
Effects of Water Potential on Anaerobic Methane Production and a Microbial Consortium


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Water Potential (Ψ) Analysis
2.3. Methane Production Potential
2.3.1. Theoretical Methane Potential (Bth)
2.3.2. Water Potential (Ψ) Inhibition Assay
2.4. Analysis of the Microbial Consortium
DNA Extraction and Quantification
2.5. Chemical Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Correlation Factors with Ψ
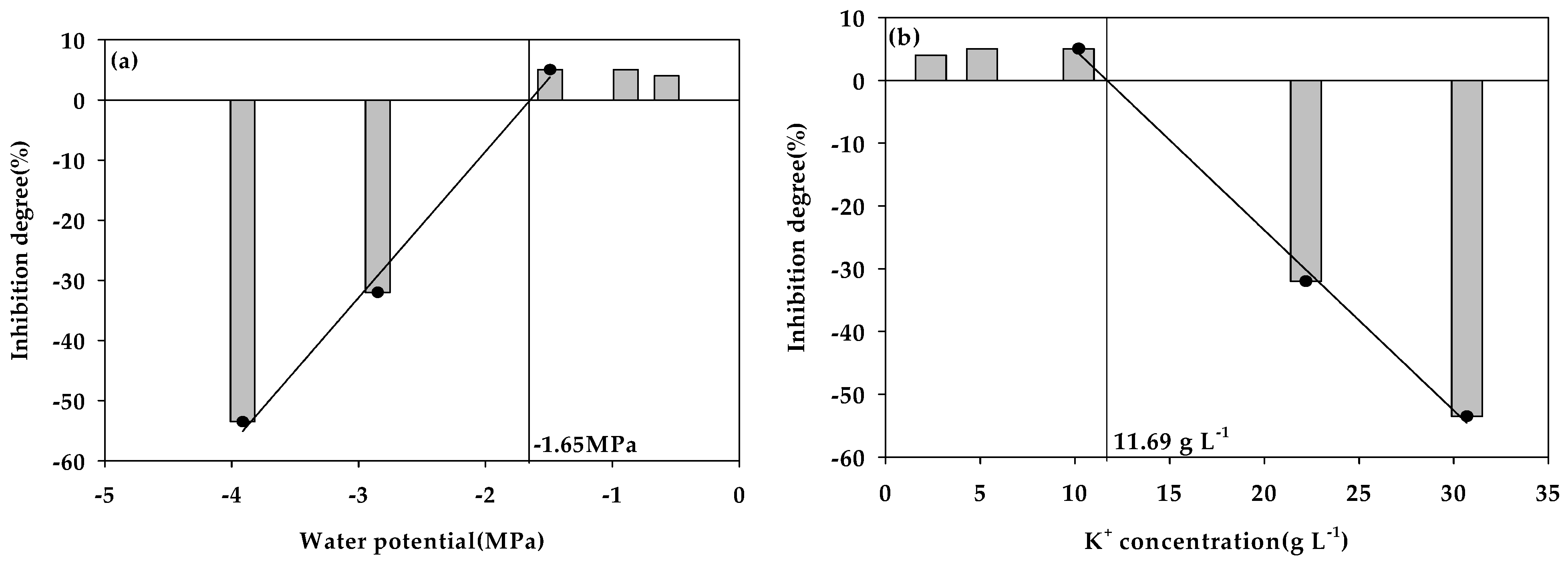
3.2. Water Potential (Ψ) Inhibition Assay
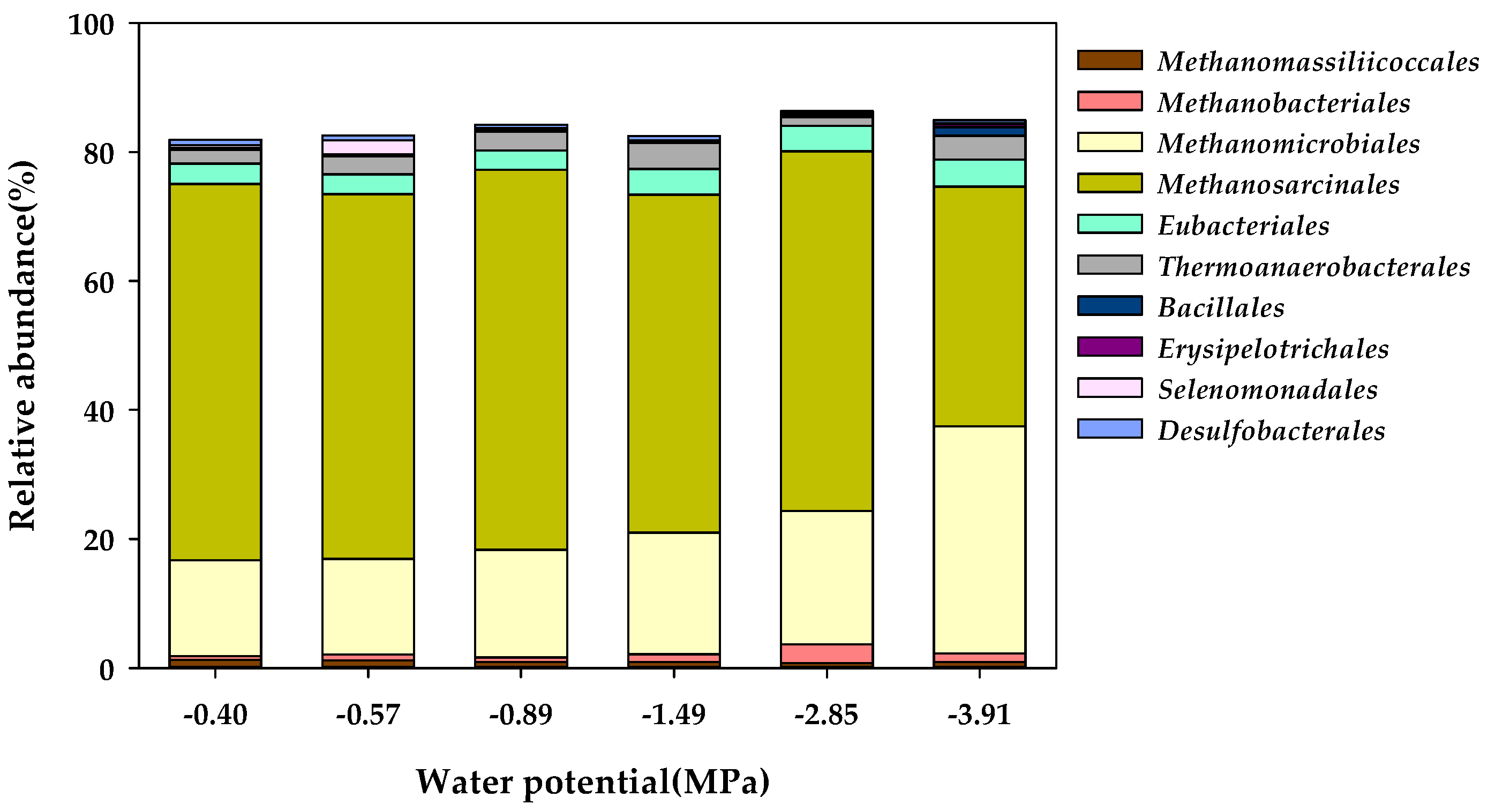
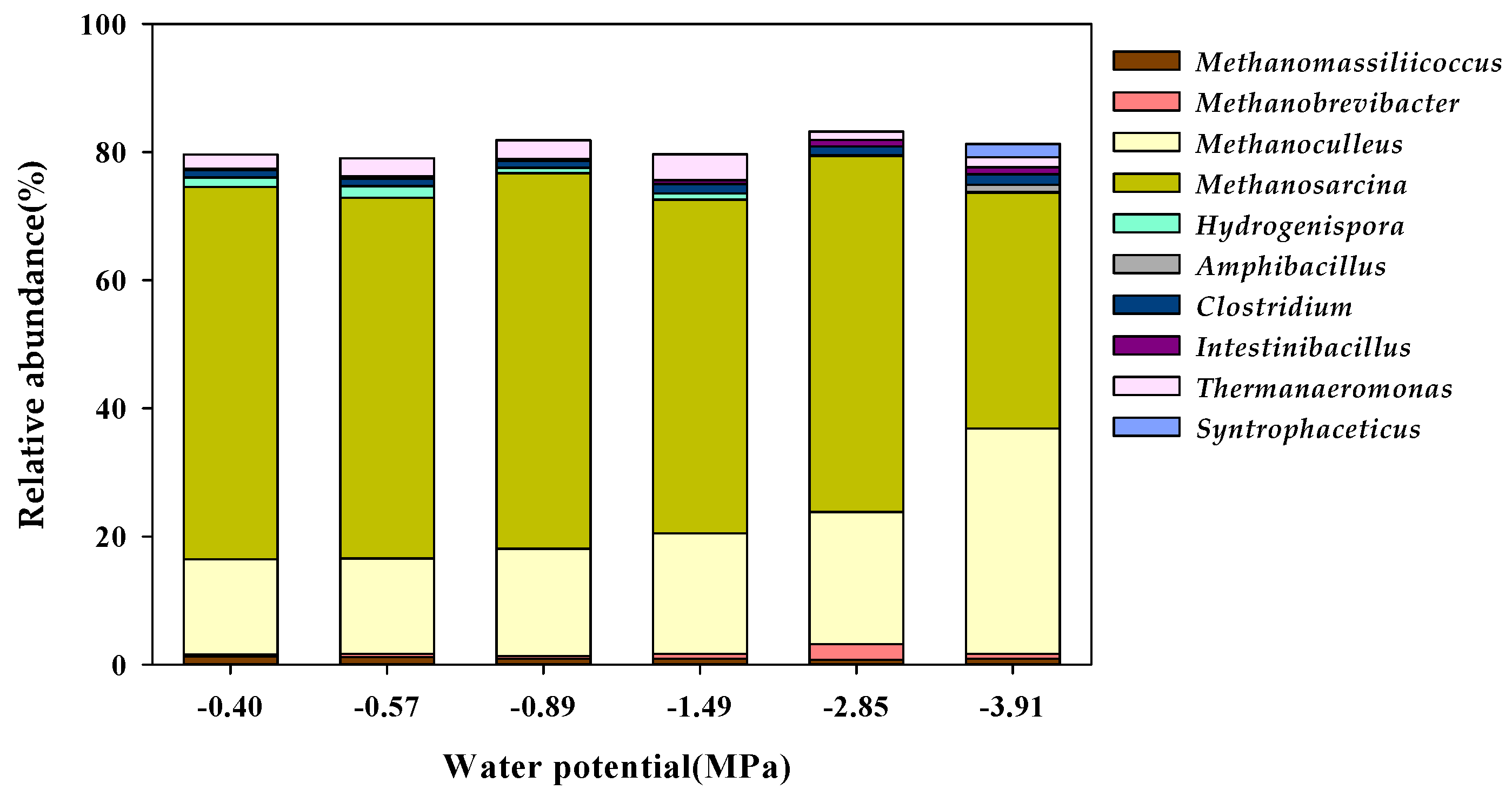
3.3. Microbial Community
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Björnsson, L.; Murto, M.; Jantsch, T.G.; Mattiasson, B. Evaluation of new methods for the monitoring of alkalinity, dissolved hydrogen and the microbial community in anaerobic digestion. Water Res. 2001, 35, 2833–2840. [Google Scholar] [CrossRef] [PubMed]
- Beuvink, J.; Spoelstra, S.; Hogendorp, R. An automated method for measuring time-course of gas production of feedstuffs incubated with buffered rumen fluid. Neth. J. Agric. Sci. 1992, 40, 401–407. [Google Scholar] [CrossRef]
- Ahring, B.K.; Sandberg, M.; Angelidaki, I. Volatile fatty acids as indicators of process imbalance in anaerobic digestors. Appl. Microbiol. Biotechnol. 1995, 43, 559–565. [Google Scholar] [CrossRef]
- Kroeker, E.; Schulte, D.; Sparling, A.; Lapp, H. Anaerobic treatment process stability. J. (Water Pollut. Control Fed.) 1979, 51, 718–727. [Google Scholar]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]
- Oh, S.-E.; Lee, M.-K.; Kim, D.-H. Continuous mesophilic-dry anaerobic digestion of organic solid waste. J. Korean Soc. Environ. Eng. 2009, 31, 341–345. [Google Scholar]
- Oh, K.-S.; Hwang, J.-K.; Song, Y.-J.; Kim, M.-J.; Park, J.-G.; Pak, D.-W. Integrated digestion of thermal solubilized sewage sludge to improve anaerobic digestion efficiency of organic waste. J. Korean Soc. Water Environ. 2022, 38, 95–102. [Google Scholar] [CrossRef]
- Lee, S.; Yoon, Y.-S.; Kang, J.-G.; Kim, K.-H.; Shin, S.K. Anaerobic co-digestion characteristics of food waste leachate and sewage sludge. J. Korea Org. Resour. Recycl. Assoc. 2016, 24, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Kim, S.-H.; Shin, H.-S. Sodium inhibition of fermentative hydrogen production. Int. J. Hydrogen Energy 2009, 34, 3295–3304. [Google Scholar] [CrossRef]
- McCarty, P.L.; McKinney, R.E. Salt toxicity in anaerobic digestion. J. (Water Pollut. Control Fed.) 1961, 33, 399–415. [Google Scholar]
- Zha, X.; Tsapekos, P.; Alvarado-Morales, M.; Lu, X.; Angelidaki, I. Potassium inhibition during sludge and biopulp co-digestion; experimental and model-based approaches. Waste Manag. 2020, 113, 304–311. [Google Scholar] [CrossRef]
- Schmidt, J.; Ahring, B. Effects of magnesium on thermophilic acetate-degrading granules in upflow anaerobic sludge blanket (UASB) reactors. Enzym. Microb. Technol. 1993, 15, 304–310. [Google Scholar] [CrossRef]
- Romero-Güiza, M.; Mata-Alvarez, J.; Chimenos, J.; Astals, S. The effect of magnesium as activator and inhibitor of anaerobic digestion. Waste Manag. 2016, 56, 137–142. [Google Scholar] [CrossRef] [PubMed]
- McCarty, P.L. Anaerobic waste treatment fundamentals. Public Works 1964, 95, 107–112. [Google Scholar]
- Jard, G.; Jackowiak, D.; Carrère, H.; Delgenès, J.-P.; Torrijos, M.; Steyer, J.-P.; Dumas, C. Batch and semi-continuous anaerobic digestion of Palmaria palmata: Comparison with Saccharina latissima and inhibition studies. Chem. Eng. J. 2012, 209, 513–519. [Google Scholar] [CrossRef]
- Fang, C.; Boe, K.; Angelidaki, I. Anaerobic co-digestion of desugared molasses with cow manure; focusing on sodium and potassium inhibition. Bioresour. Technol. 2011, 102, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Zhang, R.; Wu, S.; Liu, Z.; Qiu, B.; Fang, Y.; Sun, D. Calcium effect on anaerobic biological treatment of fresh leachate with extreme high calcium concentration. Int. Biodeterior. Biodegrad. 2014, 95, 76–83. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J. Effect of potassium inhibition on the thermophilic anaerobic digestion of swine waste. Water Environ. Res. 2007, 79, 667–674. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Y.; Liu, S.; Pan, M.; Yang, J.; Chen, S. Denitrification activities and N2O production under salt stress with varying COD/N ratios and terminal electron acceptors. Chem. Eng. J. 2013, 215, 252–260. [Google Scholar] [CrossRef]
- Wang, R.; Zheng, P.; Ding, A.-q.; Zhang, M.; Ghulam, A.; Yang, C.; Zhao, H.-P. Effects of inorganic salts on denitrifying granular sludge: The acute toxicity and working mechanisms. Bioresour. Technol. 2016, 204, 65–70. [Google Scholar] [CrossRef]
- Phae, C.-G.; Chu, Y.-S.; Park, J.-S. Investigation of effect on composting process and plant growth of salt concentration in food waste. J. Korea Org. Resour. Recycl. Assoc. 2002, 10, 103–111. [Google Scholar]
- Lupin, H.; Boeri, R.; Moschiar, S. Water activity and salt content relationship in moist salted fish products. Int. J. Food Sci. Technol. 1981, 16, 31–38. [Google Scholar] [CrossRef]
- Bremer, E.; Krämer, R. Responses of microorganisms to osmotic stress. Annu. Rev. Microbiol. 2019, 73, 313–334. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M. Bacterial osmoregulation: A paradigm for the study of cellular homeostasis. Annu. Rev. Microbiol. 2011, 65, 215–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-W.; Shin, H.-H.; Kim, J.-G. A study of dewatering phenomena of potato slice cytorrhysed by high molecules. Korean J. Food Nutr. 2006, 19, 358–365. [Google Scholar]
- Money, N.P. Measurement of pore size in the hyphal cell wall of Achlya bisexualis. Exp. Mycol. 1990, 14, 234–242. [Google Scholar] [CrossRef]
- Griffin, D. Water and microbial stress. Adv. Microb. Ecol. 1981, 5, e136. [Google Scholar]
- Rawlins, S.L.; Campbell, G.S. Water potential: Thermocouple psychrometry. Methods Soil Anal. Part 1 Phys. Mineral. Methods 1986, 5, 597–618. [Google Scholar] [CrossRef]
- Angelidaki, I.; Alves, M.; Bolzonella, D.; Borzacconi, L.; Campos, J.L.; Guwy, A.J.; Kalyuzhnyi, S.; Jenicek, P.; van Lier, J.B. Defining the biomethane potential (BMP) of solid organic wastes and energy crops: A proposed protocol for batch assays. Water Sci. Technol. 2009, 59, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Lay, J.-J.; Li, Y.-Y.; Noike, T. Mathematical model for methane production from landfill bioreactor. J. Environ. Eng. 1998, 124, 730–736. [Google Scholar] [CrossRef]
- Unesco. Background Papers and Supporting Data on the Practical Salinity Scale 1978-Unesco; Joint Panel on Oceanographic Tables and Standards: Woods Hole, MA, USA, 1981. [Google Scholar]
- Oh, S.-Y.; Yoon, Y.-M. Energy recovery efficiency of poultry slaughterhouse sludge cake by hydrothermal carbonization. Energies 2017, 10, 1876. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, A.H.; Winther-Nielsen, M.; Ahring, B.K. Kinetics of lactate, acetate and propionate in unadapted and lactate-adapted thermophilic, anaerobic sewage sludge: The influence of sludge adaptation for start-up of thermophilic UASB-reactors. Appl. Microbiol. Biotechnol. 1991, 34, 823–827. [Google Scholar] [CrossRef]
- Hansen, K.H.; Angelidaki, I.; Ahring, B.K. Anaerobic digestion of swine manure: Inhibition by ammonia. Water Res. 1998, 32, 5–12. [Google Scholar] [CrossRef]
- Trejo-Téllez, L.I.; Gómez-Merino, F.C. Nutrient solutions for hydroponic systems. In Hydroponics: A Standard Methodology for Plant Biological Research; IntechOpen: London, UK, 2012; pp. 1–22. [Google Scholar]
- Sterne, R.; Zentmyer, G.; Bingham, F. The effect of osmotic potential and specific ions on growth of Phytophthora cinnamomi. Phytopathology 1976, 66, 1398–1402. [Google Scholar] [CrossRef]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar] [CrossRef] [Green Version]
- Fougere, F.; Le Rudulier, D.; Streeter, J.G. Effects of salt stress on amino acid, organic acid, and carbohydrate composition of roots, bacteroids, and cytosol of alfalfa (Medicago sativa L.). Plant Physiol. 1991, 96, 1228–1236. [Google Scholar] [CrossRef] [Green Version]
- Brown, A. Microbial water stress. Bacteriol. Rev. 1976, 40, 803–846. [Google Scholar] [CrossRef]
- Killham, K.; Firestone, M. Salt stress control of intracellular solutes in streptomycetes indigenous to saline soils. Appl. Environ. Microbiol. 1984, 47, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Song, C.-W.; Kim, N.-C.; Ryu, J.-K.; Kim, J.-M. A study on the fuelization of livestock sludge using thermal hydrolysis. J. Korea Org. Resour. Recycl. Assoc. 2015, 23, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardwell, T.; Cattrall, R. Determination of ammonia in waste waters by a differential pH method using flow injection potentiometry and a nonactin-based sensor. Analyst 1997, 122, 89–93. [Google Scholar] [CrossRef]
- Deublein, D.; Steinhauser, A. Biogas from Waste and Renewable Resources: An Introduction; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Lohani, S.P.; Havukainen, J. Anaerobic digestion: Factors affecting anaerobic digestion process. In Waste Bioremediation; Springer: Singapore, 2018; pp. 343–359. [Google Scholar] [CrossRef]
- Migliore, G.; Alisi, C.; Sprocati, A.; Massi, E.; Ciccoli, R.; Lenzi, M.; Wang, A.; Cremisini, C. Anaerobic digestion of macroalgal biomass and sediments sourced from the Orbetello lagoon, Italy. Biomass Bioenergy 2012, 42, 69–77. [Google Scholar] [CrossRef]
- Sowers, K.R.; Gunsalus, R.P. Halotolerance in Methanosarcina spp.: Role of N (sup (epsilon))-acetyl-(beta)-lysine, (alpha)-glutamate, glycine betaine, and K (sup+) as compatible solutes for osmotic adaptation. Appl. Environ. Microbiol. 1995, 61, 4382–4388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGenity, T.J.; Sorokin, D.Y. Methanogens and methanogenesis in hypersaline environments. In Biogenesis of Hydrocarbons; Handbook of Hydrocarbon and Lipid Microbiology; Springer: Cham, Switzerland, 2010; pp. 665–680. [Google Scholar] [CrossRef]
- Lai, M.-C.; Sowers, K.R.; Robertson, D.E.; Roberts, M.F.; Gunsalus, R.P. Distribution of compatible solutes in the halophilic methanogenic archaebacteria. J. Bacteriol. 1991, 173, 5352–5358. [Google Scholar] [CrossRef] [Green Version]
- Harris, R. Effect of water potential on microbial growth and activity. Water Potential Relat. Soil Microbiol. 1981, 9, 23–95. [Google Scholar] [CrossRef]
- Carpita, N.; Sabularse, D.; Montezinos, D.; Delmer, D.P. Determination of the pore size of cell walls of living plant cells. Science 1979, 205, 1144–1147. [Google Scholar] [CrossRef]
- Wen, X.; Yang, S.; Horn, F.; Winkel, M.; Wagner, D.; Liebner, S. Global biogeographic analysis of methanogenic archaea identifies community-shaping environmental factors of natural environments. Front. Microbiol. 2017, 8, 1339. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.N.; Boyd, J.A.; Leu, A.O.; Woodcroft, B.J.; Parks, D.H.; Hugenholtz, P.; Tyson, G.W. An evolving view of methane metabolism in the Archaea. Nat. Rev. Microbiol. 2019, 17, 219–232. [Google Scholar] [CrossRef]
- Mand, T.D.; Metcalf, W.W. Energy conservation and hydrogenase function in methanogenic archaea, in particular the genus Methanosarcina. Microbiol. Mol. Biol. Rev. 2019, 83, e00020-19. [Google Scholar] [CrossRef]
- Adams, M.W.; Maier, R.J.; Wiegel, J. Incredible anaerobes from physiology to genomics to fuels. Ann. N. Y. Acad. Sci. 2008, 1125. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Merkel, A.Y.; Abbas, B.; Makarova, K.S.; Rijpstra, W.I.C.; Koenen, M.; Damsté, J.S.S.; Galinski, E.A.; Koonin, E.V.; Van Loosdrecht, M.C. Methanonatronarchaeum thermophilum gen. nov., sp. nov. and Candidatus Methanohalarchaeum thermophilum, extremely halo (natrono) philic methyl-reducing methanogens from hypersaline lakes comprising a new euryarchaeal class Methanonatronarchaeia classis nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 2199. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.-z.; Fang, S.; Zhang, L.; Huang, W.; Shao, Q.; Fang, F.; Feng, Q.; Cao, J.; Luo, J. Distribution patterns of functional microbial community in anaerobic digesters under different operational circumstances: A review. Bioresour. Technol. 2021, 341, 125823. [Google Scholar] [CrossRef] [PubMed]
- Baek, G.; Kim, J.; Kim, J.; Lee, C. Role and potential of direct interspecies electron transfer in anaerobic digestion. Energies 2018, 11, 107. [Google Scholar] [CrossRef] [Green Version]
- Aydin, S. Enhanced biodegradation of antibiotic combinations via the sequential treatment of the sludge resulting from pharmaceutical wastewater treatment using white-rot fungi Trametes versicolor and Bjerkandera adusta. Appl. Microbiol. Biotechnol. 2016, 100, 6491–6499. [Google Scholar] [CrossRef] [PubMed]
- Alenzi, A.; Hunter, C.; Spencer, J.; Roberts, J.; Craft, J.; Pahl, O.; Escudero, A. Pharmaceuticals effect and removal, at environmentally relevant concentrations, from sewage sludge during anaerobic digestion. Bioresour. Technol. 2021, 319, 124102. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Input Materials | Capacity | Digester Temperature | HRT 1 | Location |
|---|---|---|---|---|---|
| (m3 Day−1) | (°C) | (Days) | |||
| 1 | Sewage sludge | 2863 | 37.5 | 15.5 | Gangnam, Seoul |
| 2 | Pig slurry, food wastewater | 200 | 36.8 | 25.0 | Goyang, Gyeonggi |
| 3 | Pig slurry, food wastewater, livestock byproducts | 150 | 36.0 | 30.0 | Nonsan, Chungnam |
| 4 | Sewage sludge, food wastewater | 2000 | 37.5 | 25.0 | Dongnae, Busan |
| 5 | Pig slurry, food waste, sewage sludge | 320 | 37.0 | 35.0 | Seosan, Chungnam |
| 6 | Sewage sludge | 2700 | 37.5 | 30.0 | Seongnam, Gyeonggi |
| 7 | Sewage sludge, food wastewater | 140 | 34.5 | 20.0 | Ansan, Gyeonggi |
| 8 | Pig slurry, food wastewater, sewage sludge | 384 | 36.0 | 15.0 | Anyang, Gyeonggi |
| 9 | Pig slurry, food waste | 130 | 36.0 | 20.0 | Yangsan, Gyeongnam |
| 10 | Pig slurry, food wastewater | 100 | 40.0 | 35.0 | Yeoncheon, Gyeonggi |
| 11 | Food waste, slaughterhouse byproducts | 220 | 40.0 | 55.0 | Wonju, Gangwon |
| 12 | Pig slurry, food wastewater | 100 | 38.0 | 40.0 | Icheon, Gyeonggi |
| 13 | Food wastewater | 650 | 39.0 | 43.0 | Seo, Incheon |
| 14 | Food wastewater | 300 | 36.0 | 25.0 | Jeonju, Jeonbuk |
| 15 | Pig slurry, food wastewater | 125 | 38.0 | 11.0 | Seogwipo, Jeju |
| 16 | Food wastewater | 150 | 39.5 | 26.0 | Cheongju, Chungbuk |
| 17 | Sewage sludge, food wastewater | 525 | 38.0 | 30.7 | Chuncheon, Gangwon |
| 18 | Food waste | 80 | 38.0 | 19.0 | Chungju, Chungbuk |
| 19 | Pig slurry, food wastewater | 100 | 37.0 | 35.0 | Hongcheon, Gangwon |
| 20 | Food wastewater | 120 | 39.5 | 30.0 | Hwaseong, Gyeonggi |
| Parameters | pH | TS 1 | VS 2 | TCOD 3 | SCOD 4 | TKN 5 | NH4+-N 6 | Alkalinity | TVFAs 7 |
|---|---|---|---|---|---|---|---|---|---|
| (-) | (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1 as CaCO3) | (mg L−1 as Acetate) | |
| Inoculum | 7.96 | 30,044 | 14,767 | 21,050 | 6012 | 2201 | 1314 | 6389 | 113 |
| Parameters | Blank | Control | Treatments | |||||
|---|---|---|---|---|---|---|---|---|
| K1 | K2 | K3 | K4 | K5 | ||||
| Inoculum (mL) | 70 | 70 | 70 | 70 | 70 | 70 | 70 | |
| Substrate 1 (g) | 0 | 0.52 | 0.52 | 0.52 | 0.52 | 0.52 | 0.52 | |
| I/S ratio 2 (-) | - | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | |
| WP 3 adjusting solution | KCl conc. (%) | - | - | 0.46 | 0.98 | 1.95 | 4.23 | 5.85 |
| Volume (mL) | - | - | 130 | 130 | 130 | 130 | 130 | |
| DW 4 volume (mL) | 130 | 130 | - | - | - | - | - | |
| Operation volume (mL) | 200 | 200 | 200 | 200 | 200 | 200 | 200 | |
| Ψ 5 (MPa) | −0.36 (0.03) 6 | −0.40 (0.03) | −0.57 (0.03) | −0.89 (0.03) | −1.49 (0.04) | −2.85 (0.01) | −3.91 (0.02) | |
| K+ conc. 7 (%) | N.A. 8 | N.A. | 0.24 | 0.51 | 1.02 | 2.22 | 3.07 | |
| Parameters | Mean | Median | Maximum | Minimum | r 15 | |
|---|---|---|---|---|---|---|
| Ψ 1 (MPa) | −1.23 | −1.25 | −0.10 | −2.09 | - | |
| pH (-) | 7.86 | 7.83 | 8.78 | 6.89 | −0.50 ** | |
| WC 2 (wt. %, w.b.) | 96.20 | 96.30 | 98.54 | 92.13 | 0.42 * | |
| TCOD 3 (g L−1) | 30.95 | 32.34 | 52.43 | 14.30 | −0.45 * | |
| SCOD 4 (g L−1) | 9.33 | 9.67 | 24.63 | 0.00 | −0.68 *** | |
| TKN 5 (g L−1) | 4.91 | 5.66 | 6.95 | 1.05 | −0.87 *** | |
| TAN 6 (g L−1) | 3.27 | 3.30 | 5.19 | 0.30 | −0.91 *** | |
| NH4+-N 7 (g L−1) | 2.87 | 2.87 | 4.57 | 0.30 | - | |
| Alkalinity (g L−1 as CaCO3) | 16.82 | 16.57 | 41.60 | 1.45 | −0.61 *** | |
| TVFAs 8 | (cmolc L−1 as acetate) | 1.30 | 0.50 | 10.34 | 0.19 | −0.12 |
| (mg L−1 as acetate) | 778 | 302 | 6210 | 113 | - | |
| Solid content | TS 9 (g L−1) | 37.96 | 37.03 | 78.72 | 14.60 | −0.42 * |
| VS 10 (g L−1) | 20.00 | 20.15 | 28.71 | 10.62 | −0.50 ** | |
| FS 11 (g L−1) | 17.96 | 16.78 | 53.63 | 3.98 | −0.32 | |
| TSS 12 (g L−1) | 31.04 | 27.85 | 64.90 | 15.43 | −0.06 | |
| VSS 13 (g L−1) | 20.51 | 19.85 | 33.80 | 9.63 | −0.28 | |
| FSS 14 (g L−1) | 15.18 | 10.53 | 38.65 | 3.95 | −0.33 | |
| Parameters | Mean | Median | Maximum | Minimum | r 3 | |
|---|---|---|---|---|---|---|
| Ψ 1 (MPa) | −1.23 | −1.25 | −0.10 | −2.09 | - | |
| SPS 2 (p.s.u.) | 15.49 | 16.01 | 26.52 | 1.27 | −0.90 *** | |
| NH4+ | (cmolc L−1) | 20.48 | 20.47 | 32.62 | 2.16 | −0.89 *** |
| (mg L−1) | 3694 | 3693 | 5884 | 389 | - | |
| Na+ | (cmolc L−1) | 7.78 | 7.34 | 27.92 | 0.52 | −0.51 *** |
| (mg L−1) | 1789 | 1688 | 6419 | 119 | - | |
| K+ | (cmolc L−1) | 5.79 | 5.63 | 22.51 | 0.16 | −0.68 *** |
| (mg L−1) | 2263 | 2202 | 8802 | 64 | - | |
| Mg2+ | (cmolc L−1) | 0.38 | 0.37 | 0.81 | 0.03 | −0.47 ** |
| (mg L−1) | 46.58 | 44.97 | 98.75 | 3.78 | - | |
| Ca2+ | (cmolc L−1) | 1.17 | 0.72 | 10.26 | 0.20 | −0.07 |
| (mg L−1) | 234 | 144 | 2056 | 39 | - | |
| Cl− | (cmolc L−1) | 10.94 | 9.68 | 32.06 | 0.37 | −0.71 *** |
| (mg L−1) | 3879 | 3432 | 11,366 | 132 | - | |
| NO3− | (cmolc L−1) | 0.03 | 0.00 | 0.08 | 0.00 | −0.60 *** |
| (mg L−1) | 17.90 | 0.00 | 47.22 | 0.00 | - | |
| SO42− | (cmolc L−1) | 0.12 | 0.04 | 1.14 | 0.00 | −0.49 ** |
| (mg L−1) | 58.97 | 17.33 | 549.30 | 0.00 | - | |
| PO43− | (cmolc L−1) | 0.88 | 0.79 | 4.07 | 0.00 | −0.67 *** |
| (mg L−1) | 279.07 | 250.35 | 1287.00 | 0.00 | - | |
| Parameters | Fermentation Time (Days) | Control | Treatments 1 | ||||
|---|---|---|---|---|---|---|---|
| K1 | K2 | K3 | K4 | K5 | |||
| TVFAs 2 (mg/L as acetate) | 0 | 105 (0.26) 4 | 104 (0.22) | 105 (0.25) | 105 (0.09) | 105 (0.21) | 105 (0.08) |
| 12 | 1461 (0.09) | 1474 (3.26) | 1448 (7.71) | 1261 (2.44) | 1789 (10.08) | 932 (2.87) | |
| 45 | 109 (2.97) | 112 (0.81) | 107 (3.72) | 111 (1.04) | 1393 (5.99) | 1357 (5.95) | |
| 78 | 116 (0.33) | 123 (12.55) | 116 (0.56) | 115 (0.42) | 266 (1.33) | 399 (10.35) | |
| Ψ 3 (MPa) | 0 | −0.37 (0.01) | −0.55 (0.05) | −0.86 (0.01) | −1.46 (0.01) | −2.84 (0.00) | −3.94 (0.01) |
| 12 | −0.39 (0.01) | −0.55 (0.02) | −0.88 (0.00) | −1.46 (0.00) | −2.84 (0.03) | −3.91 (0.04) | |
| 45 | −0.42 (0.02) | −0.62 (0.01) | −0.92 (0.02) | −1.56 (0.01) | −2.86 (0.01) | −3.92 (0.03) | |
| 78 | −0.43 (0.040 | −0.58 (0.03) | −0.93 (0.02) | −1.49 (0.01) | −2.86 (0.00) | −3.90 (0.04) | |
| Parameters | Control | Treatments 1 | |||||
|---|---|---|---|---|---|---|---|
| K1 | K2 | K3 | K4 | K5 | |||
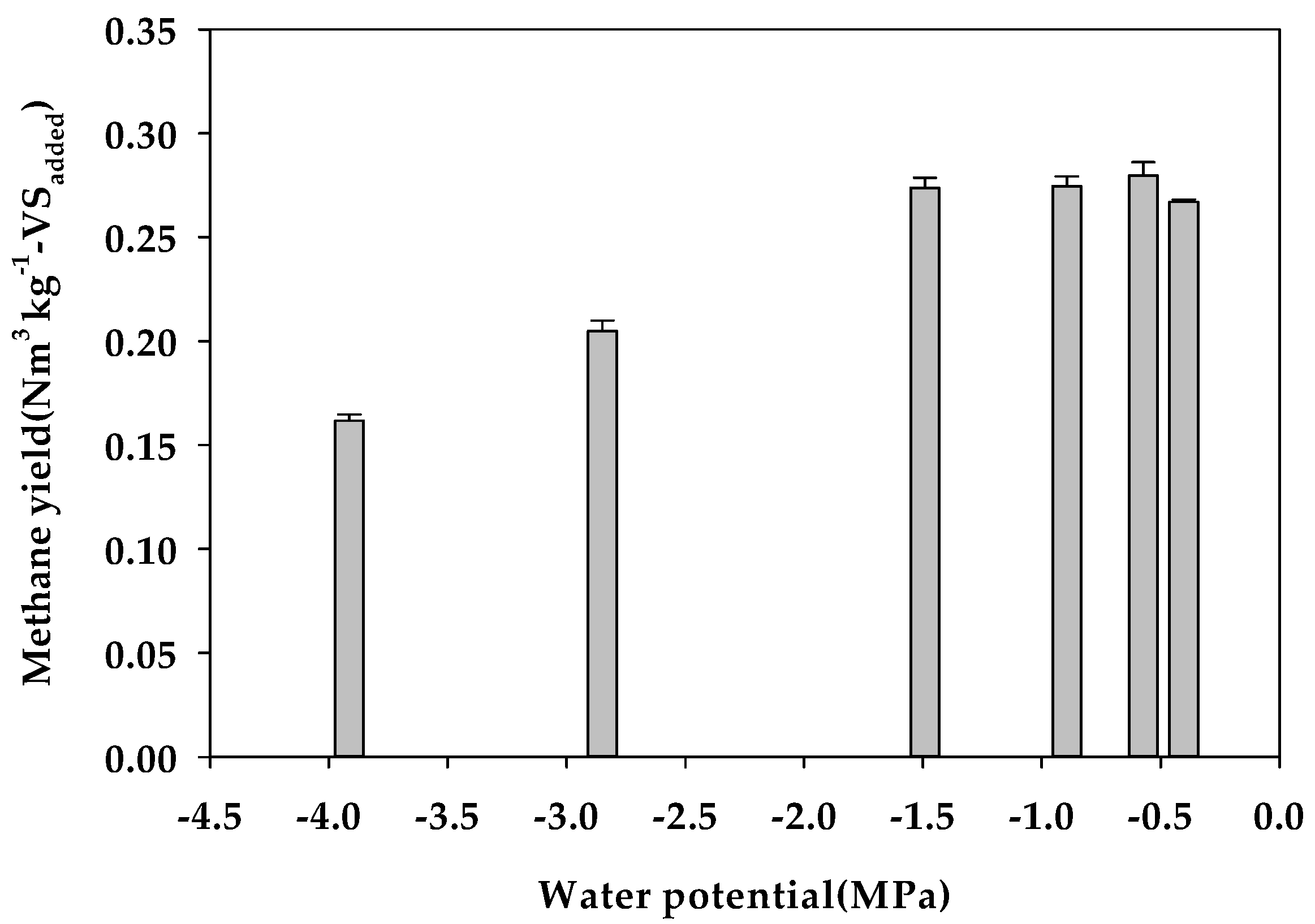
| Ψ 2 (MPa) | −0.40 | −0.57 | −0.89 | −1.49 | −2.85 | −3.91 | |
| VSr 3 (%) | 70.64 c | 73.63 b | 74.55 a | 74.54 a | 54.38 d | 46.21 e | |
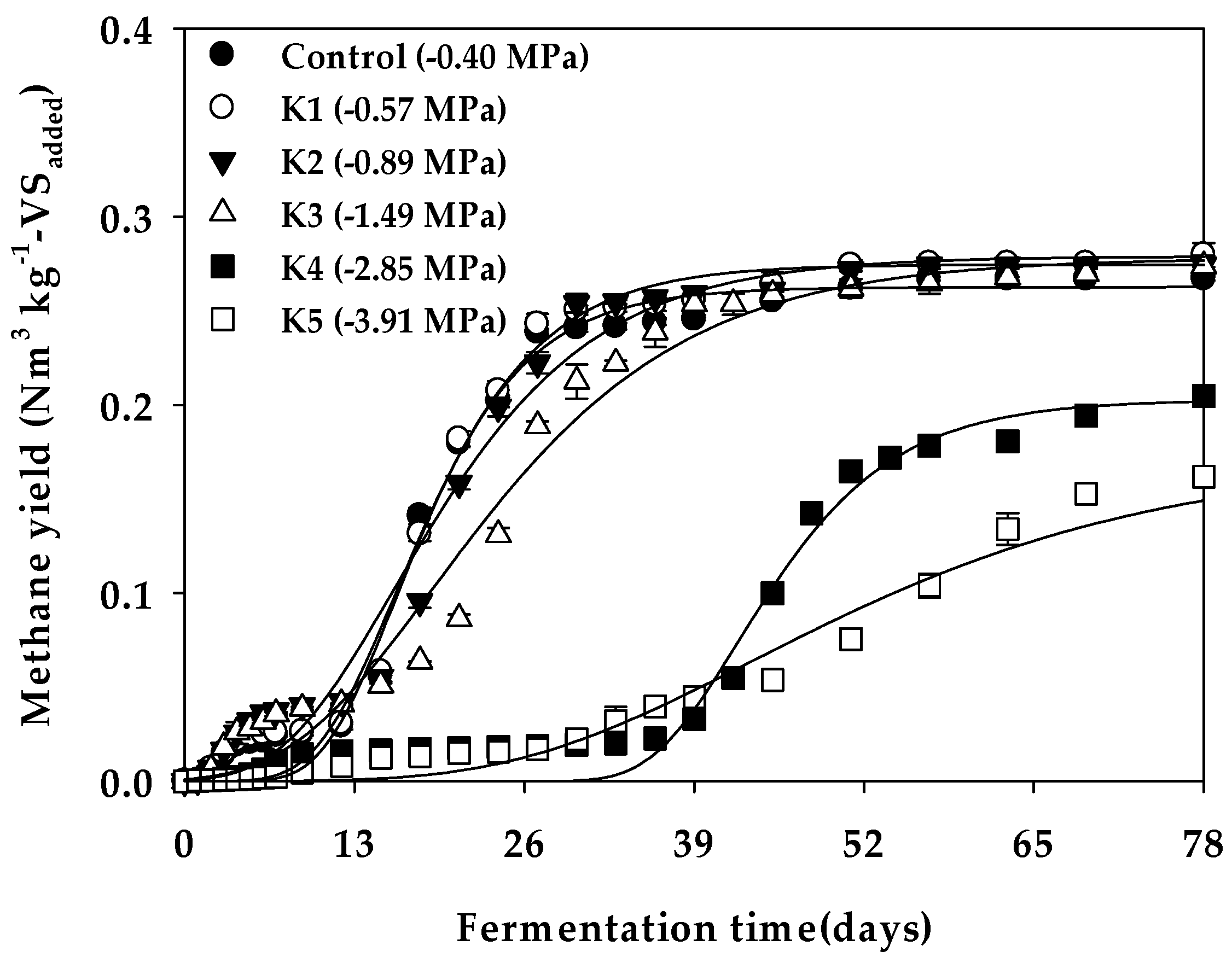
| Model parameters | Bu 4 (Nm3 kg−1-VSadded) | 0.264 c | 0.275 b | 0.278 a | 0.278 a | 0.203 d | 0.172 e |
| p 5 (mL) | 127 c | 132 b | 134 a | 134 a | 98 d | 83 e | |
| Rm 6 (mL day−1) | 8.11 a | 7.92 b | 5.84 c | 4.67 e | 5.48 d | 1.70 f | |
| λ 7 (days) | 10.34 c | 10.16 c | 7.80 e | 8.65 d | 35.96 a | 25.34 b | |
| R2 | 0.990 | 0.990 | 0.984 | 0.983 | 0.982 | 0.985 | |
| Reactor Type | Temperature | Substrate | Concentrations (mg L−1) | Reference | |
|---|---|---|---|---|---|
| Stimulatory | Inhibitory | ||||
| UASB 1 | 35 °C | IW 3, SS 4 | 200~400 | 12,000 | [14] |
| CSTR 2 | 50 °C | PS 5 | - | 3000 | [18] |
| Batch | 55 °C | CM 6 | - | 28,000 (50% inhibition) | [16] |
| Batch | 35 °C | MA 7 | - | 13,000 (52~53% inhibition) | [15] |
| Batch | 54 °C | SS | - | 8000 (50% inhibition) | [11] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeo, J.; Kim, C.-G.; Lee, J.-H.; Song, E.; Yoon, Y.-M. Effects of Water Potential on Anaerobic Methane Production and a Microbial Consortium. Fermentation 2023, 9, 244. https://doi.org/10.3390/fermentation9030244
Yeo J, Kim C-G, Lee J-H, Song E, Yoon Y-M. Effects of Water Potential on Anaerobic Methane Production and a Microbial Consortium. Fermentation. 2023; 9(3):244. https://doi.org/10.3390/fermentation9030244
Chicago/Turabian StyleYeo, Jin, Chang-Gyu Kim, Jun-Hyeong Lee, Eunhye Song, and Young-Man Yoon. 2023. "Effects of Water Potential on Anaerobic Methane Production and a Microbial Consortium" Fermentation 9, no. 3: 244. https://doi.org/10.3390/fermentation9030244
APA StyleYeo, J., Kim, C. -G., Lee, J. -H., Song, E., & Yoon, Y. -M. (2023). Effects of Water Potential on Anaerobic Methane Production and a Microbial Consortium. Fermentation, 9(3), 244. https://doi.org/10.3390/fermentation9030244