

Identification and Pathogenicity of Biscogniauxia and Sordaria Species Isolated from Olive Trees



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fieldwork and Isolation of Fungi
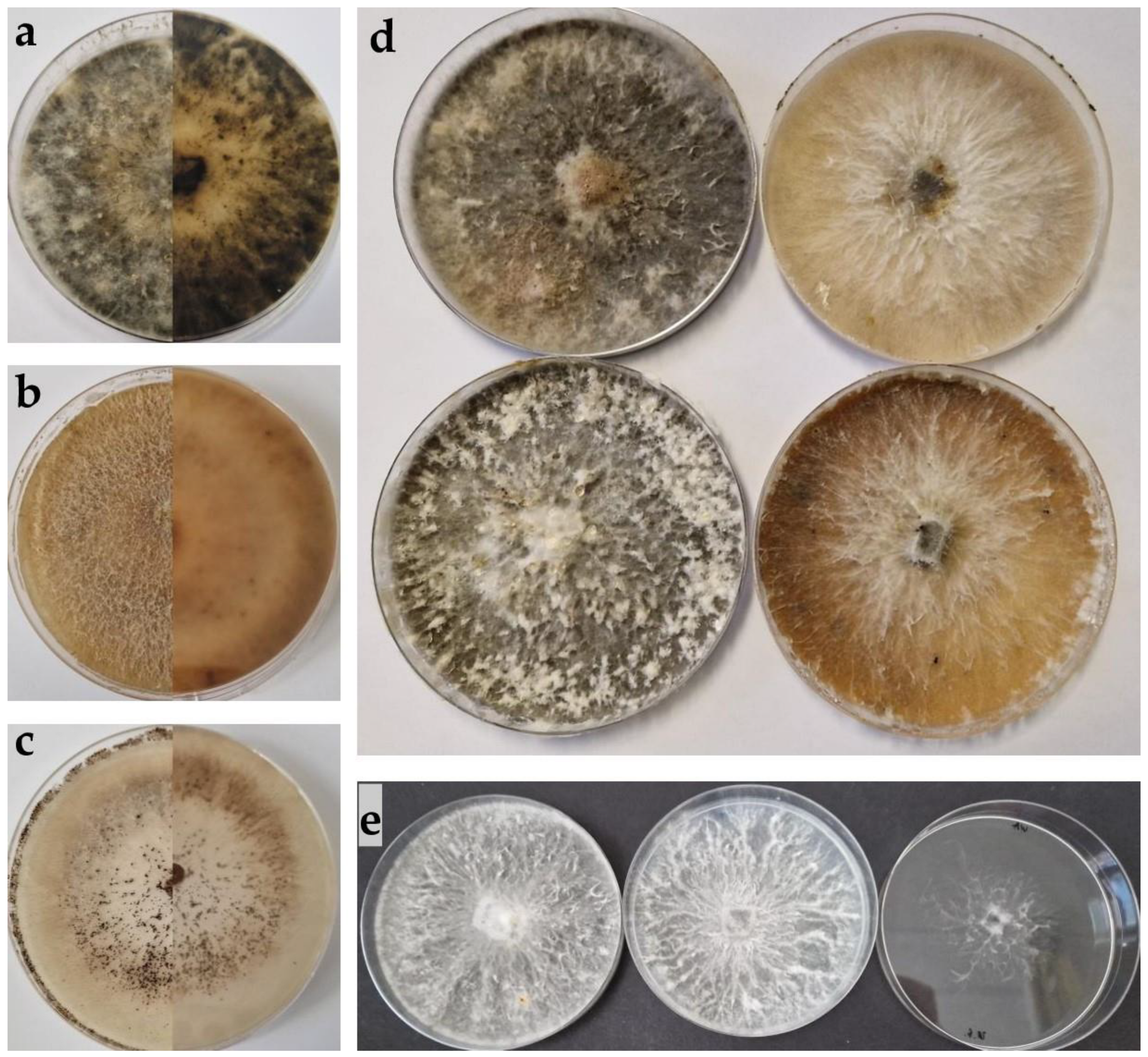
2.2. Morphological Characterisation
2.3. DNA Extraction and Amplification
2.4. DNA Sequence Assembly and Phylogenetic Analysis
2.5. Pathogenicity Tests on Olive Seedlings
2.6. Analysis of Meteorological Data
3. Results
3.1. Field Symptoms
3.2. Morphological Characterisation
3.2.1. Biscogniauxia mediterranea
3.2.2. Biscogniauxia nummularia
3.2.3. Sordaria fimicola
3.3. Molecular Phylogenetic Identification
3.4. Pathogenicity Test
3.5. Meteorological Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Torres, M.; Pierantozzi, P.; Searles, P.; Rousseaux, M.C.; García-Inza, G.; Miserere, A.; Bodoira, R.; Contreras, C.; Maestri, D. Olive cultivation in the southern hemisphere: Flowering, water requirements and oil quality responses to new crop environments. Front. Plant Sci. 2017, 8, 1830. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations. Crop and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 25 January 2024).
- Cowling, R.M.; Rundel, P.W.; Lamont, B.B.; Arroyo, M.K.; Arianoutsou, M. Plant diversity in mediterranean climate regions. Trends Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef]
- Seletković, Z.; Tikvić, I.; Vučetić, M.; Ugarović, D. Climatic features and the vegetation of Mediterranean Croatia. In Šume Hrvatskog Sredozemlja; Akademija Šumarskih Znanosti: Zagreb, Croatia, 2011. [Google Scholar]
- Úrbez-Torres, J.R.; Lawrence, D.P.; Hand, F.P.; Trouillas, F.P. Olive twig and branch dieback in California caused by Cytospora oleicola and the newly described species Cytospora olivarum sp. nov. Plant Dis. 2020, 104, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Petrović, E.; Vrandečić, K.; Ivić, D.; Ćosić, J.; Godena, S. First Report of Olive Branch Dieback in Croatia Caused by Cytospora pruinosa Defago. Microorganisms 2023, 11, 1679. [Google Scholar] [CrossRef]
- Rumbos, I.C. Cytospora oleina causing canker and dieback of olive in Greece. Plant Pathol. 1988, 37, 441–444. [Google Scholar] [CrossRef]
- Moral, J.; Agusti-Brisach, C.; Pérez-Rodríguez, M.; Xaviér, C.; Carmen-Raya, M.; Rhouma, A.; Trapero, A. Identification of fungal species associated with branch dieback of olive and resistance of table cultivars to Neofusicoccum mediterraneum and Botryosphaeria Dothidea. Plant Dis. 2017, 101, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.C.; Roux, J.; Wingfield, M.J. Cytospora species (Ascomycota, Diaporthales, Valsaceae): Introduced and native pathogens of trees in South Africa. Australas. Plant Pathol. 2006, 35, 521–548. [Google Scholar] [CrossRef]
- van Dyk, M.; Spies, C.F.J.; Mostert, L.; van der Rijst, M.; du Plessis, I.L.; Moyo, P.; van Jaarsveld, W.J.; Haleen, F. Pathogenicity testing of fungal isolates associated with olive trunk diseases in South Africa. Plant Dis. 2021, 105, 4060–4073. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R.; Peduto, F.; Vossen, P.M.; Krueger, W.H.; Gubler, W.D. Olive twig and branch dieback: Etiology, incidence, and distribution in California. Plant Dis. 2013, 97, 231–244. [Google Scholar] [CrossRef]
- Kaliterna, J.; Miličević, T.; Ivić, D.; Benčić, D.; Mesić, A. First report of Diplodia seriata as causal agent of olive dieback in Croatia. Plant Dis. 2012, 96, 290. [Google Scholar] [CrossRef]
- Ivić, D.; Petrović, E.; Godena, S. Fungi associated with canker diseases on olive in Istria (Croatia). J. Cent. Eur. Agric. 2023, 24, 470–475. [Google Scholar] [CrossRef]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Phylogeny and morphology of Diplodia species on olives in southern Italy and description of Diplodia olivarum sp nov. Fungal Divers. 2008, 21, 63–71. [Google Scholar]
- Carlucci, A.; Raimondo, M.l.; Cibelli, F.; Phillips, A.J.I.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum associated with a decline of olives in southern Italy. Phytopathol. Mediterr. 2013, 52, 517–527. [Google Scholar]
- Linaldeddu, B.T.; Rossetto, G.; Maddau, L.; Vatrano, T.; Bregant, C. Diversity and Pathogenicity of Botryosphaeriaceae and Phytophthora Species Associated with Emerging Olive Diseases in Italy. Agriculture 2023, 13, 1575. [Google Scholar] [CrossRef]
- Palou, L.; Taberner, V.; Montesinos-Herrero, C. First Report of Diplodia seriata Causing Loquat Fruit Rot in Spain. Plant Dis. 2013, 97, 421–422. [Google Scholar] [CrossRef] [PubMed]
- Spies, C.F.J.; Mostert, L.; Carlucci, A.; Moyo, P.; van Jaarsveld, W.J.; du Plessis, I.L.; van Dyk, M.; Halleen, F. Dieback and decline pathogens of olive trees in South Africa. Persoonia 2020, 45, 196–220. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Rodríguez, L.; Mondino-Hintz, P.; Alaniz-Ferro, S. Diversity of Botryosphaeriaceae species causing stem canker and fruit rot in olive trees in Uruguay. J. Phytopathol. 2022, 170, 264–277. [Google Scholar] [CrossRef]
- Petrović, E.; Vrandečić, K.; Ćosić, J.; Kanižai Šarić, G.; Godena, S. First Report of Phaeoacremonium iranianum Causing Olive Twig and Branch Dieback. Plants 2022, 11, 3578. [Google Scholar] [CrossRef]
- Raimondo, M.L.; Lops, F.; Carlucci, A. First Report of Phaeoacremonium oleae and P. viticola Associated with Olive Trunk Diseases in Italy. Plant Dis. 2022, 106, 331. [Google Scholar] [CrossRef]
- Markakis, E.A.; Soultatos, S.K.; Koubouris, G.C.; Psarras, G.; Kanetis, L.; Papadaki, A.A.; Goumas, D.E. First Report of Pseudophaeomoniella oleae Causing Wood Streaking and Decay on Olive Trees in Greece. Plant Dis. 2022, 106, 2263. [Google Scholar] [CrossRef] [PubMed]
- Agustí-Brisach, C.; Jiménez-Urbano, J.P.; Raya, M.D.; López-Moral, A.; Trapero, A. Vascular Fungi Associated with Branch Dieback of Olive in Super-High-Density Systems in Southern Spain. Plant Dis. 2021, 105, 797–818. [Google Scholar] [CrossRef]
- Ahimera, N.; Gisler, S.; Morgan, D.P.; Michailides, T.J. Effects of single-drop impactions and natural and simulated rains on the dispersal of Botryosphaeria dothidea conidia. Phytopathology 2004, 94, 1189–1197. [Google Scholar] [CrossRef]
- van Niekerk, J.M.; Calitz, F.J.; Halleen, F.; Fourie, P.H. Temporal spore dispersal patterns of grapevine trunk pathogens in South Africa. Eur. J. Plant Pathol. 2010, 127, 375–390. [Google Scholar] [CrossRef]
- Moyo, P.; Allsopp, E.; Roets, F.; Mostert, L.; Halleen, F. Arthropods Vector Grapevine Trunk Disease Pathogens. Phytopathology 2014, 104, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Bertrant, P.F.; English, H. Release and dispersal of conidia of Valsa leucostoma. Phytopathology 1976, 66, 987–991. [Google Scholar] [CrossRef]
- Gharbi, Y.; Ennouri, K.; Bouazizi, E.; Cheffi, M.; Ali Triki, M. First report of charcoal disease caused by Biscogniauxia mediterranea on Olea europaea in Tunisia. J. Plant Pathol. 2020, 102, 961. [Google Scholar] [CrossRef]
- Stadler, M. Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny, and functional biodiversity. Curr. Res. Environ. Appl. Mycol. 2011, 1, 75–133. [Google Scholar] [CrossRef]
- Kuntze, O. Revisio Generum Plantarum; Arthur Felix: Leipzig, Germany, 1891; pp. 375–1001. [Google Scholar]
- Granata, G.; Sidoti, A. Biscogniauxia nummularia: Pathogenic agent of a beech decline. For. Pathol. 2004, 34, 363–367. [Google Scholar] [CrossRef]
- Petrini, O. Fungal endophytes of trees leaves. In Microbial Ecology of Leaves; Abdews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Petrini, L.; Petrini, O. Xylariaceous fungi as endophytes. Sydowia 1985, 38, 216–234. [Google Scholar]
- Nugent, L.K.; Sihanonth, P.; Thienhirun, S.; Whalley, A.J.S. Biscogniauxia: A genus of latent invaders. Mycologist 2005, 19, 40–43. [Google Scholar] [CrossRef]
- Collado, J.; Platas, G.; Pelaez, F. Identification of an endophytic Nodulisporium sp. from Quercus ilex in central Spain as the anamorph of Biscogniauxia mediterranea by rDNA sequences analysis and effect of different ecological factors on distribution of the fungus. Mycologia 2001, 93, 875–886. [Google Scholar] [CrossRef]
- Luchi, N.; Capretti, P.; Feducci, M.; Vannini, A.; Ceccarelli, B.; Vettraino, A.M. Latent infection of Biscogniauxia nummularia in Fagus sylvatica: A possible bioindicator of beech health conditions. iForest 2015, 9, 49–54. [Google Scholar] [CrossRef]
- Hendry, S.J.; Boddy, L.; Lonsdale, D. Abiotic variables effect differential expression of latent infections in beech (Fagus sylvatica). New Phytol. 2002, 155, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Yangui, I.; Boutiti, M.Z.; Messaoud, C.; Lahbib Ben Jamma, M.; Vannini, A.; Vettraino, A.M. First report of Biscogniauxia mediterranea causing canker on Erica multiflora L. in Tunisia. J. Plant Pathol. 2019, 1001, 1273. [Google Scholar] [CrossRef]
- Rostamian, M.; Kavosi, M.R.; Bazgir, E.; Babanezhad, M. First report of Biscogniauxia mediterranea causing canker on wild almond (Amygdalus scoparia). Australas. Plant Dis. Notes 2016, 11, 30. [Google Scholar] [CrossRef]
- Mirabolpathi, M. Outbreak of charcoral disease on Quercus spp. and Zelkova carpinifolia trees in forest of Zagros and Albroz Mountains in Iran. Iran. J. Plant Pathol. 2013, 49, 257–263. [Google Scholar]
- Zabalgogeazcoa, I.; Pedro, J.; Canalis, R.M. Biscogniauxia nummularia infecting beech (Fagus sylvatica) trees and sympatric plants of the sedge Carex brevicollis. For. Pathol. 2015, 45, 346–348. [Google Scholar] [CrossRef]
- Patejuk, K.; Baturo-Cieśniewska, A.; Pusz, W.; Kaczmarek-Pieńczewska, A. Biscogniauxia Charcoal Canker—A New Potential Threat for Mid-European Forests as an Effect of Climate Change. Forests 2022, 13, 89. [Google Scholar] [CrossRef]
- La Porta, N.; Capretti, P.; Thomsen, I.M.; Kasanen, R.; Hietala, A.M.; Von Weissenberg, K. Forest pathogens with higher damage potential due to climate change in Europe. Can. J. Plant Pathol. 2008, 30, 177–195. [Google Scholar] [CrossRef]
- Ivanová, H.; Onderková, A.; Pristaš, P. Sordaria fimicola-like ascomycete isolated from Pinus coulteri needles in Slovakia. Biologia 2018, 73, 553–559. [Google Scholar] [CrossRef]
- Olive, L.S. Sordaria. In Bacteria, Bacteriophages, and Fungi; King, R.C., Ed.; Springer: Boston, MA, USA, 1974; pp. 553–562. [Google Scholar]
- Ivanová, H. Sordaria fimicola (Ascomycota, Sordariales) on Acer palmatum. Folia Oecologica 2015, 42, 67–71. [Google Scholar]
- Newcombe, G.; Campbell, J.; Griffith, D.; Baynes, M.; Launchbaugh, K.; Pendleton, R. Revisiting the life cycle of dung fungi, including Sordaria fimicola. PLoS ONE 2016, 11, e0147425. [Google Scholar] [CrossRef] [PubMed]
- Hanlin, R.T. Ilustrated Genera of Ascomycetes; APS Press: St. Paul, MN, USA, 1990. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. 38—Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR—Protocols and Applications—A Laboratory Manual; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1995, 91, 553–556. [Google Scholar] [CrossRef]
- Mazzaglia, A.; Anselmi, N.; Gasbarri, A.; Vannini, A. Development of a Polymerase Chain Reaction (PCR) assay for the specific detection of Biscogniauxia mediterranea living as an endophyte in oak tissues. Mycol. Res. 2001, 105, 952–956. [Google Scholar] [CrossRef]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined Multiple Gene Genealogies and Phenotypic Characters Differentiate Several Species Previously Identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Rogers, J.D.; Ju, Y.-M.; Candoussau, F. Biscogniauxia anceps comb. nov. and Vivantia guadalupensis gen. et sp. nov. Mycol. Res. 1996, 100, 669–674. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef]
- Ju, Y.-M.; Rogers, J.D.; San Martín, F.; Granmo, A. The genus Biscogniauxia. Mycotaxon 1998, 66, 1–98. [Google Scholar]
- Mirabolfathy, M.; Ju, Y.-M.; Hsieh, H.-M.; Rogers, J.D. Obolarina persica sp. nov., associated with dying Quercus in Iran. Mycoscience 2013, 54, 315–320. [Google Scholar] [CrossRef]
- Pinto-Sherer, T.J. The Phyloecology of Hypaxylan Sensu Lata; B.A. Florida International University: Miami, FL, USA, 1996. [Google Scholar]
- Sánchez-Ballesteros, J.; González, V.; Salazar, O.; Acero, J.; Portal, M.A.; Julián, M.; Rubio, V.; Bills, G.F.; Polishook, J.D.; Platas, G.; et al. Phylogenetic study of Hypoxylon and related genera based on ribosomal ITS sequences. Mycologia 2000, 92, 964–977. [Google Scholar] [CrossRef]
- Ju, Y.-M.; Rogers, J.D. New and interesting Biscogniauxia taxa, with a key to the world species. Mycol. Res. 2001, 105, 1123–1133. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Stadler, M.; Jones, E.B.G.; Promputtha, I.; Suwannarach, N.; Camporesi, E.; Bulgakov, T.S.; Liu, J.-K. Taxonomy, phylogeny, molecular dating and ancestral state reconstruction of Xylariomycetidae (Sordariomycetes). Fungal Divers. 2022, 112, 1–88. [Google Scholar]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Raimondo, M.L.; Lops, F.; Carlucci, A. Charcoal canker of pear, plum, and quince trees caused by Biscogniauxia rosaceum sp. nov. in southern Italy. Plant Dis. 2016, 100, 1813–1822. [Google Scholar] [CrossRef] [PubMed]
- Henriques, J.; Nóbrega, F.; Sousa, E.; Lima, A. Analysis of the genetic diversity and phylogenetic relationships of Biscogniauxia mediterranea isolates associated with cork oak. Phytoparasitica 2016, 44, 19–34. [Google Scholar] [CrossRef]
- Mirabolfathy, M.; Groenewald, J.Z.; Crous, P.W. The Occurrence of Charcoal Disease Caused by Biscogniauxia mediterranea on Chestnut-Leaved Oak (Quercus castaneifolia) in the Golestan Forests of Iran. Plant Dis. 2011, 95, 876. [Google Scholar] [CrossRef] [PubMed]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenetics Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [PubMed]
- Pažoutová, S.; Šrůtka, P.; Holuša, J.; Chudíčková, M.; Kolařík, M. The phylogenetic position of Obolarina dryophila (Xylariales). Mycol. Prog. 2010, 9, 501–507. [Google Scholar] [CrossRef]
- Bien, S.; Damm, U. Prunus trees in Germany—A hideout of unknown fungi? Mycol. Prog. 2020, 19, 667–690. [Google Scholar] [CrossRef]
- Blumenstein, K.; Bußkamp, J.; Langer, G.J.; Langer, E.J.; Terhonen, E. The Diplodia tip blight pathogen Sphaeropsis sapinea is the most common fungus in scots pines’ mycobiome, irrespective of health status-a case study from Germany. J. Fungi 2021, 7, 607. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Cai, L.; Jeewon, R.; Hyde, K.D. Phylogenetic investigations of Sordariaceae based on multiple gene sequences and morphology. Mycol. Res. 2006, 110, 137–150. [Google Scholar] [CrossRef]
- Pecchia, S.; Caggiano, B.; Da Lio, D.; Cafà, G.; Le Floch, G.; Baroncelli, R. Molecular Detection of the Seed-Borne Pathogen Colletotrichum lupini Targeting the Hyper-Variable IGS Region of the Ribosomal Cluster. Plants 2019, 8, 222. [Google Scholar] [CrossRef]
- Nygren, K.; Strandberg, R.; Wallberg, A.; Nabholz, B.; Gustafsson, T.; García, D.; Cano, J.; Guarro, J.; Johannesson, H. A comprehensive phylogeny of Neurospora reveals a link between reproductive mode and molecular evolution in fungi. Mol. Phylogenet. Evol. 2011, 59, 649–663. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- The Croatian Meteorological and Hydrological Service. Available online: https://meteo.hr/infrastruktura.php?section=mreze_postaja¶m=pmm&el=klimatoloske (accessed on 4 December 2023).
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Osono, T.; Tateno, O.; Masuya, H. Diversity and ubiquity of xylariaceous endophytes in live and dead leaves of temperate forest trees. Mycoscience 2013, 54, 54–61. [Google Scholar] [CrossRef]
- Macara, A.M. Estimativa em 1975 dos Prejuízos Causados Pelas Principais Doenças do Sobreiro Num Montado da Região Ribatejana; Boletim do Instituto dos Podutos Florestais; Cortiça Nº444: Lisabon, Portugal, 1975. [Google Scholar]
- Materatski, P.; Varanda, C.; Carvalho, T.; Dias, A.B.; Campos, M.D.; Rei, F.; Félix, M.R. Spatial and temporal variation of fungal enophytic richness and diversity associated to the phyllosphere of olive cultivars. Fungal Biol. 2019, 123, 66–76. [Google Scholar] [CrossRef]
- Godena, S.; Ivić, D.; Goreta Ban, S. Uzročnici Djelomičnog ili Potpunog Sušenja Stabala Maslina; Priručnik o Rezultatima VIP Projekta; Institut za Poljoprivredu i Turizam: Poreč, Croatia, 2019; pp. 1–49. [Google Scholar]
- Dminić, D.; Orlović, J.K.; Lukić, I.; Ježić, M.; Ćurković Perica, M.; Pernek, M. First Report of Charcoal Disease of Oak (Biscogniauxia mediterranea) on Quercus spp. in Croatia. Plant Dis. 2019, 103, 2687. [Google Scholar] [CrossRef]
- Fields, W.G. An introduction to the genus Sordaria. Neurospora Newsl. 1970, 16, 14–17. [Google Scholar] [CrossRef]
- Vaninni, A.; Paganini, R.; Anselmi, N. Factors affecting discharge and germination of ascospores of Hypoxylon mediterraneum (De Not.) Mill. Eur. J. For. Pathol. 1996, 26, 12–24. [Google Scholar] [CrossRef]
- Saharan, G.S.; Mehta, N. Sclerotinia Diseases of Crop Plants: Biology, Ecology and Disease Management; Springer: New York, NY, USA, 2008. [Google Scholar]
- Ouedraogo, A.; Fargues, J.; Goettel, M.S.; Lomer, C.J. Effect of temperature on vegetative growth among isolates of Metarhizium anisopliae and M. flavoviride. Mycopathologia 1997, 137, 37–43. [Google Scholar] [CrossRef]
- Gauthier, G.M. Dimorphism in fungal pathogens of mammals, plants, and insects. PLoS Pathog. 2015, 11, e1004608. [Google Scholar] [CrossRef]
- Henriques, J.; Nóbrega, F.; Sousa, E.; Lima, A. Diversity of Biscogniauxia mediterranea within single stromata on cork oak. J. Mycol. 2014, 2014, 324349. [Google Scholar] [CrossRef]
- Vannini, A.; Mazzaglia, A.; Anselmi, N. Use of random amplified polymorphic DNA (RAPD) for detection of genetic variation and proof of the heterothallic mating system in Hypoxylom mediterraneum. Eur. J. Forrest Pathol. 1999, 29, 209–218. [Google Scholar] [CrossRef]
- Jiménez, J.J.; Sánchez, M.E.; Trapero, A. El chancro carbonoso de Quercus III: Dispersión de ascosporas del agente causal. Boletín Sanid. Veg. Plagas 2005, 31, 577–585. [Google Scholar]
- Henriques, J.; Barrento, M.J.; Bonifácio, L.; Gomes, A.A.; Lima, A.; Sousa, E. Factors affecting the dispersion of Biscogniauxia mediterranea in Portuguese cork oak stands. Silva Lusit. 2014, 22, 83–97. [Google Scholar]
- Henriques, J.; Inácio, M.L.; Lima, A.; Sousa, E. New outbreaks of charcoal canker on young cork oak trees in Portugal. Integr. Prot. Oak For. IOBC/Wprs Bull. 2012, 76, 85–88. [Google Scholar]
- Sousa, E.M.R.d.; Santos, M.N.S.; Varela, M.C.; Henriques, J. Perda do Vigor dos Montados de Sobro e Azinho: Análise da Situação e Perspetivas; Instituto Nacional de Investigação Agrária e Veterinária, I.P.: Oeiras, Portugese, 2007. [Google Scholar]
- García, F.; Bestiona, E.; Warfielda, R.; Yvon-Durochera, G. Changes in temperature alter the relationship between biodiversity and ecosystem functioning. Proc. Natl. Acad. Sci. USA 2018, 115, 201805518. [Google Scholar] [CrossRef]
- Brown, A.J.P.; Budge, S.; Kaloriti, D.; Tillmann, A.; Jacobsen, M.D.; Yin, Z.; Ene, I.V.; Bohovych, I.; Sandai, D.; Stavroula, K.; et al. Stress adaptation in a pathogenic fungus. J. Exp. Biol. 2014, 217, 144–155. [Google Scholar] [CrossRef]
- Kim, G.-U.; Chen, D. Climate change over the Mediterranean and current destruction of marine ecosystem. Sci. Rep. 2019, 9, 18813. [Google Scholar] [CrossRef]
- Burdon, J.J.; Silk, J. Sources and Patterns of Diversity in Plant-Pathogenic Fungi. Phytopathology 1997, 87, 664–669. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Keesing, F.; Eviner, V.T. The Ecology of Infectious Diseases: Progress, Challenges, and Frontiers. In Infectious Disease Ecology: Effects of Ecosystems on Disease and of Disease on Ecosystems; Ostfeld, R.S., Keesing, F., Eviner, V.T., Eds.; Princeton University Press: Princeton, NJ, USA, 2008; pp. 469–482. [Google Scholar]
- Rupp, S.; Sohn, K. Host-Pathogen Interactions: Methods and protocols. In Methods in Molecular Biology; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2009; Volume 470. [Google Scholar]
- Lugtenberg, B. Principles of Plant-Microbe Interactions: Microbe for Sustainable Agriculture; Springer: New York, NY, USA, 2015. [Google Scholar]
- Karami, J.; Kavosi, M.R.; Babanezhad, M.; Kiapasha, K. Integrated management of the charcoal disease by silviculture, chemical and biological methods in forest parks. J. Sustain. For. 2018, 37, 429–444. [Google Scholar] [CrossRef]
- Yangui, I.; Boutiti, M.Z.; Boussaid, M.; Messaoud, C. Essential Oils of Myrtaceae Species Growing Wild in Tunisia: Chemical Variability and Antifungal Activity Against Biscogniauxia mediterranea, the Causative Agent of Charcoal Canker. Chem. Biodivers. 2017, 14, e1700058. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Basionym |
|---|---|
| Biscogniauxia baileyi (Berk. and Broome ex Cooke) Kuntze | Nummularia baileyi Berk. and Broome ex Cooke |
| Biscogniauxia bulliardii (Tul. and C. Tul.) Kuntze | Nummularia bulliardii Tul. and C. Tul. |
| Sphaeria discreta Schwein. | Sphaeria discreta Schwein. |
| Obolarina dryophila (Tul. and C. Tul.) | Nummularia dryophila Tul. and C. Tul. |
| Camillea obularia (Fr.) Læssøe | Hypoxylon obularium Fr. |
| Biscogniauxia repanda (Fr.) Kuntze | Sphaeria repanda Fr. |
| Biscogniauxia mediterranea (De Not.) Kuntze | Sphaeria mediterranea De Not. |
| Species | Isolate | Isolation Source | Country | GenBank Accession Number | References | ||
|---|---|---|---|---|---|---|---|
| ITS | TUB2 | TEF1- α | |||||
| Biscogniauxia anceps | YMJ 123 | Corylus avellana L. | France | EF026132 | AY951671 | / | [55,56] |
| B. arima F. San Martín, Y.M. Ju and J.D. Rogers | YMJ 122 | Wood | Mexico | EF026150 | AY951672 | / | [56,57] |
| B. atropunctata | YMJ 128 | Wood | United States of America | JX507799 | AY951673 | / | [57,58] |
| ATCC 38987 | Unknown | United States of America | AF201705 | / | / | [59] | |
| B. atropunctata var. intermedia | B70M | Quercus sp. | Costa Rica | AJ390412 | / | / | [60] |
| B. bartholomaei | ATCC 38992 | Unknown | United States of America | AF201719 | / | / | [59] |
| B. capnodes | YMJ 142 | Nothofagus solandri (Hook.f.) Oerst. | New Zealand | / | AY951674 | / | [56] |
| YMJ 138 | Corticated wood | Taiwan | EF026131 | AY951675 | / | [57,58] | |
| B. citriformis | YMJ 88113012 | Wood | Taiwan | JX507800 | AY951677 | / | [56] |
| YMJ 129 | Casuarina equisetifolia L. | Hawaii, USA | JX507801 | AY951678 | / | [57,58] | |
| B. cylindrispora Y.M. Ju and J.D. Rogers | YMJ 89092701 | Bark of Cinnamomum Schaeff. | Taiwan | EF026133 | AY951679 | / | [56,61] |
| B. formosana Y.M. Ju and J.D. Rogers | YMJ 89032201 | Bark | Taiwan | JX507802 | AY951680 | / | [56,58,61] |
| B. granmoi | YMJ 135 | Bark of Prunus padus L. | Austria | JX507803 | AY951681 | / | [56,57,58] |
| B. latirima Y.M. Ju and J.D. Rogers | YMJ 90080703 | Bark | Taiwan | EF026135 | AY951683 | / | [56,61] |
| B. magna Samarak. and K.D. Hyde | MFLU 18-0850 | Dead branch from unidentified host | Thailand | MW240616 | MW775577 | MW759498 | [62] |
| B. marginata | MFLUCC 12-0740 | Unknown | France | KJ958407 | / | / | [63] |
| B74A | Quercus sp. | Pennsylvania, USA | AJ390417 | / | / | [60] | |
| B. mediterranea | YMJ 147 | Corticated wood | France | EF026134 | AY951684 | / | [56,57,58] |
| Bx63 | Quercus pubescens Willd. | Italy | KT253501 | KT253535 | / | [64] | |
| Bx70 | Q. pubescens | Italy | KT253502 | KT253536 | / | [64] | |
| Bx85 | Q. pubescens | Italy | KT253503 | KT253537 | / | [64] | |
| Bm8L-19Aa | Abies alba Mill. | Poland | MN538267 | MZ221959 | MZ221965 | [42] | |
| Bm04.001 | Quercus suber L. | Portugal | KM216752 | KM267202 | KM216788 | [65] | |
| Bm07.003 | Q. suber | Portugal | KM216754 | KM267203 | KM216790 | [65] | |
| Bm10.019 | Q. suber | Portugal | KM216761 | KM267210 | KM216797 | [65] | |
| Bm10.001 | Q. suber | Portugal | KM216756 | KM267205 | KM216792 | [65] | |
| Bm10.006 | Q. suber | Portugal | KM216757 | KM267206 | KM216793 | [65] | |
| Bm10.012 | Q. rotundifolia Lam. | Portugal | KM216758 | KM267207 | KM216794 | [65] | |
| Bm11.003 | Q. suber | Portugal | KM216764 | KM267212 | KM216800 | [65] | |
| Bm12.027 | Q. robur L. | Portugal | KM216775 | KM267223 | KM216811 | [65] | |
| Bm12.032 | Q. suber | Portugal | KM216777 | KM267225 | KM216813 | [65] | |
| Pc08.002 | Insects Platypus cylindrus Fabricius which were collected directly from their galleries on Q. suber | Portugal | KM216785 | KM267234 | KM216822 | [65] | |
| Pc96.009 | Insects Platypus cylindrus Fabricius which were collected directly from their galleries on Q. suber | Portugal | KM216786 | KM267233 | KM216821 | [65] | |
| Bm10.016 | Q. suber | Italy | KM216759 | KM267208 | KM216795 | [65] | |
| CBS101016 | Q. robur | Netherlands | KM216787 | KM267235 | KM216823 | [65] | |
| Bm13.013 | Q. suber | Spain | KM216784 | KM267232 | KM216820 | [65] | |
| BM03 (BM01-BM03) | Erica multiflora L. | Tunisia | MH356285 | MK210238 | MK189173 | [38] | |
| CPC:18215 | Quercus castaneifolia C.A.Mey. | Iran | JF295127 | / | / | [66] | |
| CPC:18216 | Q. castaneifolia | Iran | JF295128 | / | / | [66] | |
| CPC:18217 | Q. castaneifolia | Iran | JF295129 | / | / | [66] | |
| ARIZ:AZ0703 | Pseudevernia intensa | Arizona, USA | HM123416 | KU684122 | / | [67] | |
| Oe.Bm 1 | Olea europaea L. | Tunisia | KY275264 | KY275263 | / | [28] | |
| B. nummularia | B72C | Fagus sylvatica L. | England | AJ390415 | / | / | [60] |
| BNUMM3 | Unknown | Italy | AJ246231 | / | / | [52] | |
| H86 | Salix alba L. | Slovakia | GQ428318 | GQ428324 | / | [68] | |
| Bn3W-19Pu | Pinus mugo subsp. uncinata (Raymond ex A.DC.) Domin | Poland | MN595068 | MZ221954 | MZ221960 | [42] | |
| Bn5L-19Pu | P. mugo subsp. uncinata | Poland | MN588203 | MZ221955 | MZ221961 | [42] | |
| Bn6L-19Pu | P. mugo subsp. uncinata | Poland | MN588202 | MZ221956 | MZ221962 | [42] | |
| Bn31M-20Aa | A. alba | Poland | MT936553 | MZ221957 | MZ221963 | [42] | |
| Bn56C-20Aa | A. alba | Poland | MT937244 | MZ221958 | MZ221964 | [42] | |
| GLMC 829 | Prunus avium (L.) L. | Germany | MT153623 | / | / | [69] | |
| NWFVA4756 | Pinus sylvestris (L.) L. | Germany | MT790313 | / | / | [70] | |
| B. petrensis Z.F. Zhang, F. Liu and L. Cai | LC5698 | Rock | China | KU746670 | KU746763 | KX855216 | [70] |
| LC5751 | Rock | China | KU746671 | KU746761 | KX855215 | [71] | |
| B. philippinensis var. microspora | YMJ 89041101 | Bark | Taiwan | EF026136 | AY951685 | / | [56,57] |
| B. repanda | B75A | Unknown | Unknown | AJ390418 | / | / | [60] |
| B. rosacearum A. Carlucci and M. L. Raimondo | Bx3 | Cydonia oblonga Mill. | Italy | KT253487 | KT253521 | / | [64] |
| Bx14 | C. oblonga | Italy | KT253488 | KT253522 | / | [64] | |
| Bx19 | C. oblonga | Italy | KT253489 | KT253523 | / | [64] | |
| Bx25 | Q. pubescens | Italy | KT253499 | KT253533 | / | [64] | |
| CSN1052 | O. europaea | South Africa | MT813910 | / | / | [18] | |
| CSN1055 | Wild olive | New Zealand | MT813912 | / | / | [18] | |
| CSN1056 | Wild olive | South Africa | MT813913 | / | / | [18] | |
| PMM2071 | O. europaea | South Africa | MT813997 | / | / | [18] | |
| B. simplicior Pouzar | YMJ 136 | Rhamnus cathartica L. | France | EF026130 | AY951686 | / | [56] |
| B. uniapiculata | YMJ 90080608 | Bark | Taiwan | JX507805 | AY951687 | / | [56] |
| Phaeoacremonium iranianum L. Mostert, Grafenhan, W. Gams and Crous | R18B4 | O. europaea | Croatia | OP627795 | OP684932 | OP684933 | [20] |
| Sordaria alcina N. Lundqvist | CBS 109460 | Unknown | Unknown | AY681198 | AY681232 | / | [72] |
| S. equicola Crous | CBS 1146992 | Zebra dung | Namibia | NR173047 | MZ078267 | MZ078226 | Unpublished |
| S. fimicola | DAFE_SP16-17 | Lupinus sp. | Italy | MK560171 | MK567929 | / | [73] |
| FGSC 2918 | Unknown | New York, USA | / | FR774339 | FR774388 | [74] | |
| CBS 398.63 | Unknown | Argentina | MH858315 | / | / | [75] | |
| CBS 485.64 | Unknown | Netherlands | MH858489 | / | / | [75] | |
| CBS 508.50 | Unknown | Unknown | AY681188 | AY681228 | / | [72] | |
| S. lappae Potebnia | CBS 154.97 | Unknown | Unknown | AY681171 | AY681205 | / | [72] |
| S. macrospora Auerswald | SORDMGRF46 | Grevillea robusta A.Cunn. ex R.Br. | Kenya | / | / | FJ904898 | Unpublished |
| S. tomentoalba Cailleux | CBS 260.78 | Unknown | Unknown | AY681195 | AY681229 | / | [72] |
| GenBank Accession Number | ||||||
|---|---|---|---|---|---|---|
| Isolate | Olive Variety | Collection Date | Location | ITS | TUB2 | TEF1- α |
| SL2 PRI | Porečka rosulja | 25 August 2021 | Kaštelir-Labinci | OQ734646 | OQ744688 | OQ754165 |
| N17 BJA1 | Istarska bjelica | 14 October 2021 | Novigrad | OQ733307 | OQ744682 | OQ744689 |
| R18 B3I | Buža | 14 October 2021 | Rovinj | OQ733361 | OQ744684 | OQ744691 |
| R18 LECII | Leccino | 14 October 2021 | Rovinj | OQ746452 | OQ744687 | OQ744694 |
| R18 LEC1 | Leccino | 14 October 2021 | Rovinj | OQ733486 | OQ744686 | OQ744693 |
| R19 B1 | Buža | 14 October 2021 | Rovinj | OQ733324 | OQ744683 | OQ744690 |
| ISN9 LDC3I | Leccio del Corno | 13 September 2021 | Poreč | OQ729816 | OQ942633 | OQ744681 |
| IMK9 36II | Buža puntoža | 13 September 2021 | Poreč | OQ646787 | OQ725012 | OQ725013 |
| R18 BII | Buža | 14 October 2021 | Rovinj | OQ933231 | OQ744685 | OQ744682 |
| V16 B3 | Buža | 14 October 2021 | Vodnjan | OQ743781 | OQ754166 | OQ754167 |
| SL1 NP2 | Unknown | 25 August 2021 | Sveti Lovreč | OQ828656 | OQ835632 | OQ835629 |
| ISN9PEN | Pendolino | 13 September 2021 | Poreč | OQ828658 | OQ835630 | OQ835631 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrović, E.; Godena, S.; Ćosić, J.; Vrandečić, K. Identification and Pathogenicity of Biscogniauxia and Sordaria Species Isolated from Olive Trees. Horticulturae 2024, 10, 243. https://doi.org/10.3390/horticulturae10030243
Petrović E, Godena S, Ćosić J, Vrandečić K. Identification and Pathogenicity of Biscogniauxia and Sordaria Species Isolated from Olive Trees. Horticulturae. 2024; 10(3):243. https://doi.org/10.3390/horticulturae10030243
Chicago/Turabian StylePetrović, Elena, Sara Godena, Jasenka Ćosić, and Karolina Vrandečić. 2024. "Identification and Pathogenicity of Biscogniauxia and Sordaria Species Isolated from Olive Trees" Horticulturae 10, no. 3: 243. https://doi.org/10.3390/horticulturae10030243
APA StylePetrović, E., Godena, S., Ćosić, J., & Vrandečić, K. (2024). Identification and Pathogenicity of Biscogniauxia and Sordaria Species Isolated from Olive Trees. Horticulturae, 10(3), 243. https://doi.org/10.3390/horticulturae10030243