 | |
| Contrast 4 | P. kudriazevii KKU20 | 7.08 | 7.03 a | 7.06 | 17.46 | 22.44 | 20.89 |
| C.tropicalis KKU20 | 7.10 | 6.99 b | 7.04 | 18.67 | 20.95 | 19.81 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suntara, C.; Wanapat, M.; Chankaew, S.; Khonkhaeng, B.; Supapong, C.; Chanjula, P.; Gunun, P.; Gunun, N.; Foiklang, S.; Phesatcha, K.; et al. Improvement of the Nutritional Quality of Psophocarpus tetragonolobus Tubers by Fermentation with Ruminal Crabtree-Negative Yeasts on the In Vitro Digestibility and Fermentation in Rumen Fluid. Fermentation 2022, 8, 209. https://doi.org/10.3390/fermentation8050209
Suntara C, Wanapat M, Chankaew S, Khonkhaeng B, Supapong C, Chanjula P, Gunun P, Gunun N, Foiklang S, Phesatcha K, et al. Improvement of the Nutritional Quality of Psophocarpus tetragonolobus Tubers by Fermentation with Ruminal Crabtree-Negative Yeasts on the In Vitro Digestibility and Fermentation in Rumen Fluid. Fermentation. 2022; 8(5):209. https://doi.org/10.3390/fermentation8050209
Chicago/Turabian StyleSuntara, Chanon, Metha Wanapat, Sompong Chankaew, Benjamad Khonkhaeng, Chanadol Supapong, Pin Chanjula, Pongsatorn Gunun, Nirawan Gunun, Suban Foiklang, Kampanat Phesatcha, and et al. 2022. "Improvement of the Nutritional Quality of Psophocarpus tetragonolobus Tubers by Fermentation with Ruminal Crabtree-Negative Yeasts on the In Vitro Digestibility and Fermentation in Rumen Fluid" Fermentation 8, no. 5: 209. https://doi.org/10.3390/fermentation8050209
APA StyleSuntara, C., Wanapat, M., Chankaew, S., Khonkhaeng, B., Supapong, C., Chanjula, P., Gunun, P., Gunun, N., Foiklang, S., Phesatcha, K., & Cherdthong, A. (2022). Improvement of the Nutritional Quality of Psophocarpus tetragonolobus Tubers by Fermentation with Ruminal Crabtree-Negative Yeasts on the In Vitro Digestibility and Fermentation in Rumen Fluid. Fermentation, 8(5), 209. https://doi.org/10.3390/fermentation8050209