
Impact of Rumen Fluid Storage on In Vitro Feed Fermentation Characteristics

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Chemical Composition
2.3. Rumen Fluid Collection
2.4. Incubation
2.5. Variables Measured
2.6. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navarro-Villa, A.; O’brien, M.; López, S.; Boland, T.; O’kiely, P. Modifications of a gas production technique for assessing in vitro rumen methane production from feedstuffs. Anim. Feed Sci. Technol. 2011, 166, 163–174. [Google Scholar] [CrossRef]
- Danielsson, R.; Ramin, M.; Bertilsson, J.; Lund, P.; Huhtanen, P. Evaluation of a gas in vitro system for predicting methane production in vivo. J. Dairy. Sci. 2017, 100, 8881–8894. [Google Scholar] [CrossRef] [PubMed]
- Getachew, G.; Robinson, P.H.; DePeters, E.J.; Taylor, S.J. Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Anim. Feed Sci. Technol. 2004, 111, 57–71. [Google Scholar] [CrossRef]
- Chaudhry, A. Slaughtered cattle as a source of rumen fluid to evaluate supplements for in vitro degradation of grass nuts and barley straw. Open. Vet. J. 2008, 2, 16–22. [Google Scholar] [CrossRef]
- Denek, N.; Can, A.; Avci, M. Frozen rumen fluid as microbial inoculum in the two-stage in vitro digestibility assay of ruminant feeds. S. Afr. J. Anim. Sci. 2010, 40, 251–256. [Google Scholar] [CrossRef]
- Luchini, N.; Broderick, G.; Combs, D. Preservation of ruminal microorganisms for in vitro determination of ruminal protein degradation. J. Anim. Sci. 1996, 74, 1134–1143. [Google Scholar] [CrossRef]
- Mould, F.; Kliem, K.; Morgan, R.; Mauricio, R. In vitro microbial inoculum: A review of its function and properties. Anim. Feed Sci. Technol. 2005, 123, 31–50. [Google Scholar] [CrossRef]
- Prates, A.; de Oliveira, J.; de la Fuente Oliver, G.; Abecia, L.; Fondevila, M. Validation of sheep rumen fluid frozen in liquid N as inoculum for in vitro gas production trials. In Options Méditerranéennes. Series A: Mediterranean Seminars; International Centre for Advanced Mediterranean Agronomic Studies: Paris, France, 2011; Volume 99, pp. 141–147. [Google Scholar]
- Belanche, A.; Palma-Hidalgo, J.M.; Nejjam, I.; Serrano, R.; Jimenez, E.; Martin-Garcia, I.; Yanez-Ruiz, D.R. In vitro assessment of the factors that determine the activity of the rumen microbiota for further applications as inoculum. J. Sci. Food Agric. 2018, 99, 163–172. [Google Scholar] [CrossRef]
- Chaudhry, A.; Mohamed, R. Fresh or frozen rumen contents from slaughtered cattle to estimate in vitro degradation of two contrasting feeds. Czech J. Anim. Sci. 2012, 57, 265–273. [Google Scholar] [CrossRef]
- Tanasawa, I. Things we do not know about cryopreservation of biological organs. Ann. N. Y. Acad. Sci. 1998, 858, 227–234. [Google Scholar] [CrossRef]
- Kisidayova, S.M.T.; Varadyova, Z. Preliminary results of the regeneration of the in vitro rumen ciliate cultures–Effect of their cytoresistance. Endocytobiosis Cell. Res. 2006, 17, 146–149. [Google Scholar]
- Nsabimana, E.; Kišidayová, S.; Macheboeuf, D.; Newbold, C.; Jouany, J. Two-step freezing procedure for cryopreservation of rumen ciliates, an effective tool for creation of a frozen rumen protozoa bank. Appl. Environ. Microbiol. 2003, 69, 3826–3832. [Google Scholar] [CrossRef] [PubMed]
- Tunkala, B.Z.; Di Giacomo, K.; Hess, P.S.A.; Dunshea, F.R.; Leury, B.J. Rumen fluid preservation for in vitro gas production systems. Anim. Feed Sci. Technol. 2022, 292, 115405. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code for the Care and Use of Animals for Scientific Purposes; National Health and Medical Research Council: Canberra, Australia, 2013. [Google Scholar]
- Horwitz, W. Official methods of analysis of AOAC International. In Agricultural Chemicals, Contaminants, Drugs; Horwitz, W., Ed.; AOAC International: Gaithersburg, MD, USA, 1997; Volume I, p. 2010. [Google Scholar]
- SCA. Feeding Standards for Australian Livestock. Ruminants; Csiro Publishing: Melbourne, Australia, 1990. [Google Scholar]
- AFFIA. Australian Fodder Industry Association—Laboratory Methods Manual; AFFIA: Melbourn, Australia, 2014. [Google Scholar]
- Gonzalez-Rivas, P.; DiGiacomo, K.; Giraldo, P.; Leury, B.; Cottrell, J.; Dunshea, F. Reducing rumen starch fermentation of wheat with three percent sodium hydroxide has the potential to ameliorate the effect of heat stress in grain-fed wethers. J. Anim. Sci. 2017, 95, 5547–5562. [Google Scholar] [CrossRef]
- Belanche, A.; Doreau, M.; Edwards, J.E.; Moorby, J.M.; Pinloche, E.; Newbold, C.J. Shifts in the rumen microbiota due to the type of carbohydrate and level of protein ingested by dairy cattle are associated with changes in rumen fermentation. J. Nutr. 2012, 142, 1684–1692. [Google Scholar] [CrossRef]
- Marten, G.C.; Barnes, R.F. Prediction of energy digestibility of forages with in vitro rumen fermentation and fungal enzyme systems. In Standardization of Analytical Methodology for Feeds: Proceedings of a Workshop Held in Ottawa, Canada, 12–l4 March 1979, Ottawa, ON, Canada; IDRC: Ottawa, ON, Canada, 1979; 128p. [Google Scholar]
- ANKOM. ANKOM Gas Production System Operator’s Manual; ANKOM Technology: Macedon, NY, USA, 2018. [Google Scholar]
- Alvarez Hess, P.S.; Eckard, R.J.; Jacobs, J.L.; Hannah, M.C.; Moate, P.J. Comparison of five methods for the estimation of methane production from vented in vitro systems. J. Sci. Food Agric. 2019, 99, 109–116. [Google Scholar] [CrossRef]
- Park, K.R.; Park, C.S.; Kim, B.G. An enzyme complex increases in vitro dry matter digestibility of corn and wheat in pigs. Springerplus 2016, 5, 1–7. [Google Scholar] [CrossRef]
- Blümmel, M.; Makkar, H.; Becker, K. In vitro gas production: A technique revisited. J. Anim. Physiol. Anim. Nutr. 1997, 77, 24–34. [Google Scholar] [CrossRef]
- Jouany, J. Volatile fatty acid and alcohol determination in digestive contents, silage juices, bacterial cultures and anaerobic fermentor contents, in microbial fermentation, biological liquids, analytic method using gas-liquid chromatography. Sci. Des Aliment. 1982, 2, 131–144. [Google Scholar]
- Weatherburn, M. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- EndNote Team. EndNote, X9 ed. Philadelphia, USA, 2013.
- Hristov, A.N.; McAllister, T.A.; Xu, Z.; Newbold, C.J. Proteolytic activity in ruminal fluid from cattle fed two levels of barley grain: A comparison of three methods of determination. J. Sci. Food Agric. 2002, 82, 1886–1893. [Google Scholar] [CrossRef]
- Hervás, G.; Frutos, P.; Giráldez, F.J.; Mora, M.J.; Fernández, B.; Mantecón, Á.R. Effect of preservation on fermentative activity of rumen fluid inoculum for in vitro gas production techniques. Anim. Feed Sci. Technol. 2005, 123–124, 107–118. [Google Scholar] [CrossRef]
- Furchtenicht, J.E.; Broderick, G.A. Effect of inoculum preparation and dietary energy on microbial numbers and rumen protein degradation activity. J. Dairy. Sci. 1987, 70, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, R.M.; Owen, E.; Mould, F.L.; Givens, I.; Theodorou, M.K.; France, J.; Davies, D.R.; Dhanoa, M.S. Comparison of bovine rumen liquor and bovine faeces as inoculum for an in vitro gas production technique for evaluating forages. Anim. Feed Sci. Technol. 2001, 89, 33–48. [Google Scholar] [CrossRef]
- Elghandour, M.; Vázquez, J.; Salem, A.; Kholif, A.; Cipriano, M.; Camacho, L.; Márquez, O. In vitro gas and methane production of two mixed rations influenced by three different cultures of Saccharomyces cerevisiae. J. Appl. Anim. Res. 2017, 45, 389–395. [Google Scholar] [CrossRef]
- Ingraham, J.; Maaløe, O.; Neidhardt, F. Growth of cells and cultures. In Growth of the Bacterial Cell; Sinauer Associates: Boston, MA, USA, 1983. [Google Scholar]
- Lee, M.R.; Merry, R.J.; Davies, D.R.; Moorby, J.M.; Humphreys, M.O.; Theodorou, M.K.; MacRae, J.C.; Scollan, N.D. Effect of increasing availability of water-soluble carbohydrates on in vitro rumen fermentation. Anim. Feed Sci. Technol. 2003, 104, 59–70. [Google Scholar] [CrossRef]
- Cone, J.W.; van Gelder, A.H.; Visscher, G.J.; Oudshoorn, L. Influence of rumen fluid and substrate concentration on fermentation kinetics measured with a fully automated time related gas production apparatus. Anim. Feed Sci. Technol. 1996, 61, 113–128. [Google Scholar] [CrossRef]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef]
- Guyader, J.; Eugène, M.; Noziere, P.; Morgavi, D.; Doreau, M.; Martin, C. Influence of rumen protozoa on methane emission in ruminants: A meta-analysis approach1. Animal 2014, 8, 1816–1825. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.; Hassan, H.; Abd-El-Raof, Y.; Abou-Zeina, H.A.; Galbt, S. Trails for cryopreservation of rumen protozoa in sheep. Glob. Vet. 2007, 1, 9–16. [Google Scholar]
- Harikrishna, C.; Mahender, M.; Reddy, Y.R.; Prakash, M.G.; Sudhakar, K.; Pavani, M. Evaluation of in vitro gas production and nutrient digestibility of complete diets supplemented with different levels of thermotolerant yeast in Nellore rams. Vet. World 2012, 5, 477–485. [Google Scholar] [CrossRef]
- Menke, K.H.; Stengass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Rymer, C.; Huntington, J.A.; Williams, B.A.; Givens, D.I. In vitro cumulative gas production techniques: History, methodological considerations and challenges. Anim. Feed Sci. Technol. 2005, 123–124, 9–30. [Google Scholar] [CrossRef]
- Meale, S.; Chaves, A.; Baah, J.; McAllister, T. Methane production of different forages in in vitro ruminal fermentation. Asian-Australas. J. Anim. Sci. 2012, 25, 86. [Google Scholar] [CrossRef]
- Kim, S.H.; Mamuad, L.L.; Jeong, C.D.; Choi, Y.J.; Lee, S.S.; Ko, J.Y.; Lee, S.S. In vitro Evaluation of Different Feeds for Their Potential to Generate Methane and Change Methanogen Diversity. Asian-Australas. J. Anim. Sci. 2013, 26, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
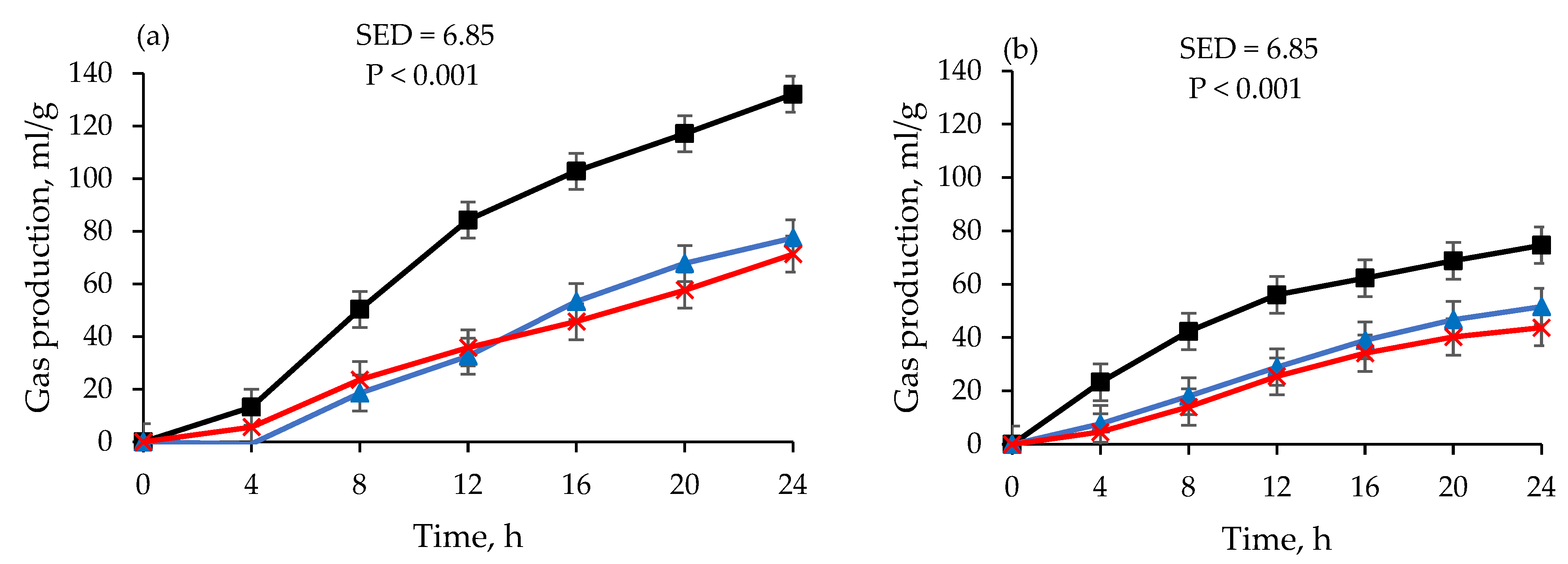
 ) and rumen fluid snap-frozen using liquid nitrogen and stored at −80 °C (−80 °C) for 14 days (
) and rumen fluid snap-frozen using liquid nitrogen and stored at −80 °C (−80 °C) for 14 days ( ) and 30 days (
) and 30 days ( ) at varying incubation times. Data at day 1 are for fresh RF. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
) and rumen fluid snap-frozen using liquid nitrogen and stored at −80 °C (−80 °C) for 14 days () and 30 days () at varying incubation times. Data at day 1 are for fresh RF. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
) at varying incubation times. Data at day 1 are for fresh RF. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
) and rumen fluid snap-frozen using liquid nitrogen and stored at −80 °C (−80 °C) for 14 days () and 30 days () at varying incubation times. Data at day 1 are for fresh RF. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
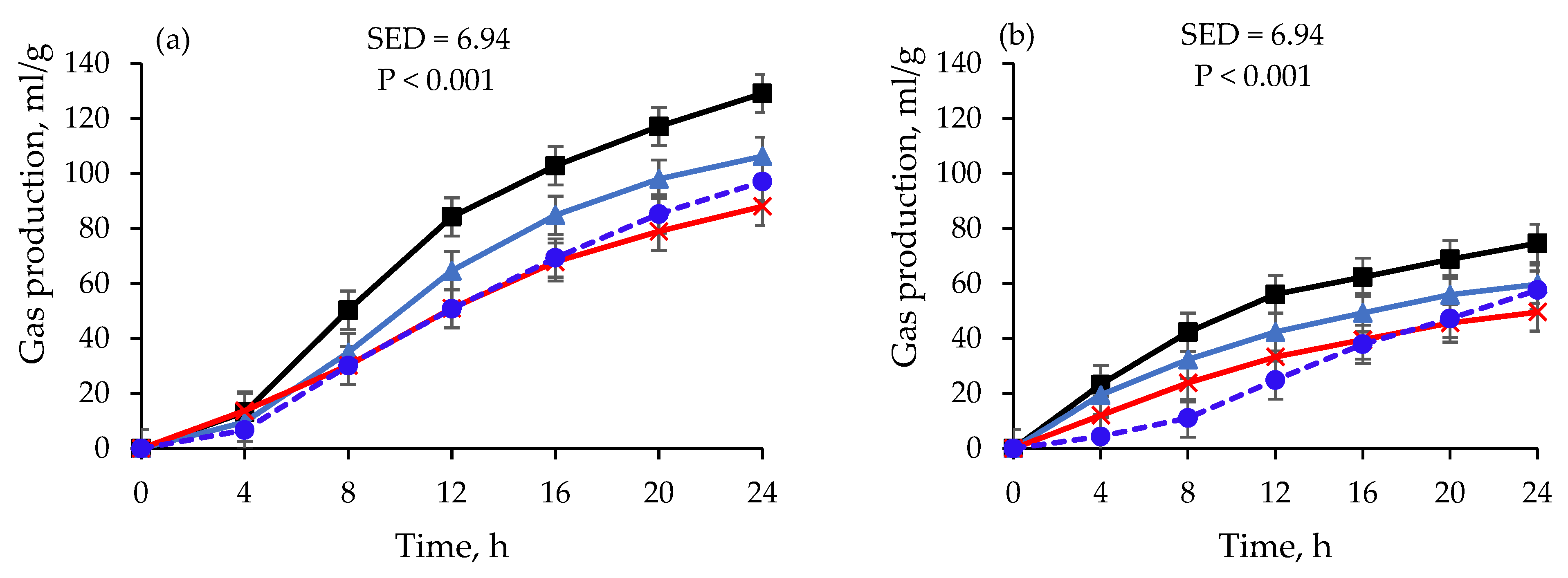
 ) and rumen fluid treated with DMSO and kept at D−20 °C for 14 days (
) and rumen fluid treated with DMSO and kept at D−20 °C for 14 days ( ), 30 days (
), 30 days ( ), and 180 days (
), and 180 days ( ) at varying incubation times. Data at day 1 are for fresh rumen fluid. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
) and rumen fluid treated with DMSO and kept at D−20 °C for 14 days (), 30 days (), and 180 days () at varying incubation times. Data at day 1 are for fresh rumen fluid. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
) at varying incubation times. Data at day 1 are for fresh rumen fluid. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.
) and rumen fluid treated with DMSO and kept at D−20 °C for 14 days (), 30 days (), and 180 days () at varying incubation times. Data at day 1 are for fresh rumen fluid. Data are least-square means of gas production after converting gas pressure into gas volume using Avogadro’s law.


{kind=link}
{kind=link}
{kind=link}
| Parameter | Lucerne Hay | Wheat Grain |
|---|---|---|
| Dry Matter | 95.2 | 90.3 |
| Moisture | 4.8 | 9.7 |
| Crude Protein | 22.5 | 13.5 |
| ADF | 24.6 | 4.9 |
| NDF | 39.9 | 11.1 |
| Fat | 4.1 | 1.5 |
| Ash | 9.9 | <1.0 |
| NFC | 23.7 | 75.9 |
| Dry matter digestibility | 65.1 | 94.3 |
| Organic matter digestibility 1 | 62.0 | 92.7 |
| ME 1 | 9.60 | 14.2 |
| Parameter | Wheat Grain | Lucerne Hay | SED 2 | S | D | S × D | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 14 | 30 | 1 | 14 | 30 | |||||
| Gas production, mL/g | 132.9 | 78.2 | 71.3 | 71.6 | 51.7 | 44.8 | 6.85 | <0.001 | <0.001 | 0.065 |
| Lag time, h | 0.75 | 5.26 | 5.20 | 0.45 | 1.37 | 1.39 | 0.195 | <0.001 | <0.001 | <0.001 |
| Gas prod. rate, mL/h | 5.39 | 3.26 | 2.97 | 3.01 | 2.12 | 1.83 | 0.293 | <0.001 | <0.001 | 0.018 |
| pH | 5.48 | 5.61 | 5.67 | 6.11 | 6.23 | 6.28 | 0.009 | <0.001 | <0.001 | 0.052 |
| DMD, % | 58.6 | 53.9 | 53.1 | 56.0 | 49.1 | 48.5 | 1.63 | <0.001 | <0.001 | 0.51 |
| Methane, % gas | 8.79 | 3.62 | 3.03 | 6.33 | 3.79 | 2.75 | 1.980 | 0.42 | 0.003 | 0.56 |
| CO2, % gas | 91.2 | 96.4 | 97.0 | 93.7 | 96.2 | 97.3 | 2.03 | 0.32 | 0.007 | 0.77 |
| Partition factor, mg/mL | 4.80 | 7.22 | 7.47 | 8.04 | 9.56 | 11.3 | 0.660 | <0.001 | <0.001 | 0.54 |
| Parameter | Wheat Grain | Lucerne | SED 2 | S | D | S × D | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 14 | 30 | 180 | 1 | 14 | 30 | 180 | |||||
| Gas production, mL/g | 132.9 | 88.3 | 106.3 | 97.3 | 71.6 | 60.3 | 54.9 | 57.5 | 6.94 | <0.001 | <0.001 | 0.015 |
| Lag time, h | 0.73 | 2.04 | 3.69 | 3.90 | 0.45 | 0.52 | 0.31 | 0.80 | 0.699 | <0.001 | <0.001 | <0.001 |
| Gas prod. rate, mL/h | 5.38 | 3.69 | 4.43 | 4.10 | 3.00 | 2.53 | 2.08 | 2.41 | 0.269 | <0.001 | <0.001 | 0.023 |
| pH | 5.48 | 5.61 | 5.59 | 5.58 | 6.11 | 6.22 | 6.25 | 6.28 | 0.092 | <0.001 | <0.001 | 0.64 |
| DMD, % | 58.6 | 57.3 | 55.1 | 50.4 | 56.0 | 52.0 | 52.0 | 48.8 | 1.53 | <0.001 | <0.001 | 0.58 |
| Methane, % gas | 8.79 | 3.72 | 3.13 | 0.66 | 6.33 | 1.83 | 2.62 | 0.42 | 0.337 | <0.001 | <0.001 | 0.002 |
| Carbon dioxide, % gas | 91.2 | 96.3 | 96.9 | 99.3 | 93.7 | 98.2 | 97.4 | 99.6 | 0.34 | <0.001 | <0.001 | 0.002 |
| Partition factor, mg/mL | 4.80 | 6.85 | 5.20 | 5.14 | 8.04 | 8.87 | 10.5 | 8.48 | 0.581 | <0.001 | <0.001 | 0.001 |
| Ammonia-N, mL/g | 0.29 | 0.26 | 0.25 | 0.22 | 0.95 | 0.69 | 0.31 | 0.29 | 0.016 | <0.001 | <0.001 | 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tunkala, B.Z.; DiGiacomo, K.; Alvarez Hess, P.S.; Dunshea, F.R.; Leury, B.J. Impact of Rumen Fluid Storage on In Vitro Feed Fermentation Characteristics. Fermentation 2023, 9, 392. https://doi.org/10.3390/fermentation9040392
Tunkala BZ, DiGiacomo K, Alvarez Hess PS, Dunshea FR, Leury BJ. Impact of Rumen Fluid Storage on In Vitro Feed Fermentation Characteristics. Fermentation. 2023; 9(4):392. https://doi.org/10.3390/fermentation9040392
Chicago/Turabian StyleTunkala, Bereket Zeleke, Kristy DiGiacomo, Pablo S. Alvarez Hess, Frank R. Dunshea, and Brian J. Leury. 2023. "Impact of Rumen Fluid Storage on In Vitro Feed Fermentation Characteristics" Fermentation 9, no. 4: 392. https://doi.org/10.3390/fermentation9040392
APA StyleTunkala, B. Z., DiGiacomo, K., Alvarez Hess, P. S., Dunshea, F. R., & Leury, B. J. (2023). Impact of Rumen Fluid Storage on In Vitro Feed Fermentation Characteristics. Fermentation, 9(4), 392. https://doi.org/10.3390/fermentation9040392