

Optimization of an Efficient Direct Shoot Induction System and Histological Evaluation of Shoot Formation in Cucurbita maxima Duch

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Explant Preparation and Plantlet Regeneration
2.3. Media Composition and Preparation
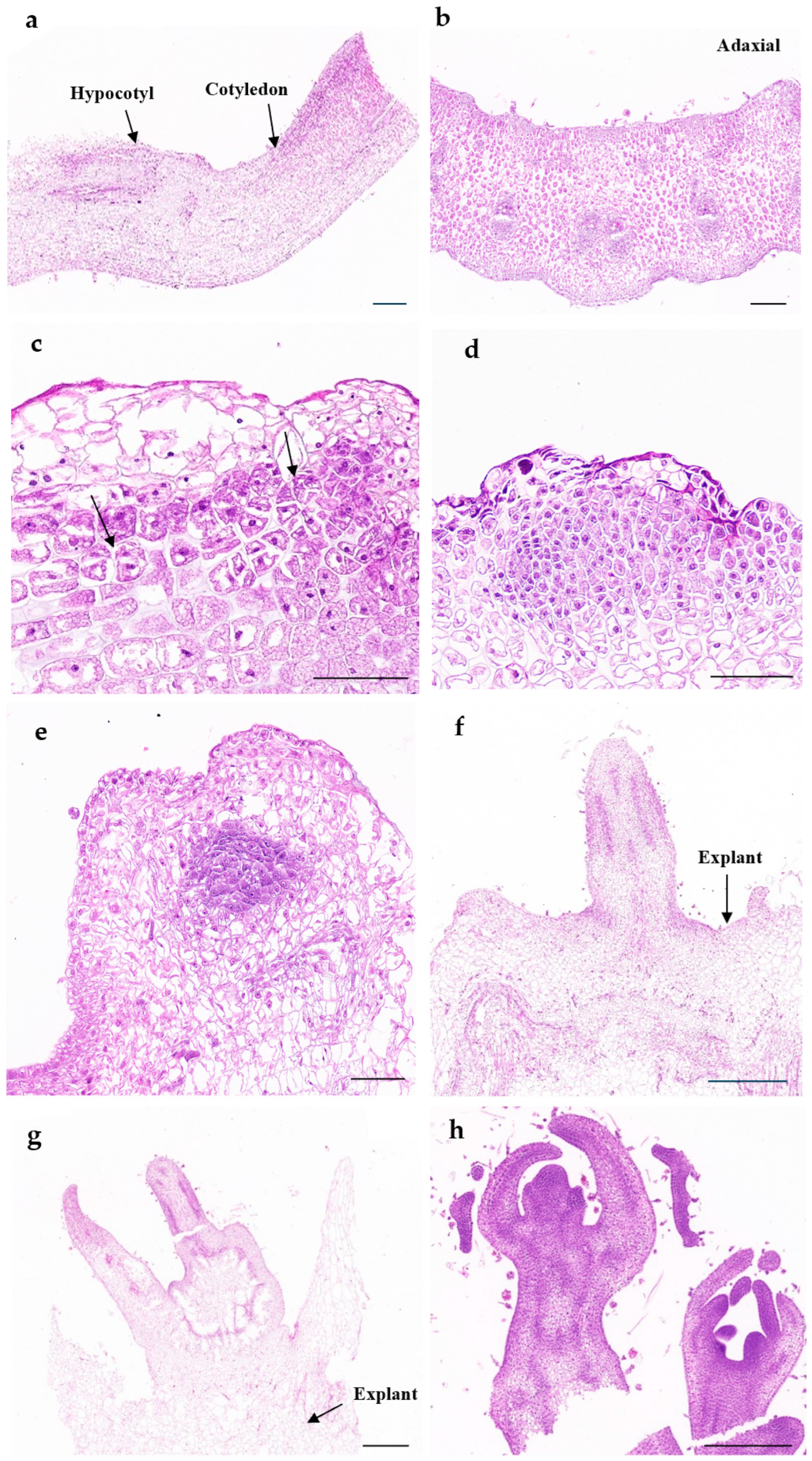
2.4. Histological Analysis
2.5. Propagation, Acclimatization, and Transplantation
2.6. Statistical Analysis
3. Results
3.1. Effects of Explant Type on Shoot Induction
3.2. Effects of Genotype on De Novo Shoot Regeneration
3.3. Effects of Media on De Novo Shoot Regeneration
3.4. Effects of Genotype and Induction Medium on De Novo Shoot Regeneration
3.5. Histological Observation of Adventitious Shoot Formation
3.6. Root Regeneration and Character Observation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miljić, M.; Rocchetti, G.; Krstić, S.; Mišan, A.; Brdar-Jokanović, M.; Marcheggiani, F.; Martinelli, E.; Lucini, L.; Damiani, E. Comparative In Vitro Antioxidant Capacity and Terpenoid Profiling of Pumpkin Fruit Pulps from a Serbian Cucurbita maxima and Cucurbita moschata Breeding Collection. Antioxidants 2021, 10, 1580. [Google Scholar] [CrossRef] [PubMed]
- El-Gendi, H.; Abu-Serie, M.M.; Kamoun, E.A.; Saleh, A.K.; El-Fakharany, E.M. Statistical Optimization and Characterization of Fucose-Rich Polysaccharides Extracted from Pumpkin (Cucurbita maxima) along with Antioxidant and Antiviral Activities. Int. J. Biol. Macromol. 2023, 232, 123372. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.M.K.; Huang, H.X.; Huang, W.J.; Xue, S.D.; Yan, S.J.; Wu, T.Q.; Li, J.X.; Zhong, Y.J. Evaluation of metabolites and antioxidant activity in pumpkin species. Nat. Prod. Commun. 2020, 15, 1934578X20920983. [Google Scholar] [CrossRef]
- Adams, G.G.; Imran, S.; Wang, S.; Mohammad, A.; Kok, M.S.; Gray, D.A.; Channell, G.A.; Harding, S.E. The Hypoglycemic Effect of Pumpkin Seeds, Trigonelline (TRG), Nicotinic Acid (NA), and D-Chiro-Inositol (DCI) in Controlling Glycemic Levels in Diabetes Mellitus. Crit. Rev. Food Sci. Nutr. 2014, 54, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.V.; Hoang, T.T.-H.; Le, N.T.; Tran, H.T.; Nguyen, C.X.; Moon, Y.-H.; Chu, H.H.; Do, P.T. An Efficient Hairy Root System for Validation of Plant Transformation Vector and CRISPR/Cas Construct Activities in Cucumber (Cucumis sativus L.). Front. Plant Sci. 2022, 12, 770062. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Li, D.; Liu, X.; Qi, J.; Gao, D.; Zhao, S.; Huang, S.; Sun, J.; Yang, L. Engineering Non-Transgenic Gynoecious Cucumber Using an Improved Transformation Protocol and Optimized CRISPR/Cas9 System. Mol. Plant 2017, 10, 1575–1578. [Google Scholar] [CrossRef]
- Baskaran, P.; Soós, V.; Balázs, E.; Van Staden, J. Shoot Apical Meristem Injection: A Novel and Efficient Method to Obtain Transformed Cucumber Plants. S. Afr. J. Bot. 2016, 103, 210–215. [Google Scholar] [CrossRef]
- Nanasato, Y.; Konagaya, K.; Okuzaki, A.; Tsuda, M.; Tabei, Y. Improvement of Agrobacterium-Mediated Transformation of Cucumber (Cucumis sativus L.) by Combination of Vacuum Infiltration and Co-Cultivation on Filter Paper Wicks. Plant Biotechnol. Rep. 2013, 7, 267–276. [Google Scholar] [CrossRef]
- Valls, M.P.; Lasa, J.M. Agrobacterium-Mediated Transformation of Commercial Melon (Cucumis Melo L., Cv. Amarillo Ore). Plant Cell Rep. 1994, 13, 145–148. [Google Scholar]
- Grumet, R. Agrobacterium Tumefaciens Mediated Transformation and Regeneration of Muskmelon Plants. Plant Cell Rep. 1990, 9, 160–164. [Google Scholar]
- Hooghvorst, I.; López-Cristoffanini, C.; Nogués, S. Efficient Knockout of Phytoene Desaturase Gene Using CRISPR/Cas9 in Melon. Sci. Rep. 2019, 9, 17077. [Google Scholar] [CrossRef]
- Akasaka-Kennedy, Y.; Tomita, K.; Ezura, H. Efficient Plant Regeneration and Agrobacterium-Mediated Transformation via Somatic Embryogenesis in Melon (Cucumis melo L.). Plant Sci. 2004, 166, 763–769. [Google Scholar] [CrossRef]
- Nizan, S.; Amitzur, A.; Dahan-Meir, T.; Benichou, J.I.C.; Bar-Ziv, A.; Perl-Treves, R. Mutagenesis of the Melon Prv Gene by CRISPR/Cas9 Breaks Papaya Ringspot Virus Resistance and Generates an Autoimmune Allele with Constitutive Defense Responses. J. Exp. Bot. 2023, 74, 4579–4596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Guo, S.; Tian, S.; Zhang, J.; Ren, Y.; Li, M.; Gong, G.; Zhang, H.; Xu, Y. CRISPR/Cas9-Mediated Mutagenesis of ClBG1 Decreased Seed Size and Promoted Seed Germination in Watermelon. Hortic. Res. 2021, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Jiang, L.; Gao, Q.; Zhang, J.; Zong, M.; Zhang, H.; Ren, Y.; Guo, S.; Gong, G.; Liu, F.; et al. Efficient CRISPR/Cas9-Based Gene Knockout in Watermelon. Plant Cell Rep. 2017, 36, 399–406. [Google Scholar] [CrossRef]
- Tian, S.; Jiang, L.; Cui, X.; Zhang, J.; Guo, S.; Li, M.; Zhang, H.; Ren, Y.; Gong, G.; Zong, M.; et al. Engineering Herbicide-Resistant Watermelon Variety through CRISPR/Cas9-Mediated Base-Editing. Plant Cell Rep. 2018, 37, 1353–1356. [Google Scholar] [CrossRef]
- Feng, Q.; Xiao, L.; He, Y.; Liu, M.; Wang, J.; Tian, S.; Zhang, X.; Yuan, L. Highly Efficient, Genotype-independent Transformation and Gene Editing in Watermelon (Citrullus Lanatus) Using a Chimeric ClGRF4-GIF1 Gene. Integr. Plant Biol. 2021, 63, 2038–2042. [Google Scholar] [CrossRef]
- Nanasato, Y.; Tabei, Y. A Method of Transformation and Current Progress in Transgenic Research on Cucumbers and Cucurbita Species. Plant Biotechnol. 2020, 37, 141–146. [Google Scholar] [CrossRef]
- Nanasato, Y.; Konagaya, K.; Okuzaki, A.; Tsuda, M.; Tabei, Y. Agrobacterium-Mediated Transformation of Kabocha Squash (Cucurbita Moschata Duch) Induced by Wounding with Aluminum Borate Whiskers. Plant Cell Rep. 2011, 30, 1455–1464. [Google Scholar] [CrossRef]
- Xin, T.; Tian, H.; Ma, Y.; Wang, S.; Yang, L.; Li, X.; Zhang, M.; Chen, C.; Wang, H.; Li, H.; et al. Targeted Creation of New Mutants with Compact Plant Architecture Using CRISPR/Cas9 Genome Editing by an Optimized Genetic Transformation Procedure in Cucurbit Plants. Hortic. Res. 2022, 9, uhab086. [Google Scholar] [CrossRef]
- Chen, X.; He, S.; Jiang, L.; Li, X.; Guo, W.; Chen, B.; Zhou, J.; Skliar, V. An Efficient Transient Transformation System for Gene Function Studies in Pumpkin (Cucurbita moschata D.). Sci. Hortic. 2021, 282, 110028. [Google Scholar] [CrossRef]
- Ananthakrishnan, G.; Xia, X.; Amutha, S.; Singer, S.; Muruganantham, M.; Yablonsky, S.; Fischer, E.; Gaba, V. Ultrasonic Treatment Stimulates Multiple Shoot Regeneration and Explant Enlargement in Recalcitrant Squash Cotyledon Explants in Vitro. Plant Cell Rep. 2007, 26, 267–276. [Google Scholar] [CrossRef]
- Jelaska, S. Embryogenesis and Organogenesis in Pumpkin Explants. Physiol. Plant 1974, 31, 257–261. [Google Scholar] [CrossRef]
- Zou, T.; Song, H.; Chu, X.; Tong, L.; Liang, S.; Gong, S.; Yang, H.; Sun, X. Efficient Induction of Gynogenesis through Unfertilized Ovary Culture with Winter Squash (Cucurbita maxima Duch.) and Pumpkin (Cucurbita moschata Duch.). Sci. Hortic. 2020, 264, 109152. [Google Scholar] [CrossRef]
- Gonsalves, C.; Xue, B.; Gonsalves, D. Somatic Embryogenesis and Regeneration from Cotyledon Explants of Six Squash Cultivars. HortScience 1995, 30, 1295–1297. [Google Scholar] [CrossRef]
- Lee, Y.K.; Chung, W.I.; Ezura, H. Efficient Plant Regeneration via Organogenesis in Winter Squash (Cucurbita maxima Duch.). Plant Sci. 2003, 164, 413–418. [Google Scholar] [CrossRef]
- Sugimoto, K.; Temman, H.; Kadokura, S.; Matsunaga, S. To Regenerate or Not to Regenerate: Factors That Drive Plant Regeneration. Curr. Opin. Plant Biol. 2019, 47, 138–150. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kim, K.-M.; Kim, C.K.; Han, J.-S. In Vitro Regeneration from Cotyledon Explants in Figleaf Gourd (Cucurbita ficifolia Bouché), a Rootstock for Cucurbitaceae. Plant Biotechnol. Rep. 2010, 4, 101–107. [Google Scholar] [CrossRef]
- Ananthakrishnan, G.; Xia, X.; Elman, C.; Singer, S.; Paris, H.S.; Gal-On, A.; Gaba, V. Shoot Production in Squash (Cucurbita Pepo) by in Vitro Organogenesis. Plant Cell Rep. 2003, 21, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Kathiravan, K.; Vengedesan, G.; Singer, S.; Steinitz, B.; Paris, H.S.; Gaba, V. Adventitious Regeneration in Vitro Occurs across a Wide Spectrum of Squash (Cucurbita pepo) Genotypes. Plant Cell Tiss. Organ Cult. 2006, 85, 285–295. [Google Scholar] [CrossRef]
- Raza, G.; Singh, M.B.; Bhalla, P.L. Somatic Embryogenesis and Plant Regeneration from Commercial Soybean Cultivars. Plants 2019, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.R.; Akhond, M.Y.; Amin, M.A.; Haque, M.S. In Vitro Regeneration of Sweetgourd (Cucurbita moschata Duch.). Plant Tissue Cult. Biotech. 2016, 26, 67–75. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jinu, U.; Sangeetha, P.; Geetha, N.; Sahi, S.V. High Frequency Plant Regeneration from Cotyledonary Node Explants of Cucumis sativus L. Cultivar ‘Green Long’ via Adventitious Shoot Organogenesis and Assessment of Genetic Fidelity by RAPD-PCR Technology. 3 Biotech 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, J.; Wu, T.; Cao, J. Shoot Regeneration and the Relationship between Organogenic Capacity and Endogenous Hormonal Contents in Pumpkin. Plant Cell Tiss. Organ Cult. 2008, 93, 323. [Google Scholar] [CrossRef]
- Samuel Aworunse, O.; Voke Omasoro, R.; Soneye, B.; Odun Obembe, O. Effect of Low BAP Levels on Multiple Shoots Induction in Indigenous Nigerian Pumpkin (Cucurbita pepo Linn.). J. Phys. Conf. Ser. 2019, 1299, 012100. [Google Scholar] [CrossRef]
- Zhang, H.; Peng, G.; Feishi, L. Efficient Plant Regeneration from Cotyledonary Node Explants of Cucumis melo L. Afr. J. Biotechnol. 2011, 10, 6757–6761. [Google Scholar]
- Ficcadenti, N.; Rotino, G.L. Genotype and Medium Affect Shoot Regeneration of Melon. Plant Cell Tiss. Organ Cult. 1995, 40, 293–295. [Google Scholar] [CrossRef]
- Ren, Y.; Bang, H.; Gould, J.; Rathore, K.S.; Patil, B.S.; Crosby, K.M. Shoot Regeneration and Ploidy Variation in Tissue Culture of Honeydew Melon (Cucumis melo L. Inodorus). In Vitro Cell. Dev. Biol.-Plant 2013, 49, 223–229. [Google Scholar] [CrossRef]
- Urbanek, A.; Zechmann, B.; Muller, M. Plant Regeneration via Somatic Embryogenesis in Styrian Pumpkin: Cytological and Biochemical Investigations. Plant Cell Tiss. Organ Cult. 2004, 79, 329–340. [Google Scholar] [CrossRef]
- Parvin, S.; Kausar, M.; Haque, M.E.; Khalekuzzaman, M.; Sikdar, B.; Islam, M.A. In Vitro Propagation of Muskmelon (Cucumis melo L.) from Nodal Segments, Shoot Tips and Cotyledonary Nodes. Rajshahi Univ. J. Life Earth Agric. Sci. 2015, 41, 71–77. [Google Scholar] [CrossRef]
- Kurtar, E.S.; Balkaya, A.; Ozbakir Ozer, M. Production of Callus Mediated Gynogenic Haploids in Winter Squash (Cucurbita maxima Duch.) and Pumpkin (Cucurbita moschata Duch.). Czech J. Genet. Plant Breed. 2018, 54, 9–16. [Google Scholar] [CrossRef]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, Efflux-Dependent Auxin Gradients as a Common Module for Plant Organ Formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | Cultivar | Seed Provider |
|---|---|---|
| L-1 | Hong-li No. 2 | Hunan Xue-Feng Seed Company, Ltd., Shaoyang, China |
| L-2 | Hong-Li 68 | Hunan Xue-Feng Seed Company, Ltd., Shaoyang, China |
| L-3 | Jin-Li | Hunan Xue-Feng Seed Company, Ltd., Shaoyang, China |
| L-4 | Qing 4 | Hunan Xue-Feng Seed Company, Ltd., Shaoyang, China |
| L-5 | JP | Hunan Xue-Feng Seed Company, Ltd., Shaoyang, China |
| Induction Culture | MS g/L | Sucrose g/L | Agar g/L | 6-Benzylaminopurine (6-BA) mg/L | Naphthaleneacetic Acid (NAA) mg/L | Indole-3-Acetic Acid (IAA) mg/L | 2,4-Dichlorophenoxyacetic Acid (2,4-D) mg/L | Media |
|---|---|---|---|---|---|---|---|---|
| Shoots induction | 4.14 | 30 | 7 | 1.0 | M1 | |||
| 4.14 | 30 | 7 | 2.0 | M2 | ||||
| 4.14 | 30 | 7 | 3.0 | M3 | ||||
| 4.14 | 30 | 7 | 4.0 | M4 | ||||
| 4.14 | 30 | 7 | 1.0 | 0.1 | M5 | |||
| 4.14 | 30 | 7 | 2.0 | 0.1 | M6 | |||
| 4.14 | 30 | 7 | 1.0 | 0.2 | M7 | |||
| 4.14 | 30 | 7 | 2.0 | 0.2 | M8 | |||
| 4.14 | 30 | 7 | 1.0 | 0.05 | M9 | |||
| 4.14 | 30 | 7 | 2.0 | 0.05 | M10 | |||
| Root induction | 4.14 | 30 | 7 | R1/MS | ||||
| 2.07 | 30 | 7 | R2 | |||||
| 4.14 | 30 | 7 | 0.1 | R3 | ||||
| 4.14 | 30 | 7 | 0.3 | R4 | ||||
| 4.14 | 30 | 7 | 0.5 | R5 |
| Media | Lines | Days for Shoot Induction (d) | Shoots Induction Rate (%) | Average Number of Shoots |
|---|---|---|---|---|
| M2 | L-1 | 13.33 ± 0.58 a | 85.34 ± 0.10 c | 1.19 ± 0.03 c |
| L-2 | 9.33 ± 0.58 c | 78.29 ± 0.27 d | 1.19 ± 0.02 c | |
| L-3 | 8.33 ± 0.58 c | 91.45 ± 0.44 a | 1.37 ± 0.06 b | |
| L-4 | 13.33 ± 0.58 a | 60.29 ± 0.28 e | 1.10 ± 0.03 d | |
| L-5 | 10.67 ± 0.58 b | 87.52 ± 0.27 b | 1.14 ± 0.01 a | |
| M6 | L-1 | 13.67 ± 0.58 ab | 91.16 ± 0.67 a | 1.23 ± 0.02 d |
| L-2 | 9.67 ± 0.58 c | 74.93 ± 0.79 c | 1.56 ± 0.02 b | |
| L-3 | 9.33 ± 0.58 c | 71.58 ± 0.36 d | 1.61 ± 0.03 a | |
| L-4 | 14.33 ± 0.58 a | 72.53 ± 0.18 e | 1.08 ± 0.01 e | |
| L-5 | 12.67 ± 0.58 c | 78.14 ± 0.09 b | 1.36 ± 0.03 c | |
| M8 | L-1 | 15.33 ± 0.58 a | 72.50 ± 0.16 e | 1.20 ± 0.03 c |
| L-2 | 9.67 ± 0.58 c | 85.67 ± 0.21 c | 1.51 ± 0.02 b | |
| L-3 | 8.67 ± 0.58 c | 95.23 ± 0.19 a | 1.87 ± 0.06 a | |
| L-4 | 14.67 ± 0.58 a | 80.33 ± 2.19 d | 1.05 ± 0.03 d | |
| L-5 | 11.67 ± 0.58 b | 90.50 ± 0.79 b | 1.20 ± 0.04 c | |
| M10 | L-1 | 15.67 ± 0.58 a | 75.46 ± 0.88 c | 1.18 ± 0.02 c |
| L-2 | 10.33 ± 0.58 c | 71.26 ± 0.22 d | 1.54 ± 0.03 a | |
| L-3 | 9.33 ± 0.58 c | 80.45 ± 0.31 a | 1.56 ± 0.03 a | |
| L-4 | 15.33 ± 0.58 a | 58.84 ± 0.43 e | 1.07 ± 0.02 d | |
| L-5 | 13.00 ± 1.00 b | 77.37 ± 0.04 b | 1.26 ± 0.01 b |
| Media | Shoots Induction Rate of L-1 (%) | Shoots Induction Rate of L-2 (%) | Shoots Induction Rate of L-3 (%) | Shoots Induction Rate of L-4 (%) | Shoots Induction Rate of L-5 (%) | Average Shoot Induction Rate (%) |
|---|---|---|---|---|---|---|
| M2 | 85.34 ± 0.10 b | 78.29 ± 0.27 b | 91.45 ± 0.44 b | 60.29 ± 0.28 c | 87.52 ± 0.27 b | 80.58 |
| M6 | 91.16 ± 0.67 a | 74.93 ± 0.79 c | 71.58 ± 0.36 d | 72.53 ± 0.18 b | 78.14 ± 0.09 c | 77.67 |
| M8 | 72.50 ± 0.16 d | 85.67 ± 0.21 a | 95.23 ± 0.19 a | 80.33 ± 2.19 a | 90.50 ± 0.79 a | 84.85 |
| M10 | 75.46 ± 0.88 c | 71.26 ± 0.22 d | 80.45 ± 0.31 c | 58.84 ± 0.43 c | 77.37 ± 0.04 c | 72.68 |
| Source | Type III Sum of Squares | df | Mean Square | F | Sig. | Partial Eta Squared |
|---|---|---|---|---|---|---|
| Corrected Model | 5543.517 a | 19 | 291.764 | 703.343 | 0.000 | 0.997 |
| Intercept | 373,905.625 | 1 | 373,905.625 | 901,357.501 | 0.000 | 1.000 |
| Lines | 2148.940 | 4 | 537.235 | 1295.089 | 0.000 | 0.992 |
| Media | 1176.469 | 3 | 392.156 | 945.354 | 0.000 | 0.986 |
| Lines * Media | 2218.107 | 12 | 184.842 | 445.591 | 0.000 | 0.993 |
| Error | 16.593 | 40 | 0.415 | |||
| Total | 379,465.736 | 60 | ||||
| Corrected Total | 5560.110 | 59 |
| Media | Average Rooting Days | Average Root Induction Rate | Average Number of Roots |
|---|---|---|---|
| R1 | 13.33 ± 0.58 a | 88.89 ± 1.93 b | 2.15 ± 0.02 d |
| R2 | 12.33 ± 0.58 a | 95.55 ± 0.84 b | 3.54 ± 0.02 c |
| R3 | 8.67 ± 0.58 b | 97.77 ± 1.93 a | 4.73 ± 0.09 a |
| R4 | 9.67 ± 0.58 b | 73.54 ± 0.22 c | 2.05 ± 0.19 b |
| R5 | 12.67 ± 0.58 a | 66.87 ± 0.26 d | 1.55 ± 0.02 c |
| Lines of Regenerated Plants | Node Position of the First Female Flower | Fruit Development Period/d | Whole Growth Period/d | Average Fruit Weight/kg |
|---|---|---|---|---|
| L-1 | 9–12 | 56 | 102 | 2.53 |
| L-3 | 8–10 | 47 | 99 | 1.77 |
| L-5 | 7–9 | 52 | 95 | 0.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Chen, Z.; Yan, J.; Jiang, H.; Dai, S.; Yang, H.; Sun, L.; Sun, X. Optimization of an Efficient Direct Shoot Induction System and Histological Evaluation of Shoot Formation in Cucurbita maxima Duch. Horticulturae 2024, 10, 165. https://doi.org/10.3390/horticulturae10020165
Song H, Chen Z, Yan J, Jiang H, Dai S, Yang H, Sun L, Sun X. Optimization of an Efficient Direct Shoot Induction System and Histological Evaluation of Shoot Formation in Cucurbita maxima Duch. Horticulturae. 2024; 10(2):165. https://doi.org/10.3390/horticulturae10020165
Chicago/Turabian StyleSong, Huijuan, Zhongquan Chen, Jiali Yan, Hong Jiang, Sihui Dai, Hongbo Yang, Longjun Sun, and Xiaowu Sun. 2024. "Optimization of an Efficient Direct Shoot Induction System and Histological Evaluation of Shoot Formation in Cucurbita maxima Duch" Horticulturae 10, no. 2: 165. https://doi.org/10.3390/horticulturae10020165
APA StyleSong, H., Chen, Z., Yan, J., Jiang, H., Dai, S., Yang, H., Sun, L., & Sun, X. (2024). Optimization of an Efficient Direct Shoot Induction System and Histological Evaluation of Shoot Formation in Cucurbita maxima Duch. Horticulturae, 10(2), 165. https://doi.org/10.3390/horticulturae10020165