
Supplementing Yogurt with Probiotic Bifidobacteria to Counter Chronic Kidney Disease
 , , , , and
, , , , and 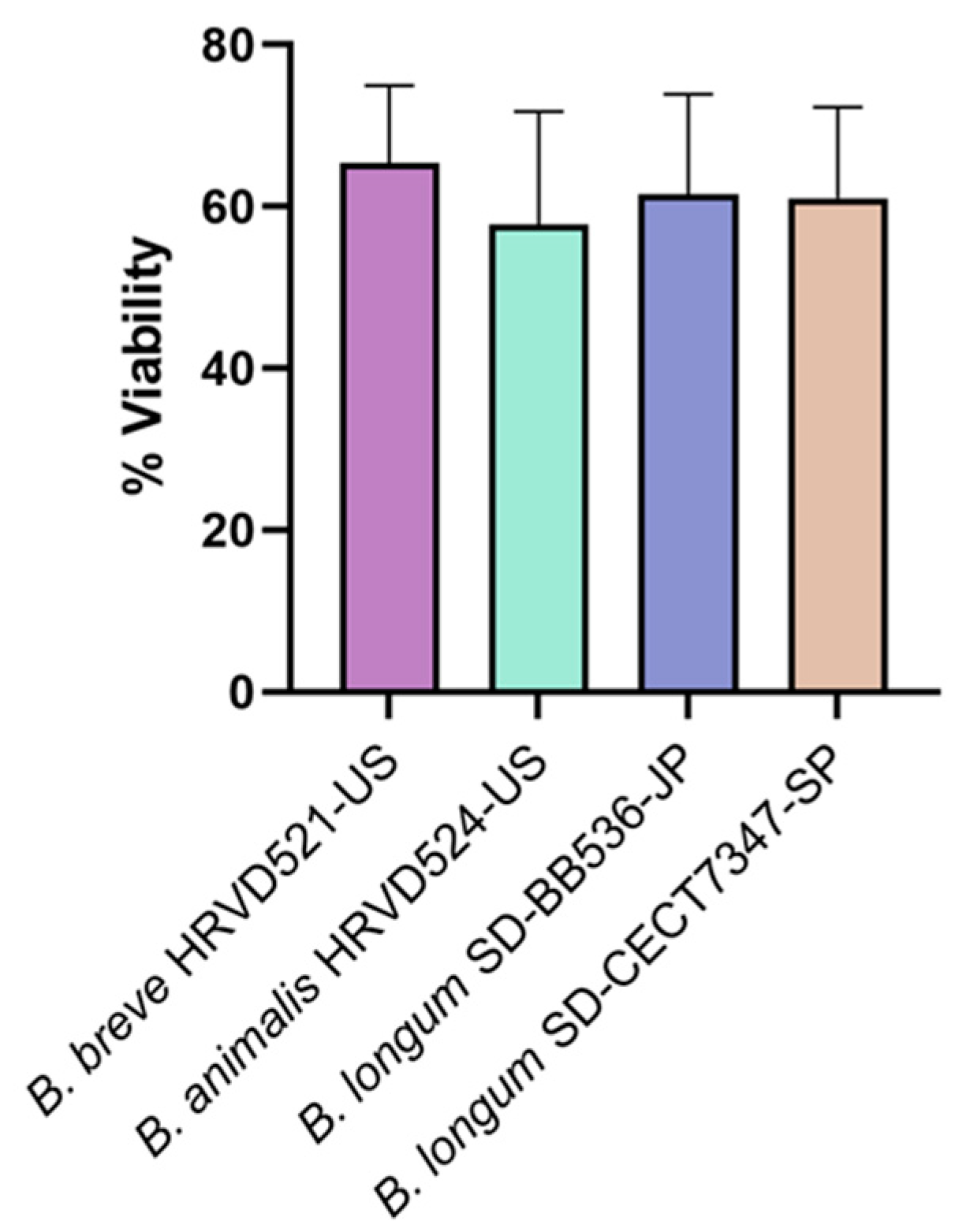{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bacterial Culture Conditions
2.3. Freeze Drying Bacteria
2.4. Production of Probiotic Yogurt Containing p-Cresol Reducing Bifidobacteria
2.5. CFU Quantification of Yogurt Bacteria
2.6. Sensory Evaluation
2.7. p-Cresol Clearance in a Simulated Colonic Environment
2.8. Sample Preparation and HPLC Analysis
2.9. Statistical Analysis
3. Results & Discussion
3.1. Viability of Lyophilized Probiotic Bifidobacteria
3.2. Survival of Probiotic Bacteria in Yogurt Samples
3.3. The pH of Yogurt Samples
3.4. Sensory Properties of Yogurt Samples
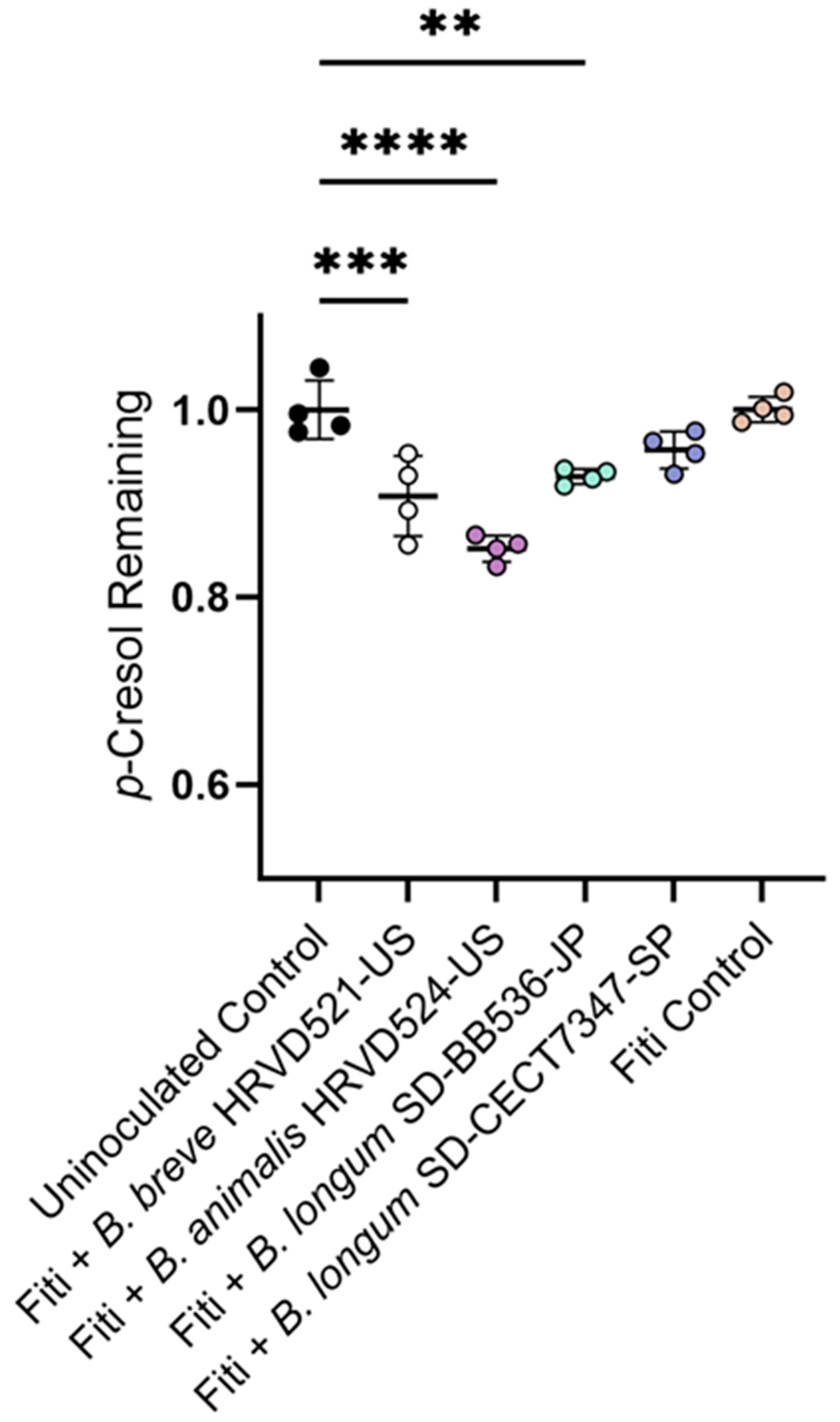
3.5. Yogurt Supplemented with Probiotic Bifidobacteria Reduce p-Cresol in a Simulated Colon Environment
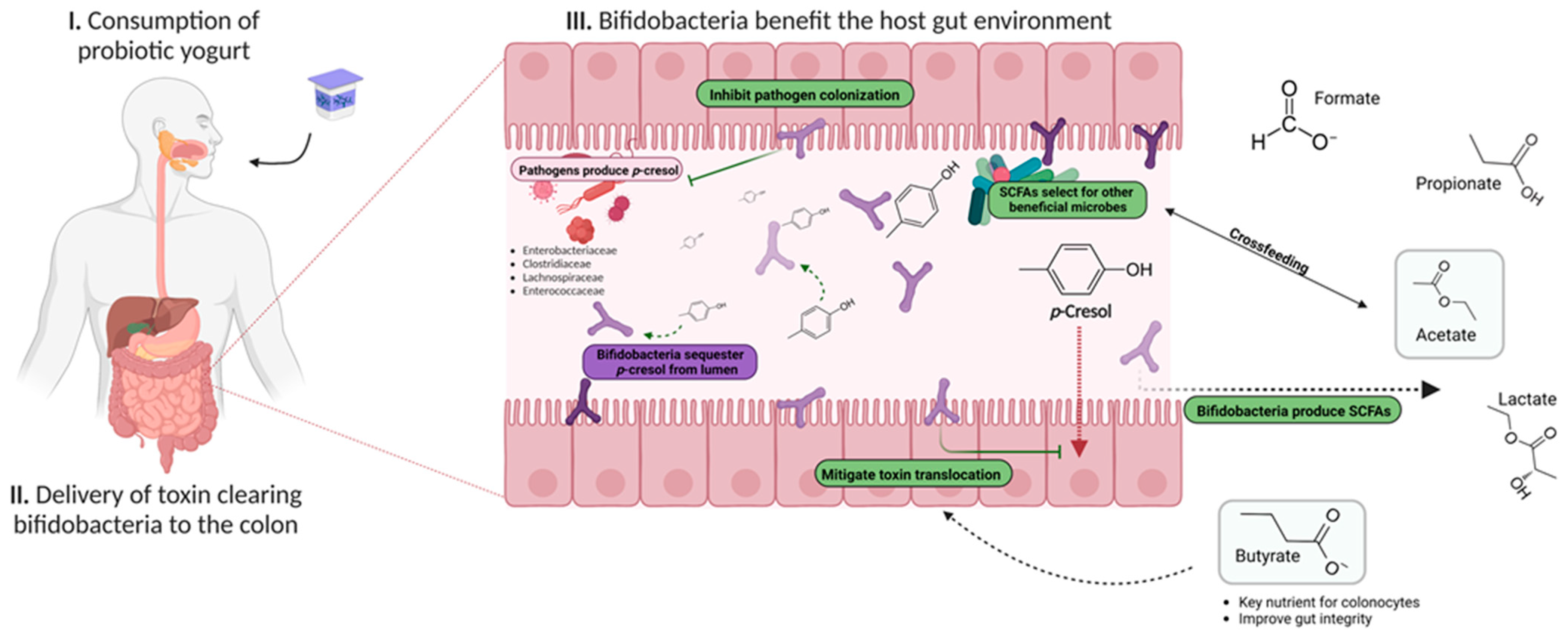
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D.R. Global prevalence of chronic kidney disease—A systematic review and meta-analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [PubMed]
- Peralta, C.A.; Risch, N.; Lin, F.; Shlipak, M.G.; Reiner, A.; Ziv, E.; Tang, H.; Siscovick, D.; Bibbins-Domingo, K. The association of African ancestry and elevated reatinine in the coronary artery risk development in young adults (CARDIA) Study. Am. J. Nurs. 2010, 31, 202–208. [Google Scholar]
- Genovese, G.; Friedman, D.J.; Ross, M.D.; Lecordier, L.; Uzureau, P.; Freedman, B.I.; Bowden, D.W.; Langefeld, C.D.; Oleksyk, T.K.; Uscinski Knob, A.L.; et al. Association of trypanolytic ApoL1 variants with kidney disease in african americans. Science 2010, 329, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Kaze, A.D.; Schutte, A.E.; Erqou, S.; Kengne, A.P.; Echouffo-Tcheugui, J.B. Prevalence of hypertension in older people in Africa: A systematic review and meta-analysis. J. Hypertens. 2017, 35, 1345–1352. [Google Scholar] [CrossRef]
- Foster, M.C.; Coresh, J.; Fornage, M.; Astor, B.C.; Grams, M.; Franceschini, N.; Boerwinkle, E.; Parekh, R.S.; Kao, W.H.L. ApoL1 variants associate with increased risk of CKD among African Americans. J. Am. Soc. Nephrol. 2013, 24, 1484–1491. [Google Scholar] [CrossRef]
- Peralta, C.A.; Katz, R.; DeBoer, I.; Ix, J.; Sarnak, M.; Kramer, H.; Siscovick, D.; Shea, S.; Szklo, M.; Shlipak, M. Racial and ethnic differences in kidney function decline among persons without chronic kidney disease. J. Am. Soc. Nephrol. 2011, 22, 1327–1334. [Google Scholar] [CrossRef]
- Young, B.A.; Maynard, C.; Boyko, E.J. Racial differences in diabetic nephropathy, cardiovascular disease, and mortality in a national population of veterans. Diabetes Care 2003, 26, 2392–2399. [Google Scholar] [CrossRef]
- Patrice, H.M.; Pascal, K.A.; François, K.F.; Hilaire, D.; Solange, D.M.; Gloria, A.E.; Pierre, C.S. Markers and Risk Factors for chronic kidney disease in sub-Saharan Africans: Baseline levels and 12-month trajectories in newly referred patients in Cameroon. BMC Nephrol. 2020, 21, 101. [Google Scholar] [CrossRef]
- Ene-Iordache, B.; Perico, N.; Bikbov, B.; Carminati, S.; Remuzzi, A.; Perna, A.; Islam, N.; Bravo, R.F.; Aleckovic-Halilovic, M.; Zou, H.; et al. Chronic kidney disease and cardiovascular risk in six regions of the world (ISN-KDDC): A cross-sectional study. Lancet Glob. Health 2016, 4, e307–e319. [Google Scholar] [CrossRef]
- Stanifer, J.W.; Jing, B.; Tolan, S.; Helmke, N.; Mukerjee, R.; Naicker, S.; Patel, U. The epidemiology of chronic kidney disease in sub-Saharan Africa: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e174–e181. [Google Scholar] [CrossRef]
- Sumaili, E.K.; Krzesinski, J.-M.; Cohen, E.P.; Nseka, N.M. Epidemiology of chronic kidney disease in the Democratic Republic of Congo: Review of cross-sectional studies from Kinshasa, the capital. Nephrol. Ther. 2010, 6, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.; Bergstrom, J.; Dzurik, R.; Gulyassy, P.; Lockwood, D.; Phillips, L. Toxins and inhibitors in chronic renal failure. Am. J. Kidney Dis. 1986, 7, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Bergwitz, C.; Jüppner, H. Regulation of phosphate homeostasis by PTH, Vitamin D, and FGF23. Annu. Rev. Med. 2010, 61, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Fevrier-Paul, A.; Soyibo, A.K.; Mitchell, S.; Voutchkov, M. Role of toxic elements in chronic kidney disease. J. Health Pollut. 2018, 8, 181202. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, S.R.; Aeddula, N.R. Chronic renal failure. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Vadakedath, S.; Kandi, V. Dialysis: A review of the mechanisms underlying complications in the management of chronic renal failure. Cureus 2017, 9, e1603. [Google Scholar] [CrossRef]
- Smart, N.A.; Titus, T.T. Outcomes of early versus late nephrology referral in chronic kidney disease: A systematic review. Am. J. Med. 2011, 124, 1073–1080.e2. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, A.; Ferguson, T.W.; Rigatto, C.; Tangri, N.; Dumanski, S.; Komenda, P. Cost of dialysis therapy by modality in Manitoba. Clin. J. Am. Soc. Nephrol. 2018, 13, 1197–1203. [Google Scholar] [CrossRef]
- Halle, M.P.E.; Kengne, A.P.; Ashuntantang, A. Referral of patients with kidney impairment for specialist care in a developing country of sub-Saharan Africa. Ren. Fail. 2009, 31, 341–348. [Google Scholar] [CrossRef]
- Liyanage, T.; Ninomiya, T.; Jha, V.; Neal, B.; Patrice, H.M.; Okpechi, I.; Zhao, M.; Lv, J.; Garg, A.X.; Knight, J.; et al. Worldwide access to treatment for end-stage kidney disease: A systematic review. Lancet 2015, 385, 1975–1982. [Google Scholar] [CrossRef]
- Lisowska-Myjak, B. Uremic toxins and their effects on multiple organ systems. Nephron. Clin. Pract. 2014, 128, 303–311. [Google Scholar] [CrossRef]
- Nataatmadja, M.; Cho, Y.; Campbell, K.; Johnson, D.W. The roles of indoxyl sulphate and p-cresyl sulphate in patients with chronic kidney disease: A review of therapeutic options. In Chronic Kidney Disease—From Pathophysiology to Clinical Improvements; IntechOpen: London, UK, 2017. [Google Scholar]
- Flythe, J.E.; Hostetter, T.H. Assessing clinical relevance of uremic toxins. Clin. J. Am. Soc. Nephrol. 2019, 14, 182–183. [Google Scholar] [CrossRef] [PubMed]
- Lakkis, J.I.; Weir, M.R. Chapter 31. Hematologic and infectious complications of chronic kidney disease. In Chronic Renal Disease, 2nd ed.; Kimmel, P.L., Rosenberg, M.E., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 477–502. [Google Scholar]
- Persico, A.M.; Napolioni, V. Urinary p-cresol in autism spectrum disorder. Neurotoxicol. Teratol. 2013, 36, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-Y.; Li, J.-R.; Wang, Y.-Y.; Lin, S.-Y.; Ou, Y.-C.; Lin, C.-J.; Wang, J.-D.; Liao, S.-L.; Chen, C.-J. p-Cresyl sulfate caused behavior disorders and neurodegeneration in mice with unilateral nephrectomy involving oxidative stress and neuroinflammation. Int. J. Mol. Sci. 2020, 21, 6687. [Google Scholar] [CrossRef]
- Stuivenberg, G.A.; Chmiel, J.A.; Akouris, P.P.; Burton, J.P.; Reid, G. Probiotic bifidobacteria mitigate the deleterious effects of para-cresol in a Drosophila melanogaster toxicity model. mSphere 2022, 7, e00446-22. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Macfarlane, G.T. Enumeration of human colonic bacteria producing phenolic and indolic compounds: Effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol. 1996, 81, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Macfarlane, G.T. Dissimilatory amino acid metabolism in human colonic bacteria. Anaerobe 1997, 3, 327–337. [Google Scholar] [CrossRef]
- Saito, Y.; Sato, T.; Nomoto, K.; Tsuji, H. Identification of phenol- and p-cresol-producing intestinal bacteria by using media supplemented with tyrosine and its metabolites. FEMS Microbiol. Ecol. 2018, 94, fiy125. [Google Scholar] [CrossRef]
- Ramakrishna, B.S.; Gee, D.; Weiss, A.; Pannall, P.; Roberts-Thomson, I.C.; Roediger, W.E. Estimation of phenolic conjugation by colonic mucosa. J. Clin. Pathol. 1989, 42, 620–623. [Google Scholar] [CrossRef]
- Schepers, E.; Glorieux, G.; Vanholder, R. The gut: The forgotten organ in uremia? Blood Purif. 2010, 29, 130–136. [Google Scholar] [CrossRef]
- Meyer, T.W.; Hostetter, T.H. Uremic solutes from colon microbes. Kidney Int. 2012, 81, 949–954. [Google Scholar] [CrossRef]
- Lim, Y.J.; Sidor, N.A.; Tonial, N.C.; Che, A.; Urquhart, B.L. Uremic toxins in the progression of chronic kidney disease and cardiovascular disease: Mechanisms and therapeutic targets. Toxins 2021, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Salgado, J.; Vehaskari, V.M.; Stewart, T.; Ferris, M.; Zhang, Q.; Wang, G.; Blanchard, E.E.; Taylor, C.M.; Kallash, M.; Greenbaum, L.A.; et al. Intestinal microbiota in pediatric patients with end stage renal disease: A midwest pediatric nephrology consortium study. Microbiome 2016, 4, 50. [Google Scholar] [CrossRef] [PubMed]
- Stanford, J.; Charlton, K.; Stefoska-Needham, A.; Ibrahim, R.; Lambert, K. The gut microbiota profile of adults with kidney disease and kidney stones: A systematic review of the literature. BMC Nephrol. 2020, 21, 215. [Google Scholar] [CrossRef] [PubMed]
- Prasad, J.; Gill, H.; Smart, J.; Gopal, P.K. Selection and characterisation of Lactobacillus and Bifidobacterium strains for use as probiotics. Int. Dairy J. 1998, 8, 993–1002. [Google Scholar] [CrossRef]
- Reid, G. The scientific basis for probiotic strains of Lactobacillus. Appl. Environ. Microbiol. 1999, 65, 3763–3766. [Google Scholar] [CrossRef] [PubMed]
- Stuivenberg, G.; Daisley, B.; Akouris, P.; Reid, G. In vitro assessment of histamine and lactate production by a multi-strain synbiotic. J. Food Sci. Technol. 2022, 59, 3419–3427. [Google Scholar] [CrossRef] [PubMed]
- Ogata, T.; Nakamura, T.; Anjitsu, K.; Yaeshima, T.; Takahashi, S.; Fukuwatar, Y.; Ishibashi, N.; Hayasawa, H.; Fujisawa, T.; Iino, H. Effect of Bifidobacterium longum BB-536 yogurt administration on the intestinal environment of healthy adults. Microb. Ecol. Health Dis. 1999, 11, 41–46. [Google Scholar]
- Iwashita, Y.; Ohya, M.; Yashiro, M.; Sonou, T.; Kawakami, K.; Nakashima, Y.; Yano, T.; Iwashita, Y.; Mima, T.; Negi, S.; et al. Dietary changes involving Bifidobacterium longum and other nutrients delays chronic kidney disease progression. Am. J. Nurs. 2018, 47, 325–332. [Google Scholar] [CrossRef]
- Zirker, L. Probiotic use in chronic kidney disease patients. J. Ren. Nutr. 2014, 24, e47–e49. [Google Scholar] [CrossRef]
- Favero, C.; Ortiz, A.; Sanchez-Niño, M.D. Probiotics for kidney disease. Clin. Kidney J. 2022, 15, 1981–1986. [Google Scholar] [CrossRef]
- Wagner, S.; Merkling, T.; Metzger, M.; Koppe, L.; Laville, M.; Boutron-Ruault, M.-C.; Frimat, L.; Combe, C.; Massy, Z.A.; Stengel, B.; et al. Probiotic intake and inflammation in patients with chronic kidney disease: An analysis of the CKD-REIN cohort. Front. Nutr. 2022, 9, 772596. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, R.A.B.; Soder, T.F.; Grokoski, K.C.; Benetti, F.; Mendes, R.H. Probiotics in the treatment of chronic kidney disease: A systematic review. J. Bras. Nefrol. 2018, 40, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kort, R.; Sybesma, W. Probiotics for every body. Trends Biotechnol. 2012, 30, 613–615. [Google Scholar] [CrossRef] [PubMed]
- Kort, R.; Westerik, N.; Mariela Serrano, L.; Douillard, F.P.; Gottstein, W.; Mukisa, I.M.; Tuijn, C.J.; Basten, L.; Hafkamp, B.; Meijer, W.C.; et al. A novel consortium of Lactobacillus rhamnosus and Streptococcus thermophilus for increased access to functional fermented foods. Microb. Cell Factories 2015, 14, 195. [Google Scholar] [CrossRef] [PubMed]
- Hummelen, R.; Changalucha, J.; Butamanya, N.L.; Koyama, T.E.; Cook, A.; Habbema, J.D.F.; Reid, G. Effect of 25 weeks probiotic supplementation on immune function of HIV patients. Gut Microbes 2011, 2, 80–85. [Google Scholar] [CrossRef]
- Anukam, K.C.; Osazuwa, E.O.; Osadolor, H.B.; Bruce, A.W.; Reid, G. Yogurt containing probiotic Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 helps resolve moderate diarrhea and increases CD4 count in HIV/AIDS patients. J. Clin. Gastroenterol. 2008, 42, 239–243. [Google Scholar] [CrossRef]
- Van Tienen, A.; Hullegie, Y.; Hummelen, R.; Hemsworth, J.; Changalucha, J.; Reid, G. Development of a locally sustainable functional food for people living with HIV in sub-Saharan Africa: Laboratory testing and sensory evaluation. Beneficial Microbes 2011, 2, 193–198. [Google Scholar] [CrossRef]
- Westerik, N.; Kort, R.; Sybesma, W.; Reid, G. Lactobacillus rhamnosus probiotic food as a tool for empowerment across the value chain in Africa. Front. Microbiol. 2018, 9, 1501. [Google Scholar] [CrossRef]
- Reid, G.; Kort, R.; Alvarez, S.; Bourdet-Sicard, R.; Benoit, V.; Cunningham, M.; Saulnier, D.M.; van Hylckama Vlieg, J.E.T.; Verstraelen, H.; Sybesma, W. Expanding the reach of probiotics through social enterprises. Benef. Microbes 2018, 9, 707–715. [Google Scholar] [CrossRef]
- Nebra, Y.; Blanch, A.R. A new selective medium for Bifidobacterium spp. Appl. Environ. Microbiol. 1999, 65, 5173–5176. [Google Scholar] [CrossRef]
- Lee, S.Y.; Vedamuthu, E.R.; Washam, C.J.; Reinbold, G.W. Agar medium for the differential enumeration of yogurt starter bacteria. J. Milk Food Technol. 1974, 37, 272–276. [Google Scholar] [CrossRef]
- Driessen, F.M.; Ubbels, J.; Stadhouders, J. Continuous manufacture of yogurt, optimal conditions and kinetics of the prefermentation process. Biotechnol. Bioeng. 1977, 19, 821–839. [Google Scholar] [CrossRef]
- Chmiel, J.A.; Carr, C.; Stuivenberg, G.A.; Venema, R.; Chanyi, R.M.; Al, K.F.; Giguere, D.; Say, H.; Akouris, P.P.; Domínguez Romero, S.A.; et al. New perspectives on an old grouping: The genomic and phenotypic variability of Oxalobacter formigenes and the implications for calcium oxalate stone prevention. Front. Microbiol. 2022, 13, 1011102. [Google Scholar] [CrossRef]
- Liu, Y.; Gibson, G.R.; Walton, G.E. A three-stage continuous culture approach to study the impact of probiotics, prebiotics and fat intake on faecal microbiota relevant to an over 60s population. JFF 2017, 32, 238–247. [Google Scholar] [CrossRef]
- King, R.A.; May, B.L.; Davies, D.A.; Bird, A.R. Measurement of phenol and p-cresol in urine and feces using vacuum microdistillation and high-performance liquid chromatography. Anal. Biochem. 2009, 384, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Stuivenberg, G.A.; Burton, J.P.; Bron, P.A.; Reid, G. Why are bifidobacteria important for infants? Microorganisms 2022, 10, 278. [Google Scholar] [CrossRef]
- Kelly, S.M.; Munoz-Munoz, J.; van Sinderen, D. Plant glycan metabolism by bifidobacteria. Front. Microbiol. 2021, 12, 609418. [Google Scholar] [CrossRef]
- Turroni, F.; Özcan, E.; Milani, C.; Mancabelli, L.; Viappiani, A.; van Sinderen, D.; Sela, D.A.; Ventura, M. Glycan cross-feeding activities between bifidobacteria under in vitro conditions. Front. Microbiol. 2015, 6, 1030. [Google Scholar] [CrossRef]
- Egan, M.; O’Connell Motherway, M.; Kilcoyne, M.; Kane, M.; Joshi, L.; Ventura, M.; van Sinderen, D. Cross-feeding by Bifidobacterium breve UCC2003 during co-cultivation with Bifidobacterium bifidum PRL2010 in a mucin-based medium. BMC Microbiol. 2014, 14, 282. [Google Scholar] [CrossRef]
- Bunesova, V.; Lacroix, C.; Schwab, C. Mucin cross-feeding of infant bifidobacteria and Eubacterium hallii. Microb. Ecol. 2018, 75, 228–238. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; de los Reyes-Gavilan, C.G. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 2015, 362, fnv176. [Google Scholar] [CrossRef] [PubMed]
- Boger, M.C.L.; Lammerts van Bueren, A.; Dijkhuizen, L. Cross-feeding among probiotic bacterial strains on prebiotic inulin involves the extracellular exo-inulinase of Lactobacillus paracasei strain W20. Appl. Environ. Microbiol. 2018, 84, e01539-18. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.C.; Duar, R.M.; Lin, X.; Perez-Munoz, M.E.; Tollenaar, S.; Oh, J.-H.; van Pijkeren, J.-P.; Li, F.; van Sinderen, D.; Gänzle, M.G.; et al. Ecological importance of cross-feeding of the intermediate metabolite 1,2-propanediol between bacterial gut symbionts. Appl. Environ. Microbiol. 2020, 86, e00190-20. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.; James, K.; Bottacini, F.; Van Sinderen, D. Biochemical analysis of cross-feeding behaviour between two common gut commensals when cultivated on plant-derived arabinogalactan. Microb. Biotechnol. 2020, 13, 1733–1747. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Granata, L.A.; Morr, C.V. Improved acid, flavor and volatile compound production in a high protein and fiber soymilk yogurt-like product. J. Food Sci. 1996, 61, 331–336. [Google Scholar] [CrossRef]
- Kang, S.-S.; Kim, M.K.; Kim, Y.-J. Comprehensive evaluation of microbiological and physicochemical properties of commercial drinking yogurts in Korea. Food Sci. Anim. Resour. 2019, 39, 820–830. [Google Scholar] [CrossRef]
- el-Daher, N.; Na’was, T.; al-Qaderi, S. The effect of the pH of various dairy products on the survival and growth of Brucella melitensis. Ann. Trop. Med. Parasitol. 1990, 84, 523–528. [Google Scholar] [CrossRef]
- Chandrapala, J.; McKinnon, I.; Augustin, M.A.; Udabage, P. The influence of milk composition on pH and calcium activity measured in situ during heat treatment of reconstituted skim milk. J. Dairy. Res. 2010, 77, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Melini, F.; Melini, V.; Luziatelli, F.; Ruzzi, M. Raw and heat-treated milk: From public health risks to nutritional quality. Beverages 2017, 3, 54. [Google Scholar] [CrossRef]
- Usta-Gorgun, B.; Yilmaz-Ersan, L. Short-chain fatty acids production by Bifidobacterium species in the presence of salep. Electron. J. Biotechnol. 2020, 47, 29–35. [Google Scholar] [CrossRef]
- Markowiak-Kopeć, P.; Śliżewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Elkot, W.F.; Ateteallah, A.H.; Al-Moalem, M.H.; Shahein, M.R.; Alblihed, M.A.; Abdo, W.; Elmahallawy, E.K. Functional, physicochemical, rheological, microbiological, and organoleptic properties of synbiotic ice cream produced from camel milk using black rice powder and Lactobacillus acidophilus LA-5. Fermentation 2022, 8, 187. [Google Scholar] [CrossRef]
- Glorieux, G.; Schepers, E.; Vanholder, R.C. Uremic toxins in chronic renal failure. Prilozi 2007, 28, 173–204. [Google Scholar]
- Schulman, G.; Vanholder, R.; Niwa, T. AST-120 for the management of progression of chronic kidney disease. Int. J. Nephrol. Renovasc. Dis. 2014, 7, 49–56. [Google Scholar] [CrossRef]
- Ogata, T.; Nakamura, T.; Anjitsu, K.; Yaeshima, T.; Takahashi, S.; Fukuwatari, Y.; Ishibashi, N.; Hayasawa, H.; Fujisawa, T.; Iino, H. Effect of Bifidobacterium longum BB536 administration on the intestinal environment, defecation frequency and fecal characteristics of human volunteers. Biosci. Microflora 1997, 16, 53–58. [Google Scholar] [CrossRef]
- Mogna, L.; Del Piano, M.; Deidda, F.; Nicola, S.; Soattini, L.; Debiaggi, R.; Sforza, F.; Strozzi, G.; Mogna, G. Assessment of the in vitro inhibitory activity of specific probiotic bacteria against different Escherichia coli strains. J. Clin. Gastroenterol. 2012, 46, S29. [Google Scholar] [CrossRef]
- Bordoni, A.; Amaretti, A.; Leonardi, A.; Boschetti, E.; Danesi, F.; Matteuzzi, D.; Roncaglia, L.; Raimondi, S.; Rossi, M. Cholesterol-lowering probiotics: In vitro selection and in vivo testing of bifidobacteria. Appl. Microbiol. Biotechnol. 2013, 97, 8273–8281. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Reddel, S.; Del Chierico, F.; Quagliariello, A.; Giancristoforo, S.; Vernocchi, P.; Russo, A.; Fiocchi, A.; Rossi, P.; Putignani, L.; El Hachem, M. Gut microbiota profile in children affected by atopic dermatitis and evaluation of intestinal persistence of a probiotic mixture. Sci. Rep. 2019, 9, 4996. [Google Scholar] [CrossRef] [PubMed]
- Navarro-López, V.; Ramírez-Boscá, A.; Ramón-Vidal, D.; Ruzafa-Costas, B.; Genovés-Martínez, S.; Chenoll-Cuadros, E.; Carrión-Gutiérrez, M.; Horga de la Parte, J.; Prieto-Merino, D.; Codoñer-Cortés, F.M. Effect of oral administration of a mixture of probiotic strains on SCORAD index and use of topical steroids in young patients with moderate atopic dermatitis: A randomized clinical trial. JAMA Dermatol. 2018, 154, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Mogna, L.; Pane, M.; Nicola, S.; Raiteri, E. Screening of different probiotic strains for their in vitro ability to metabolise oxalates: Any prospective use in humans? J. Clin. Gastroenterol. 2014, 48 (Suppl. S1), S91–S95. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.S.-Y.; Yanagisawa, N.; Hor, Y.-Y.; Lew, L.-C.; Ong, J.-S.; Chuah, L.-O.; Lee, Y.-Y.; Choi, S.-B.; Rashid, F.; Wahid, N.; et al. Bifidobacterium longum BB536 alleviated upper respiratory illnesses and modulated gut microbiota profiles in Malaysian pre-school children. Benef. Microbes 2018, 9, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Sugahara, H.; Yonezawa, S.; Yaeshima, T.; Iwatsuki, K.; Tanabe, S.; Tominaga, T.; Togashi, H.; Benno, Y.; Xiao, J. Effect of the oralintake of yogurt containing Bifidobacterium longum BB536 on the cell numbers of enterotoxigenic Bacteroides fragilis in microbiota. Anaerobe 2012, 18, 14–18. [Google Scholar] [CrossRef]
- Xiao, J.Z.; Kondo, S.; Yanagisawa, N.; Takahashi, N.; Odamaki, T.; Iwabuchi, N.; Iwatsuki, K.; Kokubo, S.; Togashi, H.; Enomoto, K.; et al. Effect of probiotic Bifidobacterium longum BB536 [Corrected] in relieving clinical symptoms and modulating plasma cytokine levels of Japanese cedar pollinosis during the pollen season. A randomized double-blind, placebo-controlled trial. J. Investi.g Allergol. Clin. Immunol. 2006, 16, 86–93. [Google Scholar]
- West, N.P.; Horn, P.L.; Pyne, D.B.; Gebski, V.J.; Lahtinen, S.J.; Fricker, P.A.; Cripps, A.W. Probiotic supplementation for respiratory and gastrointestinal illness symptoms in healthy physically active individuals. Clin. Nutr. 2014, 33, 581–587. [Google Scholar] [CrossRef]
- Turner, R.B.; Woodfolk, J.A.; Borish, L.; Steinke, J.W.; Patrie, J.T.; Muehling, L.M.; Lahtinen, S.; Lehtinen, M.J. Effect of probiotic on innate inflammatory response and viral shedding in experimental rhinovirus infection—A randomised controlled trial. Benef. Microbes 2017, 8, 207–215. [Google Scholar] [CrossRef]
- Drago, L.; De Vecchi, E.; Gabrieli, A.; De Grandi, R.; Toscano, M. Immunomodulatory effects of Lactobacillus salivarius LS01 and Bifidobacterium breve BR03, alone and in combination, on peripheral blood mononuclear cells of allergic asthmatics. Allergy Asthma Immunol. Res. 2015, 7, 409–413. [Google Scholar] [CrossRef]
- Magistrelli, L.; Amoruso, A.; Mogna, L.; Graziano, T.; Cantello, R.; Pane, M.; Comi, C. Probiotics may have beneficial effects in Parkinson’s disease: In vitro evidence. Front. Immunol. 2019, 10, 969. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; De Palma, G.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Bifidobacterium strains suppress in vitro the pro-inflammatory milieu triggered by the large intestinal microbiota of coeliac patients. J. Inflamm. 2008, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Kamanova, J.; Cinova, J.; Olivares, M.; Drasarova, H.; Tuckova, L.; Sanz, Y. Modulation of phenotypic and functional maturation of dendritic cells by intestinal bacteria and gliadin: Relevance for celiac disease. J. Leukoc. Biol. 2012, 92, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Laparra, M.; Sanz, Y. Influence of Bifidobacterium longum CECT 7347 and gliadin peptides on intestinal epithelial cell proteome. J. Agric. Food Chem. 2011, 59, 7666–7671. [Google Scholar] [CrossRef] [PubMed]
- Erb, R.W.; Eichner, C.A.; Wagner-Döbler, I.; Timmis, K.N. Bioprotection of microbial communities from toxic phenol mixtures by a genetically designed pseudomonad. Nat. Biotechnol. 1997, 15, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Fung, D.Y.; Lin, C.C.; Gailani, M.B. Effect of phenolic antioxidants on microbial Growth. Crit. Rev. Microbiol. 1985, 12, 153–183. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Libudzisz, Z. Influence of phenol, p-cresol and indole on growth and survival of intestinal lactic acid bacteria. Anaerobe 2006, 12, 80–84. [Google Scholar] [CrossRef]
- Vazquez-Gutierrez, P.; de Wouters, T.; Werder, J.; Chassard, C.; Lacroix, C. High iron-sequestrating bifidobacteria inhibit enteropathogen growth and adhesion to intestinal epithelial cells in vitro. Front. Microbiol. 2016, 7, 1480. [Google Scholar] [CrossRef]
- Aw, W.; Fukuda, S. Protective effects of bifidobacteria against enteropathogens. Microb. Biotechnol. 2019, 12, 1097–1100. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stuivenberg, G.A.; Chmiel, J.A.; Akouris, P.P.; White, J.; Wilcox, H.; Seney, S.; Burton, J.P.; Reid, G. Supplementing Yogurt with Probiotic Bifidobacteria to Counter Chronic Kidney Disease. Fermentation 2023, 9, 391. https://doi.org/10.3390/fermentation9040391
Stuivenberg GA, Chmiel JA, Akouris PP, White J, Wilcox H, Seney S, Burton JP, Reid G. Supplementing Yogurt with Probiotic Bifidobacteria to Counter Chronic Kidney Disease. Fermentation. 2023; 9(4):391. https://doi.org/10.3390/fermentation9040391
Chicago/Turabian StyleStuivenberg, Gerrit A., John A. Chmiel, Polycronis P. Akouris, Jessica White, Hannah Wilcox, Shannon Seney, Jeremy P. Burton, and Gregor Reid. 2023. "Supplementing Yogurt with Probiotic Bifidobacteria to Counter Chronic Kidney Disease" Fermentation 9, no. 4: 391. https://doi.org/10.3390/fermentation9040391
APA StyleStuivenberg, G. A., Chmiel, J. A., Akouris, P. P., White, J., Wilcox, H., Seney, S., Burton, J. P., & Reid, G. (2023). Supplementing Yogurt with Probiotic Bifidobacteria to Counter Chronic Kidney Disease. Fermentation, 9(4), 391. https://doi.org/10.3390/fermentation9040391