
A One-Step Polyphenol Removal Approach for Detection of Multiple Phytohormones from Grape Berry




Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Plant Materials
2.2.1. Method Application Materials
2.2.2. Method Optimization Materials
2.3. Physicochemical Index Analysis
2.4. Sample Preparation
2.5. UHPLC-MS/MS Conditions
2.6. Evaluation of Matrix Effects
2.7. Evaluation of the Overall Recovery
2.8. Standard Curve
3. Results and Discussion
3.1. Optimization of the MS/MS Conditions
3.2. Optimization of the Chromatographic Conditions
3.3. Optimization of Sample Pretreatment
3.3.1. Design of Sample Pretreatment
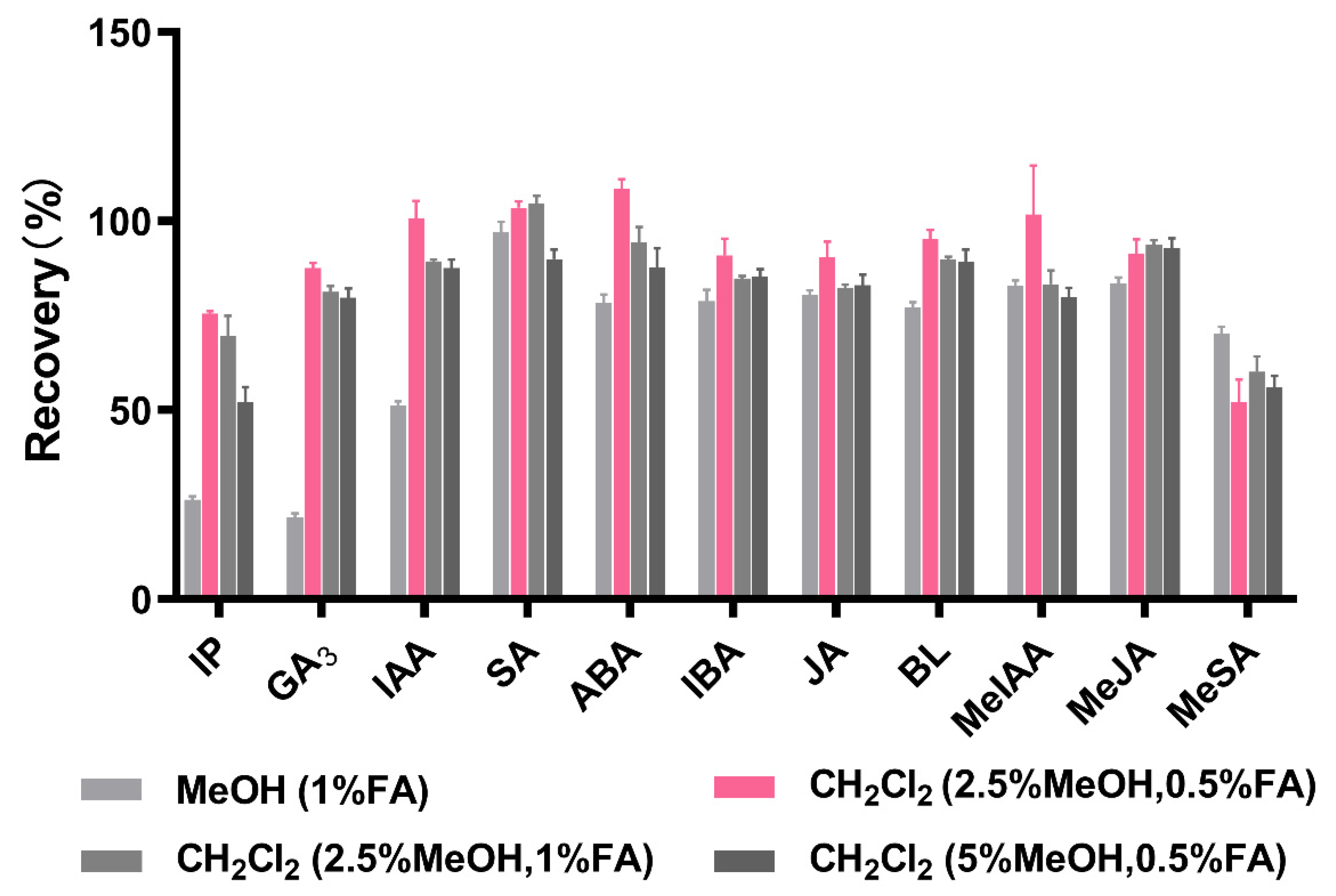
3.3.2. Purification Strategy
3.3.3. Enrichment Process
3.4. Method Verification
3.4.1. Matrix Effect
3.4.2. Recovery and Precision
3.4.3. Linearity
3.5. Method Application
3.5.1. Profiling Phytohormones during Grape Development from Different Vineyards
3.5.2. Profiling Phytohormones in Grape after Exogenous Hormone Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bari, R.; Jones, J.D.G. Role of Plant Hormones in Plant Defence Responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of Phytohormones and Their Signaling Pathways in Leaf Development and Stress Responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, F.; Kühn, N.; Muñoz, M.; Marchandon, G.; Gouthu, S.; Deluc, L.; Delrot, S.; Lauvergeat, V.; Arce-Johnson, P. The Role of Auxin during Early Berry Development in Grapevine as Revealed by Transcript Profiling from Pollination to Fruit Set. Hortic. Res. 2021, 8, 140. [Google Scholar] [CrossRef]
- Jang, G.; Chang, S.H.; Um, T.Y.; Lee, S.; Kim, J.-K.; Choi, Y.D. Antagonistic Interaction between Jasmonic Acid and Cytokinin in Xylem Development. Sci. Rep. 2017, 7, 10212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Sun, J.; Zhai, Q.; Zhou, W.; Qi, L.; Xu, L.; Wang, B.; Chen, R.; Jiang, H.; Qi, J.; et al. The Basic Helix-Loop-Helix Transcription Factor MYC2 Directly Represses PLETHORA Expression during Jasmonate-Mediated Modulation of the Root Stem Cell Niche in Arabidopsis. Plant Cell 2011, 23, 3335–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zander, M.; Chen, S.; Imkampe, J.; Thurow, C.; Gatz, C. Repression of the Arabidopsis thaliana Jasmonic Acid/Ethylene-Induced Defense Pathway by TGA-Interacting Glutaredoxins Depends on Their C-Terminal ALWL Motif. Mol. Plant 2012, 5, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Pérez, A.C.; Goossens, A. Jasmonate Signalling: A Copycat of Auxin Signalling? Plant Cell Environ. 2013, 36, 2071–2084. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Yu, J.-Q.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Benefits of Brassinosteroid Crosstalk. Trends Plant Sci. 2012, 17, 594–605. [Google Scholar] [CrossRef]
- Zhu, Z.; Lee, B. Friends or Foes: New Insights in Jasmonate and Ethylene Co-Actions. Plant Cell Physiol. 2015, 56, 414–420. [Google Scholar] [CrossRef]
- Liu, L.; Li, H.; Zeng, H.; Cai, Q.; Zhou, X.; Yin, C. Exogenous Jasmonic Acid and Cytokinin Antagonistically Regulate Rice Flag Leaf Senescence by Mediating Chlorophyll Degradation, Membrane Deterioration, and Senescence-Associated Genes Expression. J. Plant Growth Regul. 2016, 35, 366–376. [Google Scholar] [CrossRef]
- Nitschke, S.; Cortleven, A.; Iven, T.; Feussner, I.; Havaux, M.; Riefler, M.; Schmülling, T. Circadian Stress Regimes Affect the Circadian Clock and Cause Jasmonic Acid-Dependent Cell Death in Cytokinin-Deficient Arabidopsis Plants. Plant Cell 2016, 28, 1616–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Moore, S.; Chen, C.; Lindsey, K. Crosstalk Complexities between Auxin, Cytokinin, and Ethylene in Arabidopsis Root Development: From Experiments to Systems Modeling, and Back Again. Mol. Plant 2017, 10, 1480–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saniewski, M.; Ueda, J.; Miyamoto, K. Relationships between Jasmonates and Auxin in Regulation of Some Physiological Processes in Higher Plants. Acta Physiol. Plant 2002, 24, 211–220. [Google Scholar] [CrossRef]
- Hofmann, N.R. Taking Hormone Crosstalk to a New Level: Brassinosteroids Regulate Gibberellin Biosynthesis. Plant Cell 2015, 27, 2081. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.W. Auxin as Compère in Plant Hormone Crosstalk. Planta 2009, 231, 1–12. [Google Scholar] [CrossRef]
- Xi, Z.; Zhang, Z.; Huo, S.; Luan, L.; Gao, X.; Ma, L.; Fang, Y. Regulating the Secondary Metabolism in Grape Berry Using Exogenous 24-Epibrassinolide for Enhanced Phenolics Content and Antioxidant Capacity. Food Chem. 2013, 141, 3056–3065. [Google Scholar] [CrossRef]
- Lacampagne, S.; Gagné, S.; Gény, L. Involvement of Abscisic Acid in Controlling the Proanthocyanidin Biosynthesis Pathway in Grape Skin: New Elements Regarding the Regulation of Tannin Composition and Leucoanthocyanidin Reductase (LAR) and Anthocyanidin Reductase (ANR) Activities and Expression. J. Plant Growth Regul. 2010, 29, 81–90. [Google Scholar] [CrossRef]
- Xi, Z.-M.; Meng, J.-F.; Huo, S.-S.; Luan, L.-Y.; Ma, L.-N.; Zhang, Z.-W. Exogenously Applied Abscisic Acid to Yan73 (V. vinifera) Grapes Enhances Phenolic Content and Antioxidant Capacity of Its Wine. Int. J. Food Sci. Nutr. 2013, 64, 444–451. [Google Scholar] [CrossRef]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Fernández-Fernández, J.I.; Bautista-Ortín, A.B.; Gil-Muñoz, R. Influence of Methyl Jasmonate and Benzothiadiazole on the Composition of Grape Skin Cell Walls and Wines. Food Chem. 2019, 277, 691–697. [Google Scholar] [CrossRef]
- Martins, V.; Unlubayir, M.; Teixeira, A.; Gerós, H.; Lanoue, A. Calcium and methyl jasmonate cross-talk in the secondary metabolism of grape cells. Plant Physiol. Biochem. 2021, 165, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Wang, X.; Xuan, X.; Sheng, Z.; Jia, H.; Emal, N.; Liu, Z.; Zheng, T.; Wang, C.; Fang, J. Characterization and Action Mechanism Analysis of VvmiR156b/c/d-VvSPL9 Module Responding to Multiple-Hormone Signals in the Modulation of Grape Berry Color Formation. Foods 2021, 10, 896. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, K.; Maoz, I.; Kochanek, B.; Sela, N.; Lerno, L.; Ebeler, S.E.; Lichter, A. Cytokinin but Not Gibberellin Application Had Major Impact on the Phenylpropanoid Pathway in Grape. Hortic. Res. 2021, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Boss, P.K.; Robinson, S.P. Treatment of Grape Berries, a Nonclimacteric Fruit with a Synthetic Auxin, Retards Ripening and Alters the Expression of Developmentally Regulated Genes. Plant Physiol. 1997, 115, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- Berli, F.J.; Bottini, R. UV-B and Abscisic Acid Effects on Grape Berry Maturation and Quality. J. Berry Res. 2013, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under Deficit Irrigation: Hints from Physiological and Molecular Data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.-H.; Yang, B.-H.; Wang, X.-W.; Li, J.-N.; Li, S.; Yang, X.; Ren, R.-H.; Fang, Y.-L.; Xu, T.-F.; Zhang, Z.-W.; et al. ABA signaling plays a key role in regulated deficit irrigation-driven anthocyanins accumulation in ‘Cabernet Sauvignon’ grape berries. Environ. Exp. Bot. 2021, 181, 104290. [Google Scholar] [CrossRef]
- Ryu, S.; Han, J.H.; Cho, J.G.; Jeong, J.H.; Lee, S.K.; Lee, H.J. High temperature at veraison inhibits anthocyanin biosynthesis in berry skins during ripening in ‘Kyoho’ grapevines. Plant Physiol. Biochem. 2020, 157, 219–228. [Google Scholar] [CrossRef]
- Alonso, R.; Berli, F.J.; Fontana, A.; Piccoli, P.; Bottini, R. Abscisic Acid’s Role in the Modulation of Compounds That Contribute to Wine Quality. Plants 2021, 10, 938. [Google Scholar] [CrossRef]
- Li, D.; Pang, Y.; Li, H.; Guo, D.; Wang, R.; Ma, C.; Xu, W.; Wang, L.; Wang, S. Comparative analysis of the gene expression profile under two cultivation methods reveals the critical role of ABA in grape quality promotion. Sci. Hortic. 2021, 281, 109924. [Google Scholar] [CrossRef]
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic Acid Is a Major Regulator of Grape Berry Ripening Onset: New Insights into ABA Signaling Network. Front. Plant Sci. 2017, 8, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F.; Ruan, G.; Liu, H. Analytical Methods for Tracing Plant Hormones. Anal. Bioanal. Chem. 2012, 403, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Granda, V.; Cuesta, C.; Álvarez, R.; Ordás, R.; Centeno, M.L.; Rodríguez, A.; Majada, J.P.; Fernández, B.; Feito, I. Rapid Responses of C14 Clone of Eucalyptus Globulus to Root Drought Stress: Time-Course of Hormonal and Physiological Signaling. J. Plant Physiol. 2011, 168, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.; Ji, L.; Xu, D. Dynamic Changes in Endogenous Hormones in Terminal Buds from Different Crown Positions in Sequoia Sempervirens (Lamb.) Endl. HortScience 2021, 56, 538–542. [Google Scholar] [CrossRef]
- Wu, L.; Lan, J.; Xiang, X.; Xiang, H.; Jin, Z.; Khan, S.; Liu, Y. Transcriptome sequencing and endogenous phytohormone analysis reveal new insights in CPPU controlling fruit development in kiwifruit (Actinidia chinensis). PLoS ONE 2020, 15, e0240355. [Google Scholar] [CrossRef]
- Yi, M.; Zhao, L.; Wu, K.; Liu, C.; Deng, D.; Zhao, K.; Li, J.; Deng, A. Simultaneous Detection of Plant Growth Regulators Jasmonic Acid and Methyl Jasmonate in Plant Samples by a Monoclonal Antibody-Based ELISA. Analyst 2020, 145, 4004–4011. [Google Scholar] [CrossRef]
- Leng, F.; Cao, J.; Wang, S.; Jiang, L.; Li, X.; Sun, C. Transcriptomic Analyses of Root Restriction Effects on Phytohormone Content and Signal Transduction during Grape Berry Development and Ripening. Int. J. Mol. Sci. 2018, 19, 2300. [Google Scholar] [CrossRef] [Green Version]
- Delatorre, C.; Rodríguez, A.; Rodríguez, L.; Majada, J.P.; Ordás, R.J.; Feito, I. Hormonal Profiling: Development of a Simple Method to Extract and Quantify Phytohormones in Complex Matrices by UHPLC–MS/MS. J. Chromatogr. B 2017, 1040, 239–249. [Google Scholar] [CrossRef]
- He, L.; Meng, N.; Castellarin, S.D.; Wang, Y.; Sun, Q.; Li, X.-Y.; Dong, Z.-G.; Tang, X.-P.; Duan, C.-Q.; Pan, Q.-H. Combined Metabolite and Transcriptome Profiling Reveals the Norisoprenoid Responses in Grape Berries to Abscisic Acid and Synthetic Auxin. Int. J. Mol. Sci. 2021, 22, 1420. [Google Scholar] [CrossRef]
- Rawlinson, C.; Kamphuis, L.G.; Gummer, J.P.A.; Singh, K.B.; Trengove, R.D. A Rapid Method for Profiling of Volatile and Semi-Volatile Phytohormones Using Methyl Chloroformate Derivatisation and GC–MS. Metabolomics 2015, 11, 1922–1933. [Google Scholar] [CrossRef] [Green Version]
- Engelberth, J.; Schmelz, E.A.; Alborn, H.T.; Cardoza, Y.J.; Huang, J.; Tumlinson, J.H. Simultaneous quantification of jasmonic acid and salicylic acid in plants by vapor-phase extraction and gas chromatography-chemical ionization-mass spectrometry. Anal. Biochem. 2003, 312, 242–250. [Google Scholar] [CrossRef]
- Giannarelli, S.; Muscatello, B.; Bogani, P.; Spiriti, M.M.; Buiatti, M.; Fuoco, R. Comparative determination of some phytohormones in wild-type and genetically modified plants by gas chromatography–mass spectrometry and high-performance liquid chromatography–tandem mass spectrometry. Anal. Biochem. 2010, 398, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Ge, L.; Lee, A.S.Y.; Yong, J.W.H.; Tan, S.N.; Ong, E.S. Simultaneous analysis of different classes of phytohormones in coconut (Cocos nucifera L.) water using high-performance liquid chromatography and liquid chromatography–tandem mass spectrometry after solid-phase extraction. Anal. Chim. Acta 2008, 610, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Dai, J.; Han, H.; Wang, C.; Zhu, L.; Lu, C.; Chen, H. Determination of Thirteen Acidic Phytohormones and Their Analogues in Tea (Camellia Sinensis) Leaves Using Ultra High Performance Liquid Chromatography Tandem Mass Spectrometry. J. Chromatogr. B 2020, 1149, 122144. [Google Scholar] [CrossRef] [PubMed]
- Oklestkova, J.; Tarkowská, D.; Eyer, L.; Elbert, T.; Marek, A.; Smržová, Z.; Novák, O.; Fránek, M.; Zhabinskii, V.N.; Strnad, M. Immunoaffinity chromatography combined with tandem mass spectrometry: A new tool for the selective capture and analysis of brassinosteroid plant hormones. Talanta 2017, 170, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Manzi, M.; Gómez-Cadenas, A.; Arbona, V. Rapid and reproducible determination of active gibberellins in citrus tissues by UPLC/ESI-MS/MS. Plant Physiol. Biochem. 2015, 94, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Barbier, F.; Yoneyama, K.; Beveridge, C.A. A Rapid Method for Quantifying RNA and Phytohormones From a Small Amount of Plant Tissue. Front. Plant Sci. 2020, 11, 605069. [Google Scholar] [CrossRef]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant Hormonomics: Multiple Phytohormone Profiling by Targeted Metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [Green Version]
- Floková, K.; Tarkowská, D.; Miersch, O.; Strnad, M.; Wasternack, C.; Novák, O. UHPLC–MS/MS Based Target Profiling of Stress-Induced Phytohormones. Phytochemistry 2014, 105, 147–157. [Google Scholar] [CrossRef]
- Xin, P.; Guo, Q.; Li, B.; Cheng, S.; Yan, J.; Chu, J. A Tailored High-Efficiency Sample Pretreatment Method for Simultaneous Quantification of 10 Classes of Known Endogenous Phytohormones. Plant Commun. 2020, 1, 100047. [Google Scholar] [CrossRef]
- Pan, X.; Welti, R.; Wang, X. Quantitative Analysis of Major Plant Hormones in Crude Plant Extracts by High-Performance Liquid Chromatography-Mass Spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhang, W.; Gao, J.; Lu, M.; Zhang, L.; Li, J. Simultaneous Determination of Plant Hormones in Peach Based on Dispersive Liquid–Liquid Microextraction Coupled with Liquid Chromatography–Ion Trap Mass Spectrometry. J. Chromatogr. B 2015, 992, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lu, S.; Wu, H.; Chen, G.; Liu, S.; Kong, X.; Kong, W.; You, J. Determination of Multiple Phytohormones in Fruits by High-Performance Liquid Chromatography with Fluorescence Detection Using Dispersive Liquid–Liquid Microextraction Followed by Precolumn Fluorescent Labeling. J. Sep. Sci. 2015, 38, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Campillo, N.; Viñas, P.; Férez-Melgarejo, G.; Hernández-Córdoba, M. Dispersive Liquid—Liquid Microextraction for the Determination of Three Cytokinin Compounds in Fruits and Vegetables by Liquid Chromatography with Time-of-Flight Mass Spectrometry. Talanta 2013, 116, 376–381. [Google Scholar] [CrossRef]
- Cai, W.-J.; Ye, T.-T.; Wang, Q.; Cai, B.-D.; Feng, Y.-Q. A Rapid Approach to Investigate Spatiotemporal Distribution of Phytohormones in Rice. Plant Methods 2016, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Flores, M.I.A.; Romero-González, R.; Frenich, A.G.; Vidal, J.L.M. QuEChERS-Based Extraction Procedure for Multifamily Analysis of Phytohormones in Vegetables by UHPLC-MS/MS. J. Sep. Sci. 2011, 34, 1517–1524. [Google Scholar] [CrossRef]
- Müller, M.; Munné-Bosch, S. Rapid and Sensitive Hormonal Profiling of Complex Plant Samples by Liquid Chromatography Coupled to Electrospray Ionization Tandem Mass Spectrometry. Plant Methods 2011, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Coombe, B.G. Growth Stages of the Grapevine: Adoption of a System for Identifying Grapevine Growth Stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Cho, S.-K.; Abd El-Aty, A.M.; Park, K.H.; Park, J.-H.; Assayed, M.E.; Jeong, Y.-M.; Park, Y.-S.; Shim, J.-H. Simple Multiresidue Extraction Method for the Determination of Fungicides and Plant Growth Regulator in Bean Sprouts Using Low Temperature Partitioning and Tandem Mass Spectrometry. Food Chem. 2013, 136, 1414–1420. [Google Scholar] [CrossRef]
- De Nicolò, A.; Cantù, M.; D’Avolio, A. Matrix Effect Management in Liquid Chromatography Mass Spectrometry: The Internal Standard Normalized Matrix Effect. Bioanalysis 2017, 9, 1093–1105. [Google Scholar] [CrossRef]
- Wright, M.J.; Wheller, R.; Wallace, G.; Green, R. Internal Standards in Regulated Bioanalysis: Putting in Place a Decision-Making Process during Method Development. Bioanalysis 2019, 11, 1701–1713. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zou, Y.; Kaw, H.Y.; Wang, G.; Sun, H.; Cai, L.; Li, C.; Meng, L.-Y.; Li, D. Recent Developments and Emerging Trends of Mass Spectrometric Methods in Plant Hormone Analysis: A Review. Plant Methods 2020, 16, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Liu, G.; Rao, Q.; Bai, B.; Zhao, Z.; Liu, H.; Wu, A. A Liquid Chromatography Tandem Mass Spectrometry Method for Simultaneous Determination of Acid/Alkaline Phytohormones in Grapes. J. Chromatogr. B 2012, 881–882, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; He, F.; Zhang, X.-K.; Shi, Y.; Duan, C.-Q. Impact of Three Phenolic Copigments on the Stability and Color Evolution of Five Basic Anthocyanins in Model Wine Systems. Food Chem. 2022, 375, 131670. [Google Scholar] [CrossRef] [PubMed]
- Crupi, P.; Coletta, A.; Anna Milella, R.; Perniola, R.; Gasparro, M.; Genghi, R.; Antonacci, D. HPLC-DAD-ESI-MS Analysis of Flavonoid Compounds in 5 Seedless Table Grapes Grown in Apulian Region. J. Food Sci. 2012, 77, C174–C181. [Google Scholar] [CrossRef]
- Yu, J.-N.; Meng, Q.-Y.; Liu, W.-J.; Lu, Y.-L.; Ren, X.-L. Analysis of Acidic Endogenous Phytohormones in Grapes by Using Online Solid-Phase Extraction Coupled with LC-MS/MS. J. Chromatogr. Sci. 2014, 52, 1145–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Ren, Z.-Y.; Wang, Y.; Fu, Y.-Q.; Li, Y.; Meng, N.; Pan, Q.-H. Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin Affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry. Plants 2020, 9, 630. [Google Scholar] [CrossRef]
- Davies, C.; Nicholson, E.L.; Böttcher, C.; Burbidge, C.A.; Bastian, S.E.P.; Harvey, K.E.; Huang, A.-C.; Taylor, D.K.; Boss, P.K. Shiraz Wines Made from Grape Berries (Vitis vinifera) Delayed in Ripening by Plant Growth Regulator Treatment Have Elevated Rotundone Concentrations and “Pepper” Flavor and Aroma. J. Agric. Food Chem. 2015, 63, 2137–2144. [Google Scholar] [CrossRef]
- Böttcher, C.; Johnson, T.E.; Burbidge, C.A.; Nicholson, E.L.; Boss, P.K.; Maffei, S.M.; Bastian, S.E.P.; Davies, C. Use of Auxin to Delay Ripening: Sensory and Biochemical Evaluation of Cabernet Sauvignon and Shiraz. Aust. J. Grape Wine Res. 2022, 28, 208–217. [Google Scholar] [CrossRef]
- Jia, H.; Xie, Z.; Wang, C.; Shangguan, L.; Qian, N.; Cui, M.; Liu, Z.; Zheng, T.; Wang, M.; Fang, J. Abscisic Acid, Sucrose, and Auxin Coordinately Regulate Berry Ripening Process of the Fujiminori Grape. Funct. Integr. Genom. 2017, 17, 441–457. [Google Scholar] [CrossRef]
- Suzuki, M.; Yamazaki, C.; Mitsui, M.; Kakei, Y.; Mitani, Y.; Nakamura, A.; Ishii, T.; Soeno, K.; Shimada, Y. Transcriptional Feedback Regulation of YUCCA Genes in Response to Auxin Levels in Arabidopsis. Plant Cell Rep. 2015, 34, 1343–1352. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytes | Transition | Fragmentor | College Energy | Retention Time (min) | Polarity |
|---|---|---|---|---|---|
| IP | 204.13 > 136 | 99 | 16 | 2.30 | Positive |
| GA3 | 345.13 > 239.1 | 162 | 12 | 3.34 | Negative |
| SA | 137 > 93 | 89 | 20 | 4.01 | Negative |
| IAA | 174.1 > 130 | 85 | 8 | 4.30 | Negative |
| ABA | 263.1 > 153 | 104 | 8 | 5.34 | Negative |
| JA | 209.12 > 59.1 | 118 | 12 | 6.47 | Negative |
| IBA | 202.1 > 158 | 128 | 12 | 6.57 | Negative |
| MeIAA | 190.1 > 130 | 84 | 24 | 6.91 | Positive |
| MeSA | 153.1 > 121 | 65 | 16 | 7.74 | Positive |
| MeJA | 225.15 > 151 | 95 | 12 | 9.45 | Positive |
| BL | 481.36 > 445.4 | 133 | 12 | 9.66 | Positive |
| TPP | 327 > 215 | 170 | 30 | 12.60 | Positive |
| Phytohormones | Green Pericarp | Red Pericarp | Green Seed | Brown Seed |
|---|---|---|---|---|
| IP | 80.5 ± 0.6 | 80.4 ± 7.9 | 79.5 ± 0.9 | 73.0 ± 3.0 |
| GA3 | 92.7 ± 2.5 | 95.8 ± 5.8 | 89.9 ± 2.5 | 91.0 ± 3.3 |
| SA | 105.8 ± 4.0 | 104.4 ± 8.2 | 104.9 ± 4.9 | 101.2 ± 5.0 |
| IAA | 88.3 ± 1.3 | 93.6 ± 5.0 | 87.2 ± 2.4 | 86.5 ± 2.9 |
| ABA | 90.6 ± 1.1 | 90.6 ± 0.5 | 82.6 ± 6.0 | 104.0 ± 6.6 |
| JA | 86.1 ± 3.4 | 84.0 ± 7.9 | 87.0 ± 6.5 | 85.9 ± 4.5 |
| IBA | 88.6 ± 2.7 | 85.4 ± 6.1 | 83.1 ± 2.5 | 87.5 ± 3.3 |
| MeIAA | 102.8 ± 13.6 | 119.1 ± 8.5 | 91.0 ± 9.5 | 90.0 ± 2.8 |
| MeSA | 91.1 ± 1.7 | 92.5 ± 7.4 | 76.1 ± 2.9 | 81.3 ± 5.2 |
| MeJA | 88.0 ± 3.6 | 90.8 ± 4.4 | 89.6 ± 1.6 | 93.0 ± 2.5 |
| BL | 92.0 ± 4.0 | 94.5 ± 5.5 | 92.4 ± 3.2 | 93.7 ± 2.8 |
| Phytohormones | Recovery | Coefficient of Variation | ||
|---|---|---|---|---|
| Low | Medium | High | ||
| IP | 75.1 ± 6.7 | 71.6 ± 3.2 | 67.2 ± 3.5 | 4.4 |
| GA3 | 91.4 ± 5.8 | 90.2 ± 1.9 | 85 ± 1.4 | 2.1 |
| SA | 110.1 ± 3.9 | 102.6 ± 2.3 | 111.4 ± 4 | 2.2 |
| IAA | 96.6 ± 1.7 | 99.1 ± 3 | 98.1 ± 1.6 | 3.0 |
| ABA | 77 ± 8.7 | 93.6 ± 10.6 | 106.1 ± 3.6 | 11.3 |
| JA | 94.4 ± 6.8 | 96.8 ± 4.2 | 118.2 ± 1.5 | 4.4 |
| IBA | 93 ± 1.5 | 97.5 ± 1.3 | 101.1 ± 1.5 | 1.3 |
| MeIAA | 84.2 ± 9.7 | 84.9 ± 3 | 110 ± 1.3 | 3.5 |
| MeSA | 79 ± 5.8 | 75.5 ± 2.3 | 63.6 ± 4.2 | 3.1 |
| MeJA | 102 ± 5.1 | 97 ± 4.6 | 91.9 ± 1.6 | 4.7 |
| BL | 101.1 ± 10.3 | 91 ± 2.7 | 88.9 ± 0.5 | 2.9 |
| Phytohormones | Equation of Linear Regression | R2 | Linear Range (ng/mL) | LOD (ng/mL) | LOQ (ng/mL) |
|---|---|---|---|---|---|
| IP | y = 0.6853x + 0.5383 | 0.9985 | 0.1–500 | 0.001 | 0.004 |
| GA3 | y = 240.3742x + 0.0169 | 0.9999 | 0.25–1000 | 0.12 | 0.39 |
| SA | y = 18.8627x + 0.2129 | 0.9997 | 0.25–500 | 0.30 | 1.00 |
| IAA | y = 1330.6574x + 1.1899 | 0.9999 | 5–1000 | 1.66 | 5.00 |
| ABA | y = 4.0284x − 0.6548 | 0.9980 | 0.1–250 | 0.002 | 0.007 |
| JA | y = 196.3287x + 0.1764 | 0.9994 | 0.25–500 | 0.10 | 0.33 |
| IBA | y = 7069.2793x − 2.4675 | 0.9987 | 5–250 | 1.60 | 5.00 |
| MeIAA | y = 6.0952x + 0.3178 | 0.9993 | 0.1–250 | 0.04 | 0.25 |
| MeSA | y = 1016.6302x + 1.0105 | 0.9982 | 2.5–500 | 0.75 | 2.50 |
| MeJA | y = 19.3897x + 0.1052 | 0.9995 | 0.25–250 | 0.10 | 0.25 |
| BL | y = 79.2625x + 0.9110 | 0.9999 | 0.1–1000 | 0.16 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, X.; Xia, N.; Meng, X.; Duan, C.; Pan, Q. A One-Step Polyphenol Removal Approach for Detection of Multiple Phytohormones from Grape Berry. Horticulturae 2022, 8, 548. https://doi.org/10.3390/horticulturae8060548
Yao X, Xia N, Meng X, Duan C, Pan Q. A One-Step Polyphenol Removal Approach for Detection of Multiple Phytohormones from Grape Berry. Horticulturae. 2022; 8(6):548. https://doi.org/10.3390/horticulturae8060548
Chicago/Turabian StyleYao, Xuechen, Nongyu Xia, Xiao Meng, Changqing Duan, and Qiuhong Pan. 2022. "A One-Step Polyphenol Removal Approach for Detection of Multiple Phytohormones from Grape Berry" Horticulturae 8, no. 6: 548. https://doi.org/10.3390/horticulturae8060548
APA StyleYao, X., Xia, N., Meng, X., Duan, C., & Pan, Q. (2022). A One-Step Polyphenol Removal Approach for Detection of Multiple Phytohormones from Grape Berry. Horticulturae, 8(6), 548. https://doi.org/10.3390/horticulturae8060548