
The Influence of Sugar Composition and pH Regulation in Batch and Continuous Acetone–Butanol–Ethanol Fermentation

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Medium Fermentation
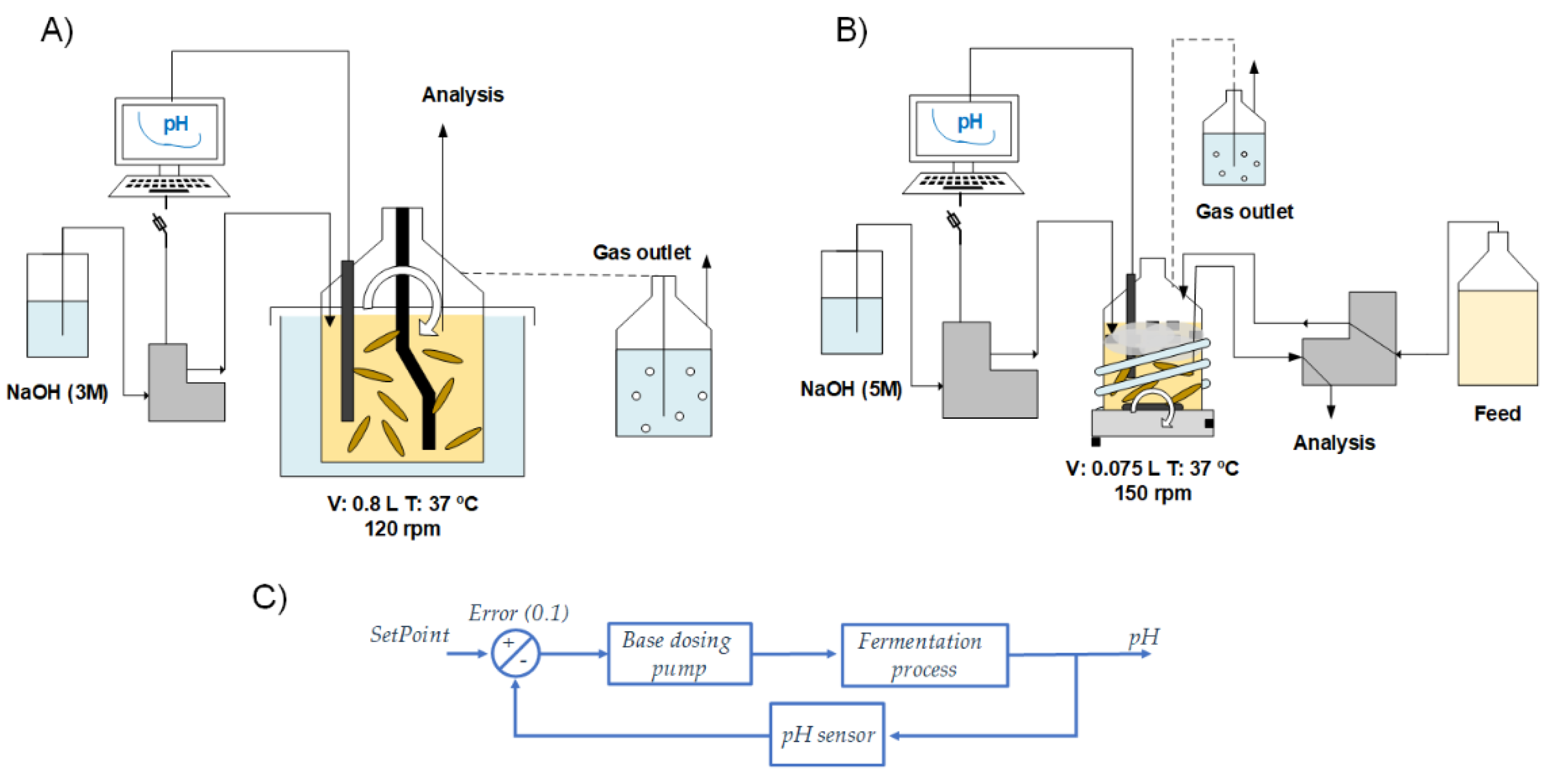
2.2. Experimental Setup
2.2.1. Batch Fermentations
2.2.2. Continuous Fermentation
2.3. Analytical Methods
3. Results and Discussion
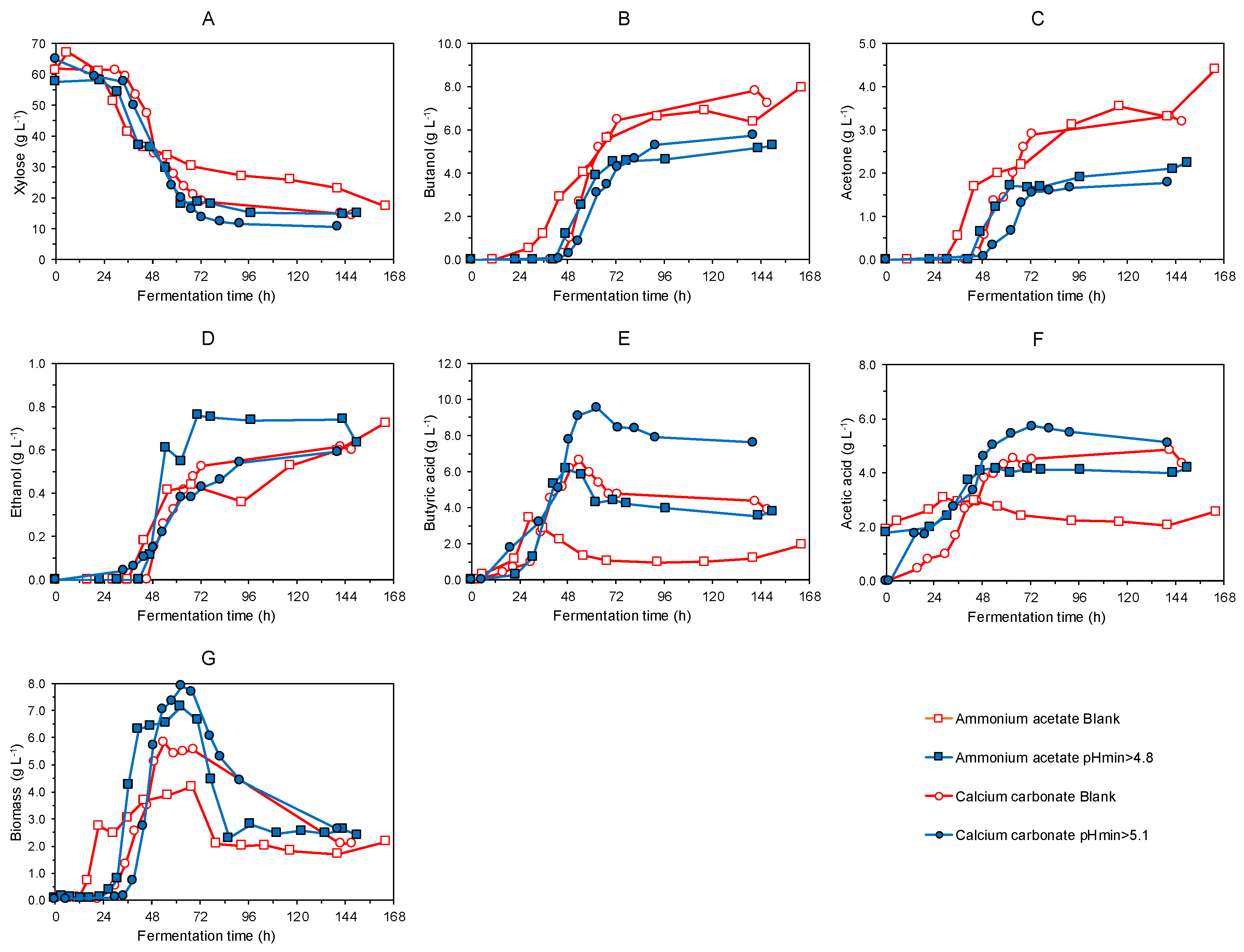
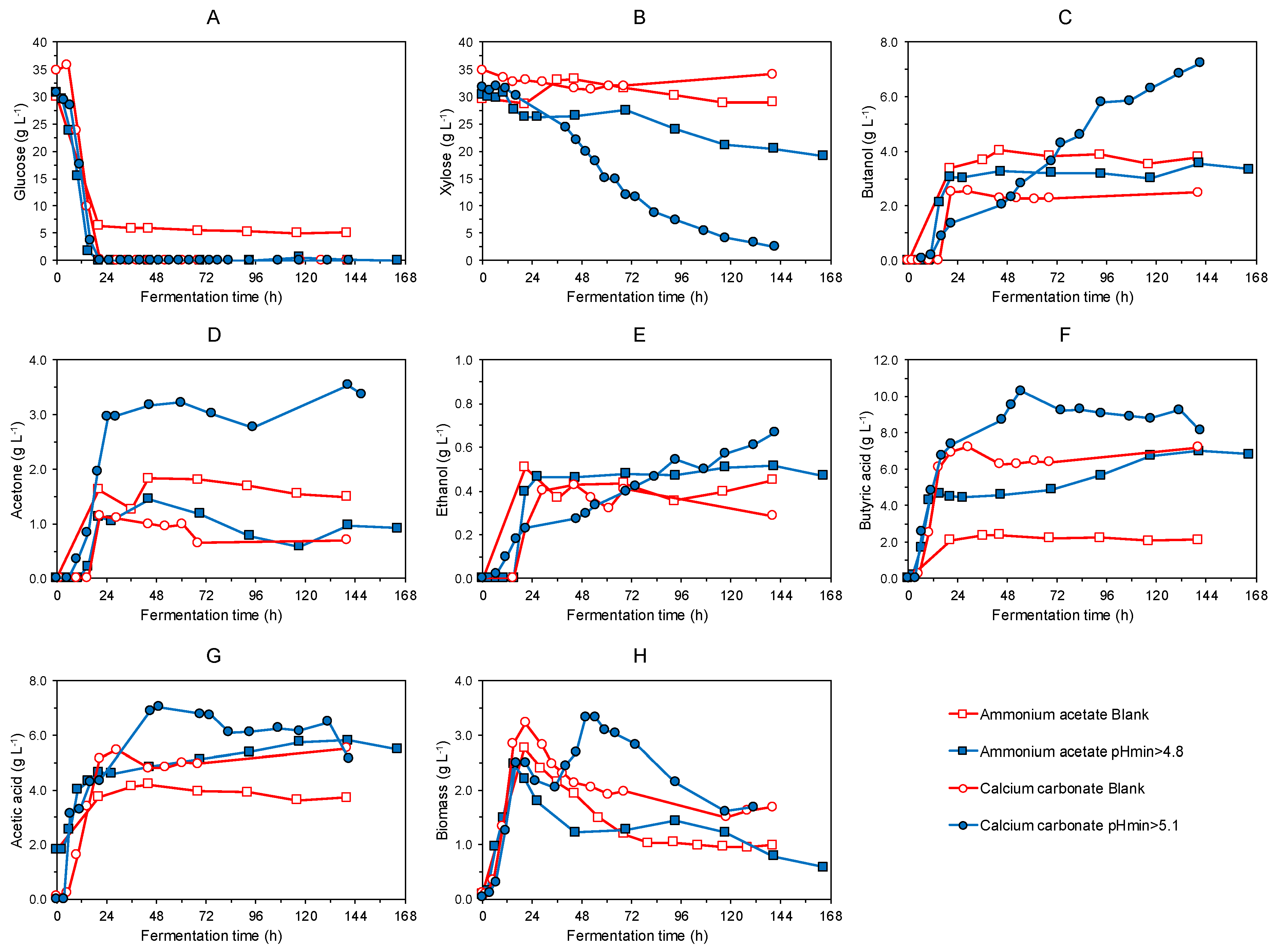
3.1. Batch Fermentation
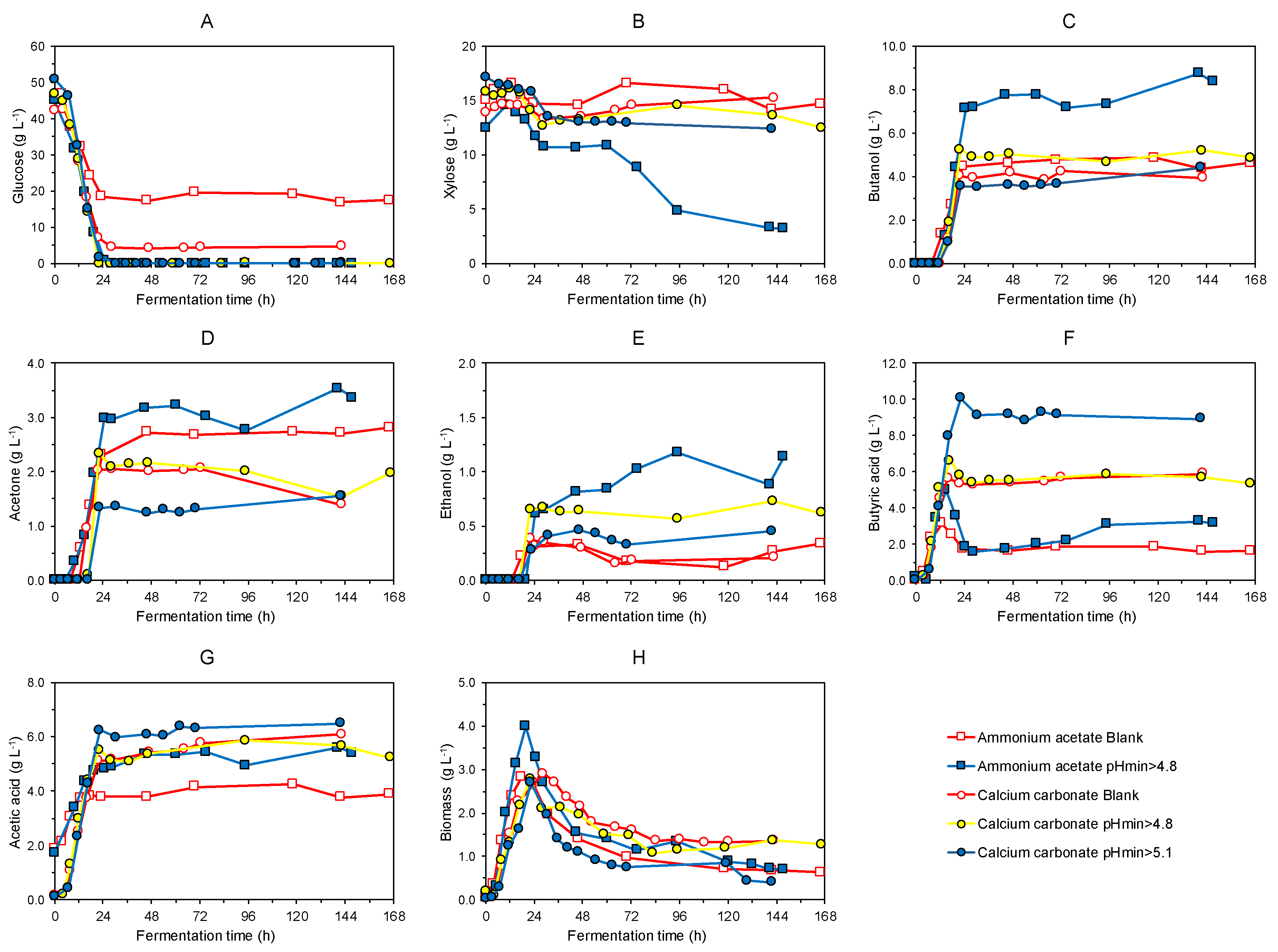
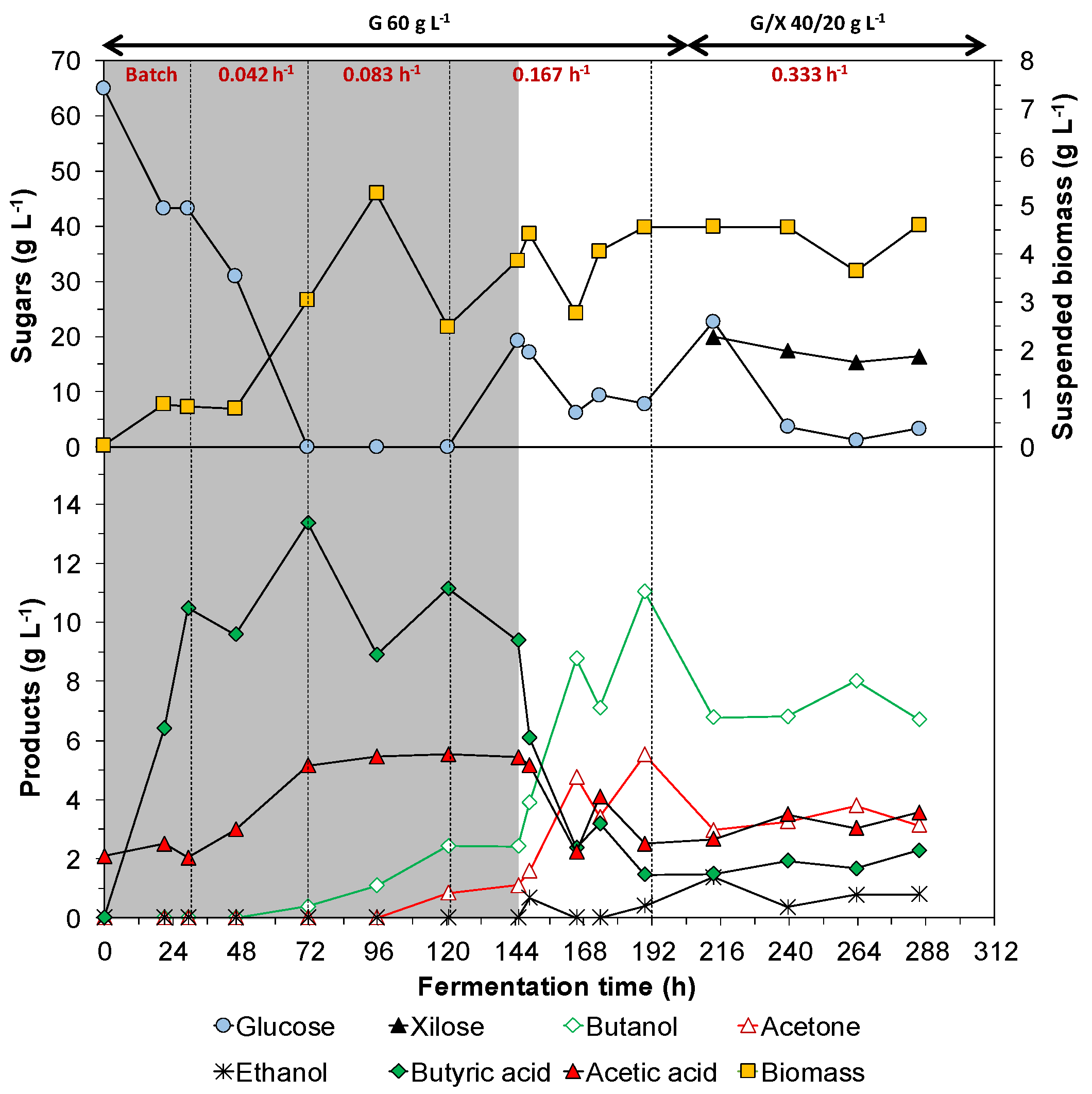
3.2. Continous Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bharathiraja, B.; Jayamuthunagai, J.; Sudharsanaa, T.; Bharghavi, A.; Praveenkumar, R.; Chakravarthy, M.; Devarajan, Y. Biobutanol—An Impending Biofuel for Future: A Review on Upstream and Downstream Processing Tecniques. Renew. Sustain. Energy Rev. 2017, 68, 788–807. [Google Scholar] [CrossRef]
- Dürre, P. Biobutanol: An Attractive Biofuel. Biotechnol. J. 2007, 2, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.F.; Ramli, N.; Kamal Bahrin, E.; Abd-Aziz, S. Cellulosic Biobutanol by Clostridia: Challenges and Improvements. Renew. Sustain. Energy Rev. 2017, 79, 1241–1254. [Google Scholar] [CrossRef]
- Narisetty, V.; Cox, R.; Bommareddy, R.; Agrawal, D.; Ahmad, E.; Pant, K.K.; Chandel, A.K.; Bhatia, S.K.; Kumar, D.; Binod, P.; et al. Valorisation of Xylose to Renewable Fuels and Chemicals, an Essential Step in Augmenting the Commercial Viability of Lignocellulosic Biorefineries. Sustain. Energy Fuels 2022, 6, 29–65. [Google Scholar] [CrossRef]
- Ren, C.; Gu, Y.; Hu, S.; Wu, Y.; Wang, P.; Yang, Y.; Yang, C.; Yang, S.; Jiang, W. Identification and Inactivation of Pleiotropic Regulator CcpA to Eliminate Glucose Repression of Xylose Utilization in Clostridium Acetobutylicum. Metab. Eng. 2010, 12, 446–454. [Google Scholar] [CrossRef]
- Birgen, C.; Dürre, P.; Preisig, H.A.; Wentzel, A. Butanol Production from Lignocellulosic Biomass: Revisiting Fermentation Performance Indicators with Exploratory Data Analysis. Biotechnol. Biofuels 2019, 12, 167. [Google Scholar] [CrossRef]
- Buendia-Kandia, F.; Rondags, E.; Framboisier, X.; Mauviel, G.; Dufour, A.; Guedon, E. Diauxic Growth of Clostridium Acetobutylicum ATCC 824 When Grown on Mixtures of Glucose and Cellobiose. AMB Express 2018, 8, 85. [Google Scholar] [CrossRef]
- Díez-antolínez, R.; Hijosa-valsero, M.; Paniagua-garcía, A.I. Effect of Nutrient Supplementation on Biobutanol Production from Cheese Whey by ABE (Acetone–Butanol–Ethanol) Fermentation. Chem. Eng. Trans. 2016, 49, 217–222. [Google Scholar] [CrossRef]
- Qureshi, N.; Blaschek, H.P. Production of Acetone Butanol Ethanol (ABE) by a Hyper-Producing Mutant Strain of Clostridium Beijerinckii BA101 and Recovery by Pervaporation. Biotechnol. Prog. 1999, 15, 594–602. [Google Scholar] [CrossRef]
- Monot, F.; Martin, J.R.; Petitdemange, H.; Gay, R. Acetone and Butanol Production by Clostridium Acetobutylicum in a Synthetic Medium. Appl. Environ. Microbiol. 1982, 44, 1318–1324. [Google Scholar] [CrossRef]
- Raganati, F.; Olivieri, G.; Götz, P.; Marzocchella, A.; Salatino, P. Butanol Production from Hexoses and Pentoses by Fermentation of Clostridium Acetobutylicum. Anaerobe 2015, 34, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Valles, A.; Álvarez-Hornos, F.J.; Martínez-Soria, V.; Marzal, P.; Gabaldón, C. Comparison of Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation Processes for Butanol Production from Rice Straw. Fuel 2020, 282, 118831. [Google Scholar] [CrossRef]
- Jiang, W.; Wen, Z.; Wu, M.; Li, H.; Yang, J.; Lin, J.; Lin, Y.; Yang, L.; Cen, P. The Effect of PH Control on Acetone-Butanol-Ethanol Fermentation by Clostridium Acetobutylicum ATCC 824 with Xylose and D-Glucose and D-Xylose Mixture. Chin. J. Chem. Eng. 2014, 22, 937–942. [Google Scholar] [CrossRef]
- Maddox, I.S.; Steiner, E.; Hirsch, S.; Wessner, S.; Gutierrez, N.A.; Gapes, J.R.; Schuster, K.C. The Cause of “Acid Crash” and “Acidogenic Fermentations” during the Batch Acetone-Butanol-Ethanol (ABE-) Fermentation Process. J. Mol. Microbiol. Biotechnol. 2000, 2, 95–100. [Google Scholar]
- Yang, X.; Tu, M.; Xie, R.; Adhikari, S.; Tong, Z. A Comparison of Three PH Control Methods for Revealing Effects of Undissociated Butyric Acid on Specific Butanol Production Rate in Batch Fermentation of Clostridium Acetobutylicum. AMB Express 2013, 3, 3. [Google Scholar] [CrossRef]
- Ezeji, T.C.; Qureshi, N.; Blaschek, H.P. Acetone Butanol Ethanol (ABE) Production from Concentrated Substrate: Reduction in Substrate Inhibition by Fed-Batch Technique and Product Inhibition by Gas Stripping. Appl. Microbiol. Biotechnol. 2004, 63, 653–658. [Google Scholar] [CrossRef]
- Capilla, M.; San-Valero, P.; Izquierdo, M.; Penya-roja, J.M.; Gabaldón, C. The Combined Effect on Initial Glucose Concentration and PH Control Strategies for Acetone-Butanol-Ethanol (ABE) Fermentation by Clostridium Acetobutylicum DSM 792. Biochem. Eng. J. 2021, 167, 107910. [Google Scholar] [CrossRef]
- Guo, T.; Sun, B.; Jiang, M.; Wu, H.; Du, T.; Tang, Y.; Wei, P.; Ouyang, P. Enhancement of Butanol Production and Reducing Power Using a Two-Stage Controlled-PH Strategy in Batch Culture of Clostridium Acetobutylicum XY16. World J. Microbiol. Biotechnol. 2012, 28, 2551–2558. [Google Scholar] [CrossRef]
- Luo, H.; Zheng, P.; Xie, F.; Yang, R.; Liu, L.; Han, S.; Zhao, Y.; Bilal, M. Co-Production of Solvents and Organic Acids in Butanol Fermentation by: Clostridium Acetobutylicum in the Presence of Lignin-Derived Phenolics. RSC Adv. 2019, 9, 6919–6927. [Google Scholar] [CrossRef]
- Vees, C.A.; Neuendorf, C.S.; Pflügl, S. Towards Continuous Industrial Bioprocessing with Solventogenic and Acetogenic Clostridia: Challenges, Progress and Perspectives. J. Ind. Microbiol. Biotechnol. 2020, 47, 753–787. [Google Scholar] [CrossRef]
- Dolejš, I.; Krasňan, V.; Stloukal, R.; Rosenberg, M.; Rebroš, M. Butanol Production by Immobilised Clostridium Acetobutylicum in Repeated Batch, Fed-Batch, and Continuous Modes of Fermentation. Bioresour. Technol. 2014, 169, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, Y.; Ding, F.Y.; Zhao, T.; Wu, J.L.; Guo, T.; Ren, H.F.; Li, B.B.; Niu, H.Q.; Cao, Z.; et al. Biobutanol Production in a Clostridium Acetobutylicum Biofilm Reactor Integrated with Simultaneous Product Recovery by Adsorption. Biotechnol. Biofuels 2014, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Ounine, K.; Petitdemange, H.; Raval, G.; Gay, R. Regulation and Butanol Inhibition of D-Xylose and D-Glucose Uptake in Clostridium Acetobutylicum. Appl. Environ. Microbiol. 1985, 49, 874–878. [Google Scholar] [CrossRef] [PubMed]
- El Kanouni, A.; Zerdani, I.; Zaafa, S.; Znassni, M.; Loutfi, M.; Boudouma, M. The Improvement of Glucose/Xylose Fermentation by Clostridium Acetobutylicum Using Calcium Carbonate. World J. Microbiol. Biotechnol. 1998, 14, 431–435. [Google Scholar] [CrossRef]
- Darkwah, K.; Nokes, S.E.; Seay, J.R.; Knutson, B.L. Mechanistic Simulation of Batch Acetone–Butanol–Ethanol (ABE) Fermentation with in Situ Gas Stripping Using Aspen PlusTM. Bioprocess Biosyst. Eng. 2018, 41, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Bankar, S.B.; Survase, S.A.; Singhal, R.S.; Granström, T. Continuous Two Stage Acetone-Butanol-Ethanol Fermentation with Integrated Solvent Removal Using Clostridium Acetobutylicum B 5313. Bioresour. Technol. 2012, 106, 110–116. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucose/Xylose Ratio | Ammonium Acetate (2.2 g L−1) | Calcium Carbonate (5 g L−1) | ||
|---|---|---|---|---|
| Blank | pH Control | Blank | pH Control | |
| 0:1 | Yes | 4.8 | Yes | 5.1 |
| 1:1 | Yes | 4.8 | Yes | 5.1 |
| 3:1 | Yes | 4.8 | Yes | 4.8 and 5.1 |
| Time (h) | Minimum pH Set Point | Substrate | Concentration (g L−1) | D (h−1) |
|---|---|---|---|---|
| 0–30 | 6.0 | Glucose | 60 | Batch |
| 30–72 | 6.0 | Glucose | 60 | 0.042 |
| 72–121 | 6.0 | Glucose | 60 | 0.083 |
| 121–144 | 6.0 | Glucose | 60 | 0.167 |
| 144–192 | 4.8 | Glucose | 60 | 0.167 |
| 192–216 | 4.8 | Glucose | 60 | 0.333 |
| 216–288 | 4.8 | Glucose/Xylose | 40:20 | 0.333 |
| Time (h) | Substrate (g L−1) | pH | D (h−1) | g C g C of Monosaccharide Consumed−1 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Acetic Acid | Butyric Acid | Butanol | Acetone | Ethanol | Suspended Biomass | Yield | Unaccounted | ||||
| 0–30 | Glucose (60) | 6.0 | Batch | 0.00 | 0.66 | 0.00 | 0.00 | 0.00 | 0.05 | 0.70 | 0.30 |
| 30–72 | Glucose (60) | 6.0 | 0.042 | 0.14 | 0.39 | 0.01 | 0.00 | 0.00 | 0.05 | 0.59 | 0.41 |
| 72–121 | Glucose (60) | 6.0 | 0.083 | 0.09 | 0.21 | 0.06 | 0.02 | 0.00 | 0.09 | 0.47 | 0.53 |
| 121–144 | Glucose (60) | 6.0 | 0.167 | 0.13 | 0.30 | 0.10 | 0.04 | 0.00 | 0.13 | 0.70 | 0.30 |
| 144–192 | Glucose (60) | 4.8 | 0.167 | 0.07 | 0.05 | 0.28 | 0.13 | 0.01 | 0.11 | 0.65 | 0.35 |
| 192–216 | Glucose (60) | 4.8 | 0.333 | 0.07 | 0.05 | 0.27 | 0.11 | 0.05 | 0.16 | 0.72 | 0.28 |
| 216–288 | Glucose/Xylose (40:20) | 4.8 | 0.333 | 0.08 | 0.07 | 0.28 | 0.13 | 0.02 | 0.14 | 0.72 | 0.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capilla, M.; Silvestre, C.; Valles, A.; Álvarez-Hornos, F.J.; San-Valero, P.; Gabaldón, C. The Influence of Sugar Composition and pH Regulation in Batch and Continuous Acetone–Butanol–Ethanol Fermentation. Fermentation 2022, 8, 226. https://doi.org/10.3390/fermentation8050226
Capilla M, Silvestre C, Valles A, Álvarez-Hornos FJ, San-Valero P, Gabaldón C. The Influence of Sugar Composition and pH Regulation in Batch and Continuous Acetone–Butanol–Ethanol Fermentation. Fermentation. 2022; 8(5):226. https://doi.org/10.3390/fermentation8050226
Chicago/Turabian StyleCapilla, Miguel, Carlos Silvestre, Alejo Valles, Francisco Javier Álvarez-Hornos, Pau San-Valero, and Carmen Gabaldón. 2022. "The Influence of Sugar Composition and pH Regulation in Batch and Continuous Acetone–Butanol–Ethanol Fermentation" Fermentation 8, no. 5: 226. https://doi.org/10.3390/fermentation8050226
APA StyleCapilla, M., Silvestre, C., Valles, A., Álvarez-Hornos, F. J., San-Valero, P., & Gabaldón, C. (2022). The Influence of Sugar Composition and pH Regulation in Batch and Continuous Acetone–Butanol–Ethanol Fermentation. Fermentation, 8(5), 226. https://doi.org/10.3390/fermentation8050226