
Inhibitory Effects of Ammonia on Archaeal 16S rRNA Transcripts in Thermophilic Anaerobic Digester Sludge

Abstract
:1. Introduction
2. Materials and Methods
2.1. Operation of Sludge Anaerobic Incubation
2.2. Physicochemical Analyses
2.3. Archaeal 16S rRNA Gene Amplification and Reverse Transcription PCR
2.4. High-Throughput Sequencing and Data Analysis
2.5. Genes Quantification by qPCR
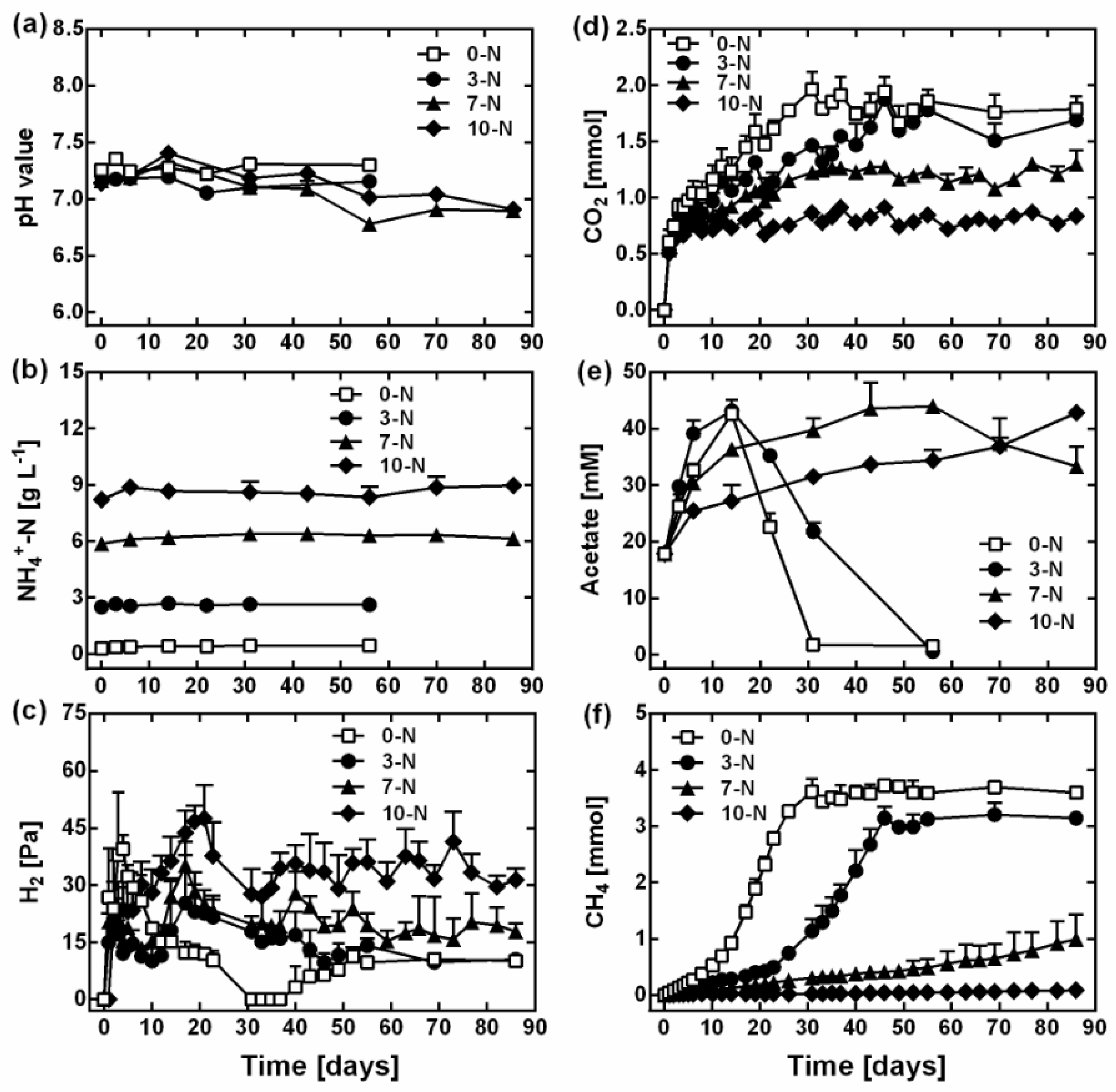
3. Results
3.1. Effect of Ammonia on Methanogenesis
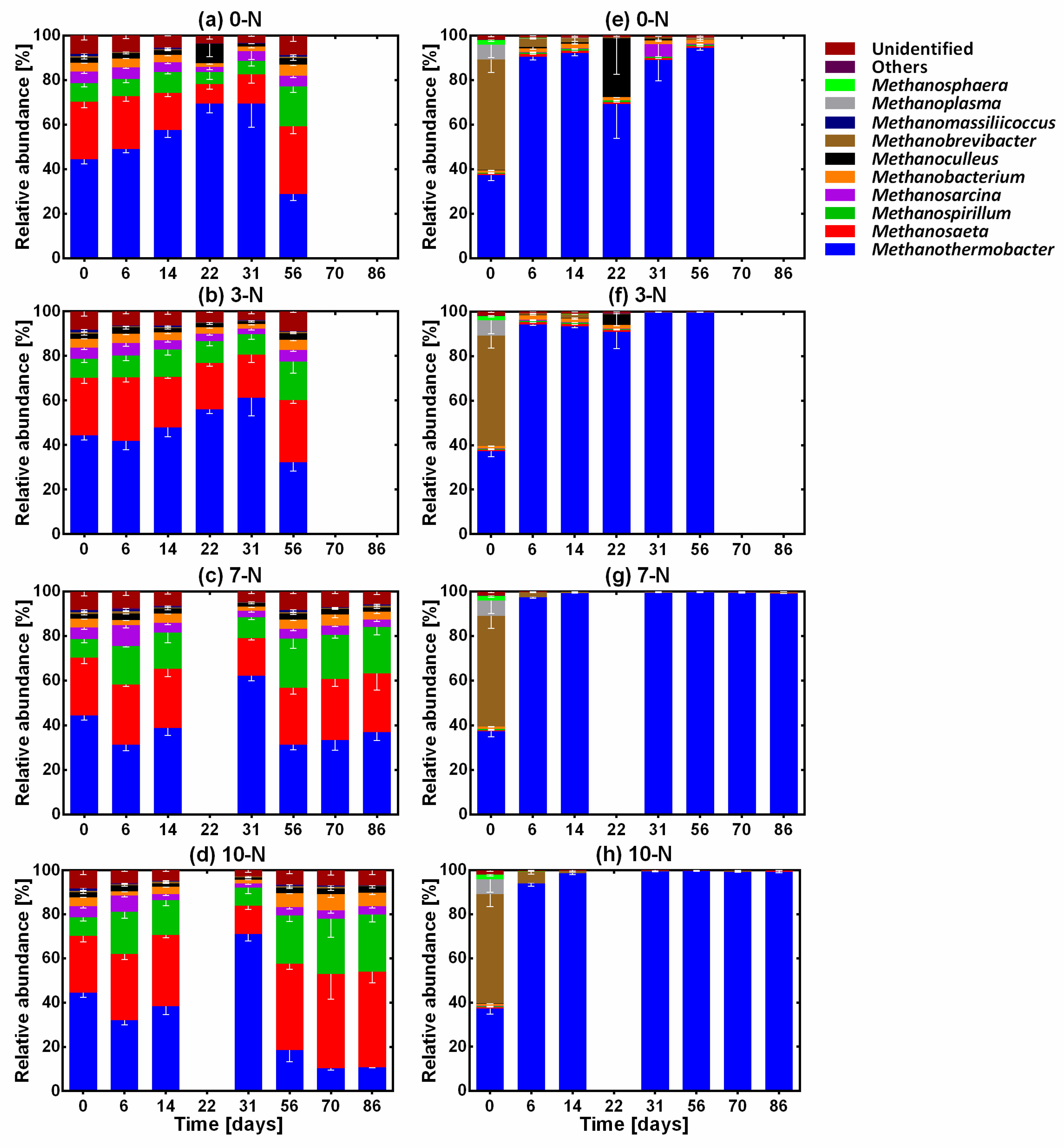
3.2. Methanogenic Archaea Dynamics
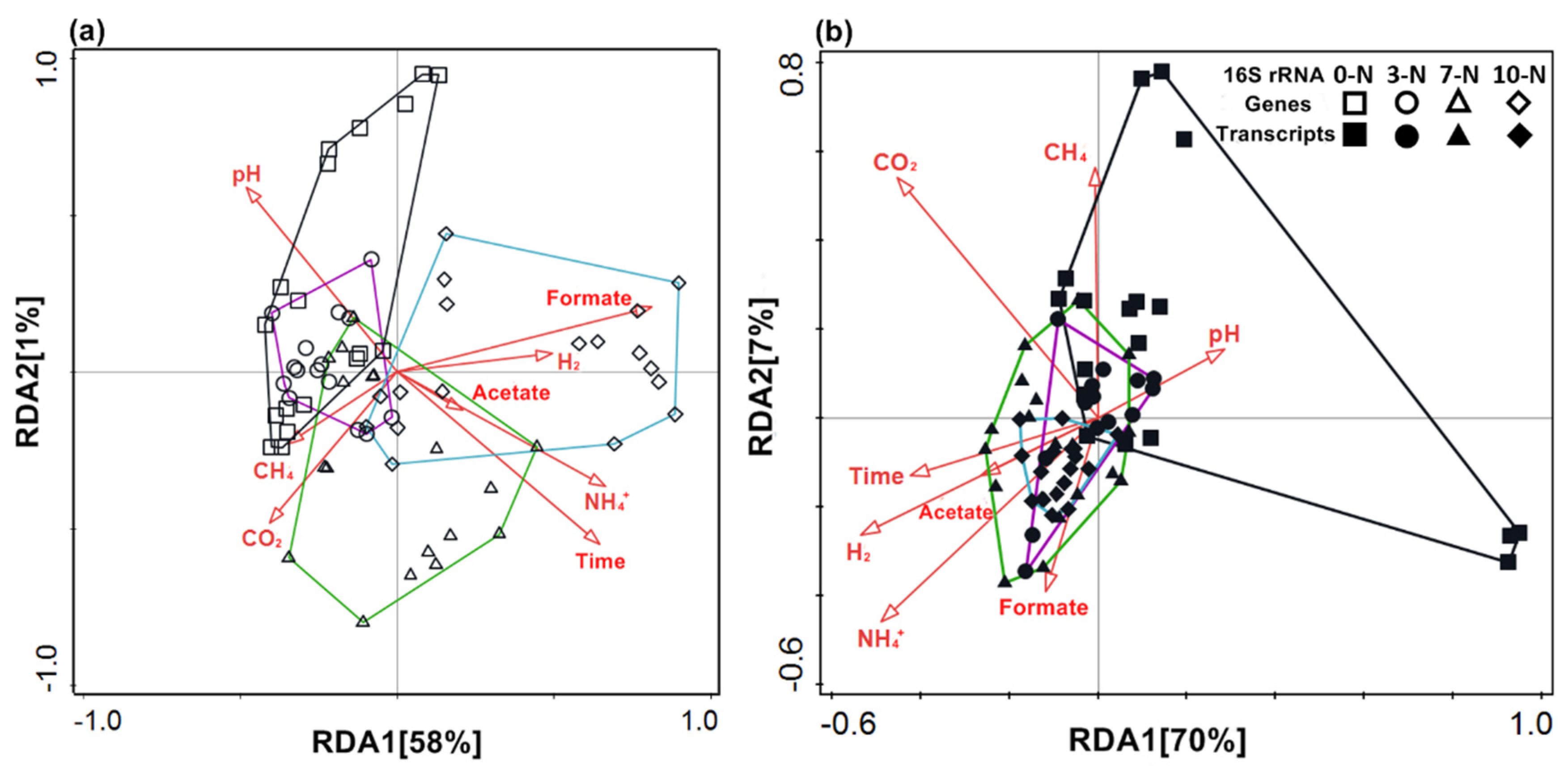
3.3. Correlation Analysis
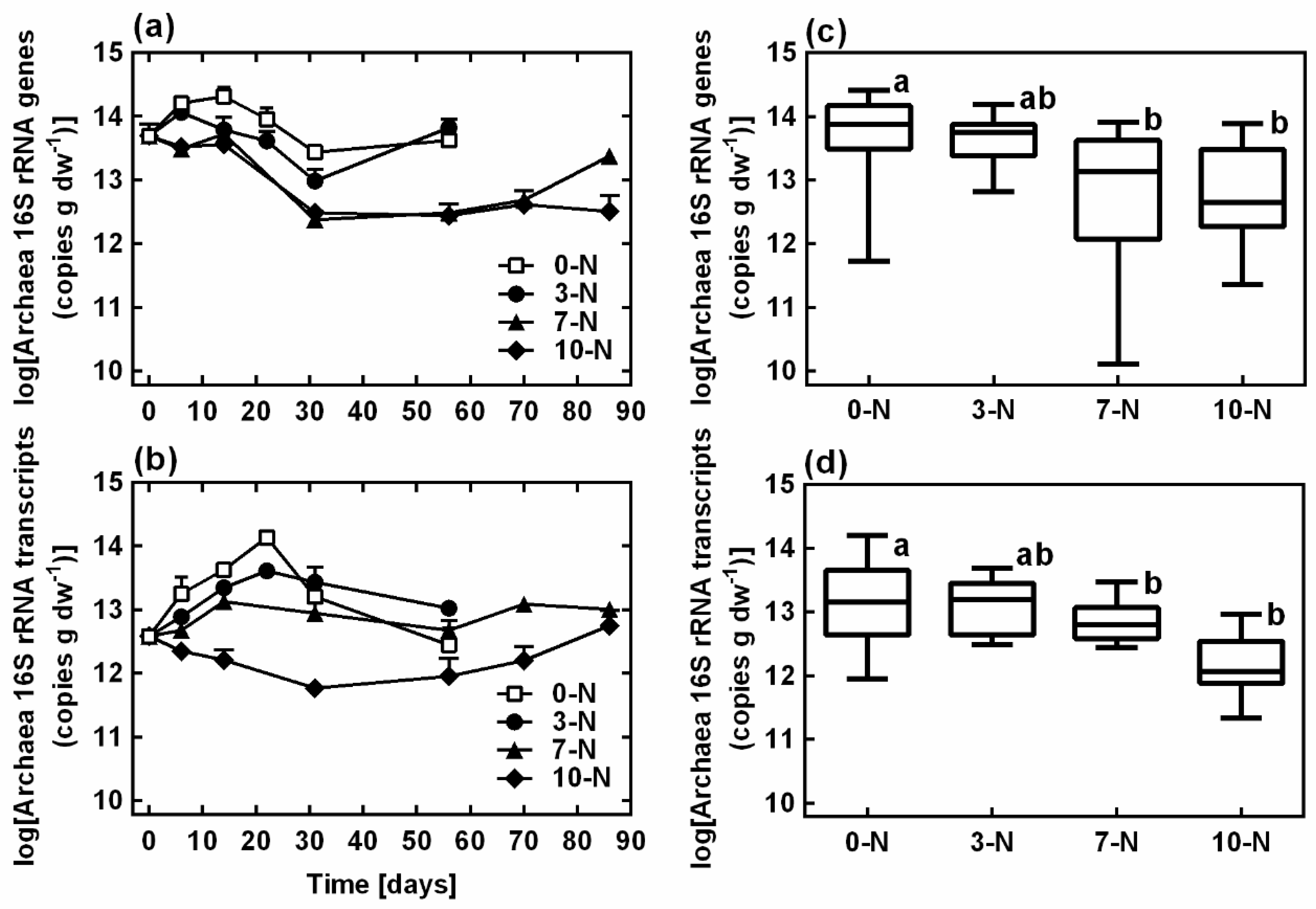
3.4. Archaeal 16S rRNA Gene and Transcript Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKendry, P. Energy production from biomass (Part 1): Overview of biomass. Bioresour. Technol. 2002, 83, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yuan, Q.; Lu, Y.H. Inhibitory effects of ammonia on syntrophic propionate oxidation in anaerobic digester sludge. Water Res. 2018, 146, 275–287. [Google Scholar] [CrossRef]
- Guo, X.H.; Wang, C.; Sun, F.Q.; Zhu, W.J.; Wu, W.X. A comparison of microbial characteristics between the thermophilic and mesophilic anaerobic digesters exposed to elevated food waste loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef]
- Shi, X.C.; Guo, X.L.; Zuo, J.N.; Wang, Y.J.; Zhang, M.Y. A comparative study of thermophilic and mesophilic anaerobic codigestion of food waste and wheat straw: Process stability and microbial community structure shifts. Waste Manag. 2018, 75, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, R.; Masse, D.I.; Singh, G. A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresour. Technol. 2013, 143, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Lin, Z.; Sun, Z.Y.; Gou, M.; Xia, Z.Y.; Tang, Y.Q. A comparative study of mesophilic and thermophilic anaerobic digestion of municipal sludge with high-solids content: Reactor performance and microbial community. Bioresour. Technol. 2020, 302, 122851. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.W.; He, P.J.; Zhang, H.; Shao, L.M.; Lu, F. Metabolic regulation of mesophilic Methanosarcina barkeri to ammonium inhibition. Environ. Sci. Technol. 2022, 56, 8897–8907. [Google Scholar] [CrossRef]
- Jo, Y.; Cayetano, R.D.A.; Kim, G.B.; Park, J.; Kim, S.H. The effects of ammonia acclimation on biogas recovery and the microbial population in continuous anaerobic digestion of swine manure. Environ. Res. 2022, 212, 113483. [Google Scholar] [CrossRef]
- Garcia, M.L.; Angenent, L.T. Interaction between temperature and ammonia in mesophilic digesters for animal waste treatment. Water Res. 2009, 43, 2373–2382. [Google Scholar] [CrossRef]
- Barber, W.P.F. Thermal hydrolysis for sewage treatment: A critical review. Water Res. 2016, 104, 53–71. [Google Scholar] [CrossRef]
- Yu, D.W.; Zhang, J.Y.; Chulu, B.H.; Yang, M.; Nopens, I.; Wei, Y.S. Ammonia stress decreased biomarker genes of acetoclastic methanogenesis and second peak of production rates during anaerobic digestion of swine manure. Bioresour. Technol. 2020, 317, 124012. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, D.T.N.; Suter, H.C.; Scales, P.J.; Chen, D.L. Lignite addition during anaerobic digestion of ammonium rich swine manure enhances biogas production. J. Environ. Chem. Eng. 2021, 9, 104669. [Google Scholar] [CrossRef]
- Wilson, C.A.; Novak, J.; Takacs, I.; Wett, B.; Murthy, S. The kinetics of process dependent ammonia inhibition of methanogenesis from acetic acid. Water Res. 2012, 46, 6247–6256. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Sasaki, K.; Watanabe, K.; Yumoto, I.; Kamagata, Y. Physiological and transcriptomic analyses of the thermophilic, aceticlastic methanogen Methanosaeta thermophila responding to ammonia stress. Microbes Environ. 2014, 29, 162–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerrillo, M.; Viñas, M.; Bonmatí, A. Overcoming organic and nitrogen overload in thermophilic anaerobic digestion of pig slurry by coupling a microbial electrolysis cell. Bioresour. Technol. 2016, 216, 362–372. [Google Scholar] [CrossRef]
- Yin, D.M.; Westerholm, M.; Qiao, W.; Bi, S.J.; Wandera, S.M.; Fan, R.; Jiang, M.M.; Dong, R.J. An explanation of the methanogenic pathway for methane production in anaerobic digestion of nitrogen-rich materials under mesophilic and thermophilic conditions. Bioresour. Technol. 2018, 264, 42–50. [Google Scholar] [CrossRef]
- Hagen, L.H.; Frank, J.A.; Zamanzadeh, M.; Eijsink, V.G.H.; Pope, P.B.; Horn, S.J.; Arntzen, M.Ø. Quantitative metaproteomics highlight the metabolic contributions of uncultured phylotypes in a thermophilic anaerobic digester. Appl. Environ. Microb. 2017, 83, e01955-16. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.P.; Leite, A.F.; Harms, H.; Glaser, K.; Liebetrau, J.; Kleinsteuber, S.; Nikolausz, M. Microbial community shifts in biogas reactors upon complete or partial ammonia inhibition. Appl. Microbiol. Biotechnol. 2019, 103, 519–533. [Google Scholar] [CrossRef]
- Lv, Z.P.; Hu, M.; Harms, H.; Richnow, H.H.; Liebetrau, J.; Nikolausz, M. Stable isotope composition of biogas allows early warning of complete process failure as a result of ammonia inhibition in anaerobic digesters. Bioresour. Technol. 2014, 167, 251–259. [Google Scholar] [CrossRef]
- Vigneron, A.; Cruaud, P.; Pignet, P.; Caprais, J.C.; Gayet, N.; Cambon-Bonavita, M.A.; Godfroy, A.; Toffin, L. Bacterial communities and syntrophic associations involved in anaerobic oxidation of methane process of the sonora margin cold seeps, guaymas basin. Environ. Microbiol. 2014, 16, 2777–2790. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.A.; Arntzen, M.Ø.; Sun, L.; Hagen, L.H.; Mchardy, A.C.; Horn, S.J. Novel syntrophic populations dominate an ammonia-tolerant methanogenic microbiome. Msystems 2016, 1, e00092-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yuan, Q.; Lu, Y.H. Inhibitory effects of ammonia on methanogen mcrA transcripts in anaerobic digester sludge. FEMS Microbiol. Ecol. 2014, 87, 368–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, J.P.; Peng, J.J.; Lu, Y.H. Succession of bacterial populations during plant residue decomposition in rice field soil. Appl. Environ. Microb. 2009, 75, 4879–4886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noll, M.; Matthies, D.; Frenzel, P.; Derakshani, M.; Liesack, W. Succession of bacterial community structure and diversity in a paddy soil oxygen gradient. Environ. Microbiol. 2005, 7, 382–395. [Google Scholar] [CrossRef]
- Ma, K.; Conrad, R.; Lu, Y. Responses of methanogen mcrA genes and their transcripts to an alternate dry/wet cycle of paddy field soil. Appl. Environ. Microb. 2012, 78, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Kemnitz, D.; Kolb, S.; Conrad, R. Phenotypic characterization of Rice Cluster III archaea without prior isolation by applying quantitative polymerase chain reaction to an enrichment culture. Environ. Microbiol. 2005, 7, 553–565. [Google Scholar] [CrossRef]
- Wang, H.; Fotidis, I.A.; Angelidaki, I. Ammonia effect on hydrogenotrophic methanogens and syntrophic acetate oxidizing bacteria. FEMS Microbiol. Ecol. 2015, 91, fiv130. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, Y.F.; Angelidaki, I. Ammonia inhibition on hydrogen enriched anaerobic digestion of manure under mesophilic and thermophilic conditions. Water Res. 2016, 105, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Sawayama, S.; Tada, C.; Tsukahara, K.; Yagishita, T. Effect of ammonium addition on methanogenic community in a fluidized bed anaerobic digestion. J. Biosci. Bioeng. 2004, 97, 65–70. [Google Scholar] [CrossRef]
- Schnurer, A.; Nordberg, A. Ammonia, a selective agent for methane production by syntrophic acetate oxidation at mesophilic temperature. Water Sci. Technol. 2008, 57, 735–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prathiviraj, R.; Chellapandi, P. Comparative genomic analysis reveals starvation survival systems in Methanothermobacter thermautotrophicus ΔH. Anaerobe 2020, 64, 102216. [Google Scholar] [CrossRef] [PubMed]
- Wasserfallen, A.; Nölling, J.; Pfister, P.; Reeve, J.; de Macario, E.C. Phylogenetic analysis of 18 thermophilic Methanobacterium isolates supports the proposals to create a new genus, Methanothermobacter gen. nov., and to reclassify several isolates in three species, Methanothermobacter thermautotrophicus comb. nov., Methanothermobacter wolfeii comb. nov., and Methanothermobacter marburgensis sp. nov. Int. J. Syst. Evol. Micr. 2000, 50 Pt 1, 43–53. [Google Scholar]
- Capson-Tojo, G.; Moscoviz, R.; Astals, S.; Robles, A.; Steyer, J.P. Unraveling the literature chaos around free ammonia inhibition in anaerobic digestion. Renew. Sust. Energy Rev. 2020, 117, 109487. [Google Scholar] [CrossRef]
- Yi, Y.; Dolfing, J.; Jin, G.; Fang, X.Y.; Han, W.H.; Liu, L.Y.; Tang, Y.Q.; Cheng, L. Thermodynamic restrictions determine ammonia tolerance of methanogenic pathways in Methanosarcina barkeri. Water Res. 2023, 232, 119664. [Google Scholar] [CrossRef]
- Cerrillo, M.; Viñas, M.; Bonmatí, A. Unravelling the active microbial community in a thermophilic anaerobic digester-microbial electrolysis cell coupled system under different conditions. Water Res. 2017, 110, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Ao, T.J.; Xie, Z.J.; Zhou, P.; Liu, X.F.; Wan, L.P.; Li, D. Comparison of microbial community structures between mesophilic and thermosphilic anaerobic digestion of vegetable waste. Bioproc. Biosyst. Eng. 2021, 44, 1201–1214. [Google Scholar] [CrossRef]
- Kato, S.; Kosaka, T.; Watanabe, K. Comparative transcriptome analysis of responses of Methanothermobacter thermautotrophicus to different environmental stimuli. Environ. Microbiol. 2008, 10, 893–905. [Google Scholar] [CrossRef]
- Munk, B.; Bauer, C.; Gronauer, A.; Lebuhn, M. A metabolic quotient for methanogenic archaea. Water Sci. Technol. 2012, 66, 2311–2317. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Conrad, R.; Lu, Y. Transcriptional response of methanogen mcrA genes to oxygen exposure of rice field soil. Environ. Microbiol. Rep. 2011, 3, 320–328. [Google Scholar] [CrossRef]
- Smith, K.S.; Ingram-Smith, C. Methanosaeta, the forgotten methanogen? Trends Microbiol. 2007, 15, 150–155. [Google Scholar] [CrossRef]
- Iino, T.; Mor, K.; Suzuki, K.I. Methanospirillum lacunae sp. nov., a methane-producing archaeon isolated from a puddly soil, and emended descriptions of the genus Methanospirillum and Methanospirillum hungatei. Int. J. Syst. Evol. Microbiol. 2010, 60 Pt 11, 2563–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misiukiewicz, A.; Gao, M.; Filipiak, W.; Cieslak, A.; Patra, A.K.; Szumacher-Strabel, M. Review: Methanogens and methane production in the digestive systems of nonruminant farm animals. Animal 2021, 15, 100060. [Google Scholar] [CrossRef] [PubMed]
- Karakashev, D.; Batstone, D.J.; Angelidaki, I. Influence of environmental conditions on methanogenic compositions in anaerobic biogas reactors. Appl. Environ. Microb. 2005, 71, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Steinhaus, B.; Garcia, M.L.; Shen, A.Q.; Angenent, L.T. A portable anaerobic microbioreactor reveals optimum growth conditions for the methanogen Methanosaeta concilii. Appl. Environ. Microb. 2007, 73, 1653–1658. [Google Scholar] [CrossRef] [Green Version]
- Nettmann, E.; Bergmann, I.; Pramschufer, S.; Mundt, K.; Plogsties, V.; Herrmann, C.; Klocke, M. Polyphasic analyses of methanogenic archaeal communities in agricultural biogas plants. Appl. Environ. Microb. 2010, 76, 2540–2548. [Google Scholar] [CrossRef] [Green Version]
- Sprott, G.D.; Patel, G.B. Ammonia toxicity in pure cultures of methanogenic bacteria. Syst. Appl. Microbiol. 1986, 7, 358–363. [Google Scholar] [CrossRef]
- Lu, Y.; Liaquat, R.; Astals, S.; Jensen, P.D.; Batstone, D.J.; Tait, S. Relationship between microbial community, operational factors and ammonia inhibition resilience in anaerobic digesters at low and moderate ammonia background concentrations. New Biotechnol. 2018, 44, 23–30. [Google Scholar] [CrossRef] [Green Version]
- De Vrieze, J.; Verstraete, W. Perspectives for microbial community composition in anaerobic digestion: From abundance and activity to connectivity. Environ. Microbiol. 2016, 18, 2797–2809. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Archaeal 16S rRNA Genes | Archaeal 16S rRNA Transcripts | |||||||
|---|---|---|---|---|---|---|---|---|
| Explanation [%] | Contribution [%] | Pseudo-F | p | Explanation [%] | Contribution [%] | Pseudo-F | p | |
| Formate | 38.3 | 63.9 | 41.6 | 0.002 ** | 0.2 | 0.3 | 0.6 | 0.528 |
| Time | 16.7 | 27.9 | 24.4 | 0.002 ** | 1.8 | 2.4 | 4.7 | 0.016 * |
| Acetate | 1.3 | 2.1 | 1.9 | 0.158 | 2.2 | 2.8 | 5.4 | 0.008 ** |
| CO2 | 1.8 | 3.1 | 2.8 | 0.086 | 31.3 | 40.0 | 43.0 | 0.002 ** |
| NH4+ | 1.0 | 1.7 | 1.5 | 0.21 | 8.1 | 10.4 | 18.4 | 0.002 ** |
| H2 | 0.4 | 0.6 | 0.6 | 0.426 | 20.5 | 26.2 | 17.3 | 0.002 ** |
| CH4 | 0.2 | 0.3 | 0.3 | 0.688 | 11.6 | 14.9 | 20.7 | 0.002 ** |
| pH | 0.2 | 0.3 | 0.3 | 0.69 | 2.3 | 3.0 | 6.5 | 0.002 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Song, C.; Li, J.; Zhang, C.; Li, P. Inhibitory Effects of Ammonia on Archaeal 16S rRNA Transcripts in Thermophilic Anaerobic Digester Sludge. Fermentation 2023, 9, 728. https://doi.org/10.3390/fermentation9080728
Wang S, Song C, Li J, Zhang C, Li P. Inhibitory Effects of Ammonia on Archaeal 16S rRNA Transcripts in Thermophilic Anaerobic Digester Sludge. Fermentation. 2023; 9(8):728. https://doi.org/10.3390/fermentation9080728
Chicago/Turabian StyleWang, Shenghui, Caihong Song, Jie Li, Chen Zhang, and Peng Li. 2023. "Inhibitory Effects of Ammonia on Archaeal 16S rRNA Transcripts in Thermophilic Anaerobic Digester Sludge" Fermentation 9, no. 8: 728. https://doi.org/10.3390/fermentation9080728
APA StyleWang, S., Song, C., Li, J., Zhang, C., & Li, P. (2023). Inhibitory Effects of Ammonia on Archaeal 16S rRNA Transcripts in Thermophilic Anaerobic Digester Sludge. Fermentation, 9(8), 728. https://doi.org/10.3390/fermentation9080728