
Designing a Waste-Based Culture Medium for the Production of Plant Growth Promoting Microorganisms Based on Cladodes Juice from Opuntia ficus-indica Pruning

 , ,
, ,  , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Plant Material and Juice Extraction
2.3. Determination of Yield, pH, Total Soluble Solids, Density and Dry-Weight Matter on Juice
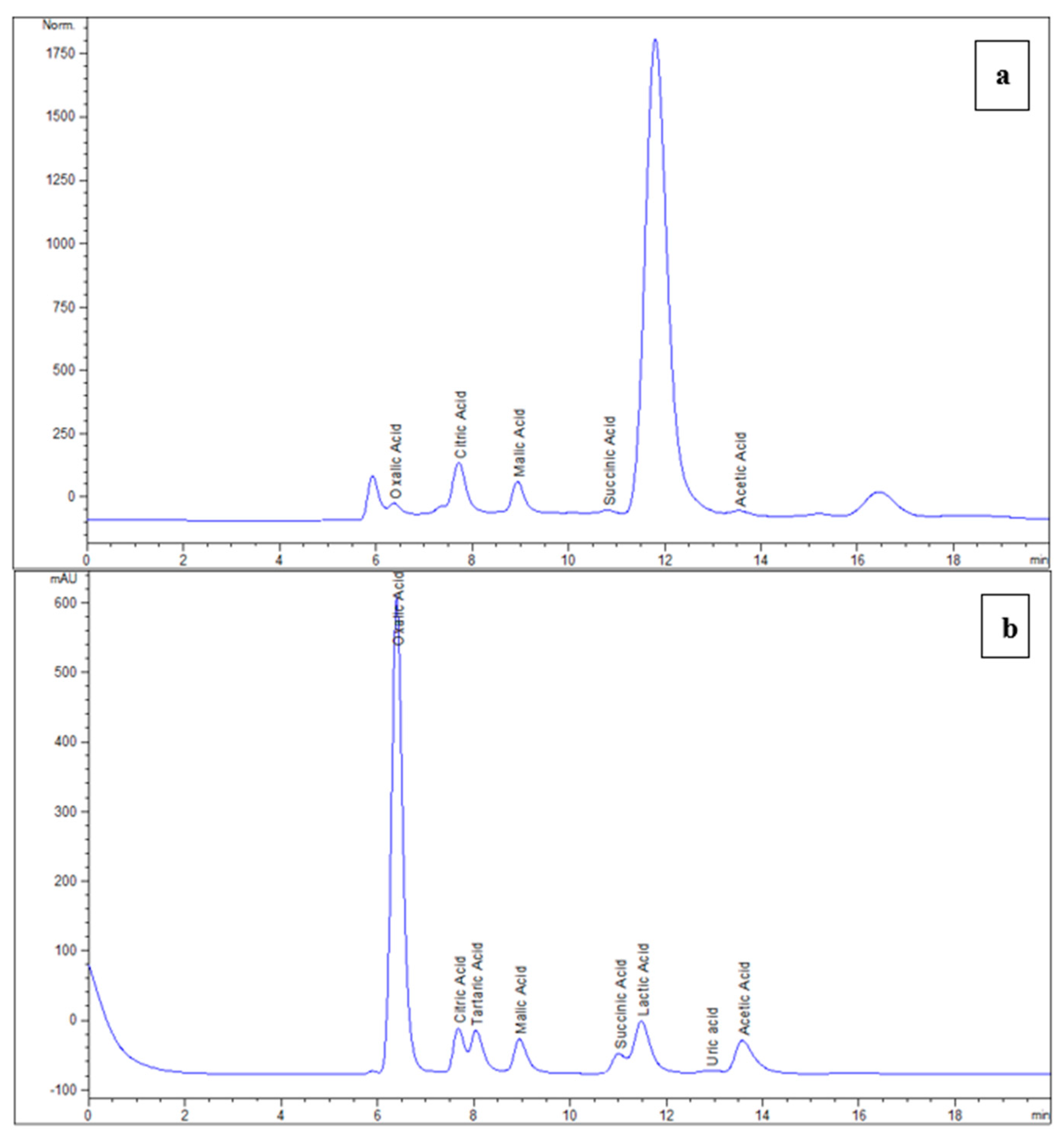
2.4. Determination of Sugars and Organic Acids in Juice
2.5. Shaken Flasks Cultivation of Five Microbial Strains on Different Dilution of Cladode Juice and Conventional Media
2.6. Production of Microbial Biomass in Bioreactor
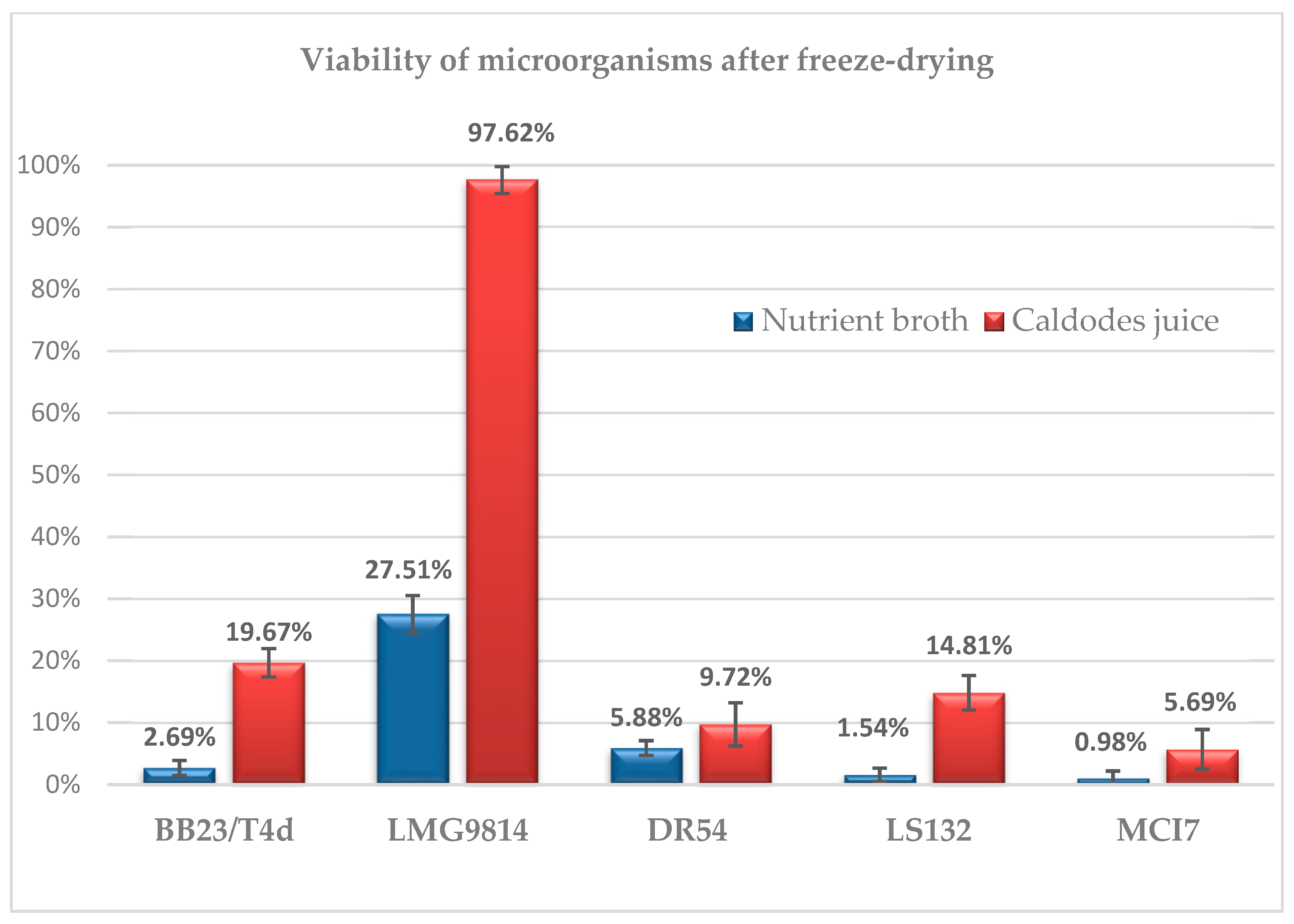
2.7. Drying Process of Microbial Biomass and Evaluation of Viable Cells
- (Ng) was the number of viable cells determined in 1 g of product after 24 h from the drying process;
- (Ne) was the number of theoretical viable cells in the product, obtained by dividing the number of CFU present in 1 mL of liquid culture by the final weight of the product (in this case, 5 g zeolite + 1 mL liquid culture).
- (N0) and (N) were the number of viable cells from the vials before and after lyophilization, respectively.
2.8. Statistical Analysis
3. Results and Discussion
3.1. Determination of Yield, pH, Total Soluble Solids, Density and Dry-Weight Matter on Juice
3.2. Sugars and Organic Acids Analyses of Cladodes Juice
3.3. Shaken Flasks Cultivation of Five Microbial Strains on Different Dilution of Cladodes Juice and Conventional Media
3.4. Production of Microbial Biomass in Bioreactor
3.5. Drying Processes of Microbial Biomass and Evaluation of Viable Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bijay-Singh; Craswell, E. Fertilizers and nitrate pollution of surface and ground water: An increasingly pervasive global problem. SN Appl. Sci. 2021, 3, 518. [Google Scholar] [CrossRef]
- Bisht, N.; Chauhan, P.S. Excessive and Disproportionate Use of Chemicals Cause Soil Contamination and Nutritional Stress, In Soil Contamination—Threats and Sustainable Solutions; Larramendy, M.L., Soloneski, S., Eds.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as biofertilizers, a potential approach for sustainable crop production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Raghuwanshi, R. Opportunities and challenges to sustainable agriculture in India. Nebio 2012, 3, 78–86. [Google Scholar]
- Kawalekar, J.S. Role of biofertilizers and biopesticides for sustainable agriculture. J. Bio. Innov. 2013, 2, 73–78. [Google Scholar]
- Tabacchioni, S.; Passato, S.; Ambrosino, P.; Huang, L.; Caldara, M.; Cantale, C.; Hett, J.; Del Fiore, A.; Fiore, A.; Schlüter, A.; et al. Identification of beneficial microbial consortia and bioactive compounds with potential as plant biostimulants for a sustainable agriculture. Microorganisms 2021, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, W.; Hou, J. Plant growth and soil microbial impacts of enhancing licorice with inoculating dark septate endophytes under drought stress. Front. Microbiol. 2019, 10, 2277. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, S.S.; Hassan, G.; Samoon, S.; Rather, H.; Dar, S.A.; Zehra, B. Bio-fertilizers in organic agriculture. J. Phytol. 2010, 2, 42–54. [Google Scholar]
- Vassilev, N.; Vassileva, M.; Lopez, A.; Martos, V.; Reyes, A.; Maksimovic, I.; Eichler-Löbermann, B.; Malusà, E. Unexploited potential of some biotechnological techniques for biofertilizer production and formulation. Appl. Microbiol. Biotechnol. 2015, 99, 4983–4996. [Google Scholar] [CrossRef]
- Vassilev, N.; Malusa, E.; Requena, A.R.; Martos, V.; López, A.; Maksimovic, I.; Vassileva, M. Potential application of glycerol in the production of plant beneficial microorganisms. J. Ind. Microbiol. Biotechnol. 2017, 44, 735–743. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid. Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Tzimorotas, D.; Afseth, N.K.; Lindberg, D.; Kjørlaug, O.; Axelsson, L.; Shapaval, V. Pretreatment of different food rest materials for bioconversion into fungal lipid-rich biomass. Bioprocess. Biosyst. Eng. 2018, 41, 1039–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, E.Z. Production of polyhydroxy alkanoates (PHAs) by Bacillus subtilis and Escherichia coli grown on cane molasses fortified with ethanol. Braz. Arch. Biol. Technol. 2014, 57, 145–154. [Google Scholar] [CrossRef]
- Tropea, A. Food waste valorization. Fermentation 2022, 8, 168. [Google Scholar] [CrossRef]
- Youssef, H.H.; Hamza, M.A.; Fayez, M.; Mourad, E.F.; Saleh, M.Y.; Sarhan, M.S.; Suker, R.M.; Eltahlawy, A.A.; Nemr, R.A.; El-Tahan, M.; et al. Plant-based culture media: Efficiently support culturing rhizobacteria and correctly mirror their in-situ diversity. J. Adv. Res. 2016, 7, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Nour, E.H.; Hamza, M.A.; Fayez, M.; Monib, M.; Ruppel, S.; Hegazi, N.A. The crude plant juices of desert plants as appropriate culture media for the cultivation of rhizospheric microorganisms. J. Adv. Res. 2012, 3, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Pham, V.H.; Kim, J. Cultivation of unculturable soil bacteria. Trends. Biotechnol. 2012, 30, 475–484. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Khulan, A.; Kim, J. Development of a novel cultivation technique for uncultured soil bacteria. Sci. Rep. 2019, 9, 6666. [Google Scholar] [CrossRef]
- Mourad, E.F.; Sarhan, M.S.; Daanaa, H.A.; Abdou, M.; Morsi, A.T.; Abdelfadeel, M.R.; Elsawey, H.; Nemr, R.; El-Tahan, M.; Hamza, M.A.; et al. Plant materials are sustainable substrates supporting new technologies of plant-only-based culture media for in vitro culturing of the plant microbiota. Microbes Environ. 2018, 33, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Kuloyo, O.O.; du Preez, J.C.; García-Aparicio, M.P.; Kilian, S.G.; Steyn, L.; Görgens, J. Opuntia ficus-indica cladodes as feedstock for ethanol production by Kluyveromyces marxianus and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2014, 30, 3173–3183. [Google Scholar] [CrossRef] [Green Version]
- Haddish, K. Production of single cell protein from fruit of beles (Opuntia ficus-indica) peels using Saccharomyces cerevisiae. J. Microbiol. Exp. 2015, 2, 00073. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, T.G.; Pereira, P.; Silva, M.A.; Vicente, F.; Ramalho, R.; Costa, H.S. Prickly pear. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Dublin, Ireland, 2020; pp. 709–728. [Google Scholar] [CrossRef]
- Derabli, B.; Nancib, A.; Nancib, N.; Aníbal, J.; Raposo, S.; Rodrigues, B.; Boudrant, J. Opuntia ficusindica waste as a cost effective carbon source for lactic acid production by Lactobacillus plantarum. Food Chem. 2022, 370, 131005. [Google Scholar] [CrossRef] [PubMed]
- Blair, B.B.; Yim, W.C.; Cushman, J.C. Characterization of a microbial consortium with potential for biological degradation of cactus pear biomass for biofuel production. Heliyon 2021, 7, e07854. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Agüero, J.; Aguirre-Rivera, J.; Hernández, H. Systematic notes and a detailed description of Opuntia ficus-indica (L.) Mill. (Cactaceae). Agrociencia 2005, 39, 395–408. [Google Scholar]
- Silva, M.A.; Albuquerque, T.G.; Pereira, P.; Ramalho, R.; Vicente, F.; Oliveira, M.B.P.P.; Costa, H.S. Opuntia ficus-indica (L.) Mill. A multi-benefit potential to be exploited. Molecules 2021, 26, 951. [Google Scholar] [CrossRef] [PubMed]
- Perucini-Avendaño, M.; Nicolás-García, M.; Jiménez-Martínez, C.; Perea-Flores, M.J.; Gómez-Patiño, M.B.; Arrieta-Báez, D.; Dávila-Ortiz, G. Cladodes: Chemical and structural properties, biological activity, and polyphenols profile. Food Sci. Nutr. 2021, 9, 4007–4017. [Google Scholar] [CrossRef]
- Ginestra, G.; Parker, M.L.; Bennett, R.N.; Robertson, J.; Mandalari, G.; Narbad, A.; Lo Curto, R.B.; Bisignano, G.; Faulds, C.B.; Waldron, K.W. Anatomical, chemical, and biochemical characterization of cladodes from prickly pear [Opuntia ficus-Indica (L.) Mill.]. J. Agric. Food Chem. 2009, 57, 10323–10330. [Google Scholar] [CrossRef]
- Andreu, L.; Nuncio-Jáuregui, N.; Carbonell-Barrachina, Á.A.; Legua, P.; Hernández, F. Antioxidant properties and chemical characterization of Spanish Opuntia ficus-indica Mill. cladodes and fruits. J. Sci. Food Agric. 2018, 98, 1566–1573. [Google Scholar] [CrossRef]
- Yang, L.; Lu, M.; Carl, S.; Mayer, J.A.; Cushman, J.C.; Tian, E.; Lin, H. Biomass characterization of Agave and Opuntia as poten-tial biofuel feedstocks. Biomass Bioenergy 2015, 76, 43–53. [Google Scholar] [CrossRef]
- Inglese, P.; Basile, F.; Schirra, M. Cactus pear fruit production. In Cacti: Biology and Uses; Nobel, P.S., Ed.; University of California Press: Berkeley, CA, USA, 2002; pp. 163–184. [Google Scholar]
- Inglese, P. Cactus pear, Opuntia ficus-indica L. (Mill.) for fruit production: An overview. In Proceedings of the Improved utilization of cactus pear for food, feed, soil and water conservation and other products in Africa, Mekelle, Ethiopia, 19–21 October 2009. [Google Scholar]
- Procacci, S.; Bojorquez-Quintal, E.; Platamone, G.; Oliviero, M.; Lo Vecchio, V.; Morreale, V.; Alisi, C.; Balducchi, R.; Bacchetta, L.P. Opuntia ficus-indica pruning waste recycling: Recovery and characterization of mucilage from cladodes. Nat. Resour. 2021, 12, 91–107. [Google Scholar] [CrossRef]
- Hett, J.; Neuhoff, D.; Döring, T.F.; Masoero, G.; Ercole, E.; Bevivino, A. Effects of multi-species microbial inoculants on early wheat growth and litterbag microbial activity. Agronomy 2022, 12, 899. [Google Scholar] [CrossRef]
- Boutakiout, A.; Elothmani, D.; Hanine, H.; Mahrouz, M.; Le Meurlay, D.; Hmid, I.; Ennahli, S. Effects of different harvesting seasons on antioxidant activity and phenolic content of prickly pear cladode juice. J. Saudi Soc. Agric. Sci. 2018, 17, 471–480. [Google Scholar] [CrossRef]
- Betancourt-Domínguez, M.A.; Hernández-Pérez, T.; García-Saucedo, P.; Cruz-Hernández, A.; Paredes-López, O. Physico-chemical changes in cladodes (Nopalitos) from cultivated and wild cacti (Opuntia spp.). Plant. Foods Hum. Nutr 2006, 61, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Luskar, L.; Polanšek, J.; Hladnik, A.; Čeh, B. On-Farm Composting of Hop Plant Green Waste—Chemical and Biological Value of Compost. Appl. Sci. 2022, 12, 4190. [Google Scholar] [CrossRef]
- Vassileva, M.; Malusà, E.; Sas-Paszt, L.; Trzcinski, P.; Galvez, A.; Flor-Peregrin, E.; Shilev, S.; Canfora, L.; Mocali, S.; Vassilev, N. Fermentation strategies to improve soil bio-inoculant production and quality. Microorganisms 2021, 9, 1254. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, G.; Gautam, D.; Bedi, M.K. Optimization of dairy sludge for growth of Rhizobium cells. Biomed. Res. Int. 2013, 2013, e845264. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Bai, Z.; Jin, B.; Xiao, R.; Zhuang, G. Bioconversion of wastewater from sweet potato starch production to Paenibacillus polymyxa biofertilizer for tea plants. Sci. Rep. 2014, 4, e4131. [Google Scholar] [CrossRef]
- Romano, I.; Ventorino, V.; Ambrosino, P.; Testa, A.; Chouyia, F.E.; Pepe, O. Development and application of low-cost and eco-sustainable bio-stimulant containing a new plant growth-promoting strain Kosakonia pseudosacchari TL13. Front. Microbiol. 2020, 11, 2044. [Google Scholar] [CrossRef]
- Tsai, W.-T.; Lin, Y.-Q. Analysis of promotion policies for the valorization of food waste from industrial sources in Tai wan. Fermentation 2021, 7, 51. [Google Scholar] [CrossRef]
- Wolny-Koładka, K.; Malina, D.; Suder, A.; Pluta, K.; Wzorek, Z. Bio-based synthesis of silver nanoparticles from waste agricultural biomass and its antimicrobial activity. Processes 2022, 10, 389. [Google Scholar] [CrossRef]
- Lopes, F.C.; Ligabue-Braun, R. Agro-industrial residues: Eco-friendly and inexpensive substrates for microbial pigments production. Front. Sustain. Food Syst. 2021, 5, 589414. [Google Scholar] [CrossRef]
- Madeira, J.V., Jr.; Contesini, F.J.; Calzado, F.; Rubio, M.V.; Zubieta, M.P.; Lopes, D.B.; de Melo, R.R. Agro-industrial residues and microbial enzymes: An overview on the eco-friendly bioconversion into high value-added products. In Biotechnology of Microbial Enzymes; Brahmachari, G., Ed.; Elsevier: London, UK, 2017; pp. 475–511. [Google Scholar] [CrossRef]
- Cantabella, D.; Dolcet-Sanjuan, R.; Solsona, C.; Vilanova, L.; Torres, R.; Teixidó, N. Optimization of a food industry-waste-based medium for the production of the plant growth promoting microorganism Pseudomonas oryzihabitans PGP01 based on agro-food industries by-products. Biotechnol. Rep. 2021, 32, e00675. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Bueis, R.; Mulas, R.; Gómez, X.; González-Andrés, F. Innovative liquid formulation of digestates for producing a biofertilizer based on Bacillus siamensis: Field testing on sweet pepper. J. Plant. Nutr. Soil. Sci. 2017, 180, 748–758. [Google Scholar] [CrossRef]
- Damir, O.; Mladen, P.; Bozidar, S.; Srdan, N. Cultivation of the bacterium Azotobacter chroococcum for the preparation of biofertilizers. Afr. J. Biotechnol. 2011, 10, 3104–3111. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, H.; Bashir, H.; Nawaz, A.; Haq, I. Optimization of growth conditions for Azotobacter species and their use as biofertilizer. J. Bacteriol. Mycol. 2018, 6, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yu, Y.; Wang, Y.; Wei, X.; Liao, M.; Rong, X.; Chen, J. Development of new protocol for freeze-drying preservation of Pseudoalteromonas nigrifaciens and its protective effect on other marine bacteria. Electron. J. Biotechnol. 2020, 44, 1–5. [Google Scholar] [CrossRef]
- Haruta, S.; Kanno, N. Survivability of microbes in natural environments and their ecological impacts. Microbes Environ. 2015, 30, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Dam, S.; Pagès, J.-M.; Masi, M. Stress responses, outer membrane permeability control and antimicrobial resistance in Enterobacteriaceae. Microbiology 2018, 164, 260–267. [Google Scholar] [CrossRef]
- Gregory, G.J.; Boyd, E.F. Stressed out: Bacterial response to high salinity using compatible solute biosynthesis and uptake systems, lessons from Vibrionaceae. Comp. Struct. Biotechnol. J. 2021, 19, 1014–1027. [Google Scholar] [CrossRef]
- Hallsworth, J.E. Stress-free microbes lack vitality. Fungal Biol. 2018, 122, 379–385. [Google Scholar] [CrossRef]
- Berninger, T.; González López, Ó.; Bejarano, A.; Preininger, C.; Sessitsch, A. Maintenance and assessment of cell viability in formulation of non-sporulating bacterial inoculants. Microb. Biotechnol. 2018, 11, 277–301. [Google Scholar] [CrossRef] [Green Version]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant. Sci. 2015, 2, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Value ± SD | |
|---|---|
| Yield (%) | 20.2 ± 1.20 |
| pH | 4.3 ± 0.1 |
| Density (g·cm−3) | 1.03 ± 0.01 |
| TSS as °Brix | 5.9 ± 0.2 |
| Dry mass (% fw) | 6.83 ± 0.3 |
| RT (min) | Detector | Concentration (g L−1) Mean ± SD | |
|---|---|---|---|
| Sugars | |||
| Maltose | 7.405 | RID | 1.08 ± 0.28 |
| Sucrose | 7.495 | RID | 2.38 ± 0.58 |
| Glucose | 8.555 | RID | 8.82 ± 0.60 |
| Fructose | 9.285 | RID | 5.79 ± 1.28 |
| Arabinose | 9.785 | RID | traces |
| Organic Acids | |||
| Oxalic acid | 6.362 | UV 210 nm | 0.13 ± 0.2 |
| Citric acid | 7.763 | UV 210 nm | 8.90 ± 0.93 |
| Tartaric acid | 7.951 | UV 210 nm | n.d |
| Malic acid | 8.990 | UV 210 nm | 5.80 ± 0.59 |
| Succinic acid | 10.988 | UV 210 nm | 1.26 ± 0.16 |
| Acetic acid | 13.79 | UV 210 nm | traces |
| Bacterial Strains | Nutrient Broth | Cladodes Juice Concentration | ||
|---|---|---|---|---|
| 15 (%) | 25 (%) | 50 (%) | ||
| A. chrococcum LS 132 | 9.26 ± 0.08 a * | 8.86 ± 0.05 b | 9.24 ± 0.02 a | 9.32 ± 0.04 a |
| B. ambifaria MCI 7 | 9.45 ± 0.15 a | 9.04 ± 0.03 b | 9.28 ± 0.04 a | 9.4 ± 0.05 a |
| B. amyloliquefaciens LMG 9814 | 8.26 ± 0.04 a | 7.85 ± 0.11 b | 8.2± 0.01 a | 8.3 ± 0.01 a |
| P. fluorescens DR54 | 9.4 ± 0.06 a | 8.69 ± 0.01 b | 8.86 ± 0.07 b | 9.2 ± 0.02 a |
| R. aquatilis BB23/T4d | 9.18 ± 0.06 a | 8.61 ± 0.08 b | 9.23 ± 0.05 a | 9.43 ± 0.02 a |
| Bacterial Strains | Nutrient Broth (Log CFU mL−1) | Cladodes Juice 15% + Sucrose 0.2% (Log CFU mL−1) |
|---|---|---|
| A. chroococcum LS132 | 9.12 ± 0.01 a * | 9.03 ± 0.01 a |
| B. ambifaria MCI7 | 9.08 ± 0.03 a | 8.98 ± 0.04 a |
| B. amyloliquefaciens LMG 9814 | 8.08 ± 0.05 a | 7.91 ± 0.06 a |
| P. fluorescens DR54 | 9.00 ± 0.02 a | 8.96 ± 0.03 a |
| R. aquatilis BB23/T4d | 9.35 ± 0.01 a | 9.08 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magarelli, R.A.; Trupo, M.; Ambrico, A.; Larocca, V.; Martino, M.; Palazzo, S.; Balducchi, R.; Joutsjoki, V.; Pihlanto, A.; Bevivino, A. Designing a Waste-Based Culture Medium for the Production of Plant Growth Promoting Microorganisms Based on Cladodes Juice from Opuntia ficus-indica Pruning. Fermentation 2022, 8, 225. https://doi.org/10.3390/fermentation8050225
Magarelli RA, Trupo M, Ambrico A, Larocca V, Martino M, Palazzo S, Balducchi R, Joutsjoki V, Pihlanto A, Bevivino A. Designing a Waste-Based Culture Medium for the Production of Plant Growth Promoting Microorganisms Based on Cladodes Juice from Opuntia ficus-indica Pruning. Fermentation. 2022; 8(5):225. https://doi.org/10.3390/fermentation8050225
Chicago/Turabian StyleMagarelli, Rosaria Alessandra, Mario Trupo, Alfredo Ambrico, Vincenzo Larocca, Maria Martino, Salvatore Palazzo, Roberto Balducchi, Vesa Joutsjoki, Anne Pihlanto, and Annamaria Bevivino. 2022. "Designing a Waste-Based Culture Medium for the Production of Plant Growth Promoting Microorganisms Based on Cladodes Juice from Opuntia ficus-indica Pruning" Fermentation 8, no. 5: 225. https://doi.org/10.3390/fermentation8050225
APA StyleMagarelli, R. A., Trupo, M., Ambrico, A., Larocca, V., Martino, M., Palazzo, S., Balducchi, R., Joutsjoki, V., Pihlanto, A., & Bevivino, A. (2022). Designing a Waste-Based Culture Medium for the Production of Plant Growth Promoting Microorganisms Based on Cladodes Juice from Opuntia ficus-indica Pruning. Fermentation, 8(5), 225. https://doi.org/10.3390/fermentation8050225